苗木離體葉片水分狀況的葉綠素熒光參數定量診斷
2010-12-05 02:49:34段愛國楊文忠張建國張俊佩何彩云
浙江農林大學學報 2010年4期
段愛國,楊文忠,張建國,張俊佩,何彩云
(1.中國林業科學研究院 林業研究所 國家林業局林木培育重點實驗室,北京 100091;2.新疆林業科學研究院,新疆 烏魯木齊 830000)
葉綠素熒光分析技術能有效地了解逆境條件下葉片光系統對光能的吸收、傳遞、轉換、耗散、分配等內在性特點[1-2]。當植物離體或自然生長狀況下的葉片受到大氣或土壤水分脅迫時,植物葉片葉綠素熒光參數就會發生規律性變化,從而間接地指示不同干旱脅迫強度時植物光合生理的變化[3-9]。在定量分析方面,有研究指出,干旱脅迫條件下,葉綠素熒光參數的變化程度與抗旱性指標存在顯著相關性,可作為抗旱樹種篩選的評定指標[10-11]。總體而言,以往的研究較為側重于葉綠素熒光參數與水分脅迫程度間的定性分析,而相對缺乏脅迫過程中葉綠素熒光參數與水分脅迫程度的定量相關分析,不利于水分脅迫條件下熒光參數變化機制的系統揭示,進而一定程度上限制了熒光參數的說理性和指示性。基于此,本研究以云南省元謀縣金沙江干熱河谷區10種主要植被恢復樹種苗木離體葉片為試材,探討離體葉片室內自然失水過程中的葉綠素熒光反應機制,以期建立特定環境下苗木葉片水分動態與葉綠素熒光參數的內在相關關系,探索并初步實現植物葉片水分動態的熒光診斷。
1 材料與方法
1.1 植物材料
2006年4月初將采自云南省元謀縣干熱河谷區的10個主要植被恢復樹種種子播種在中國林業科學研究院溫室內,于2007年6月選取各樹種生長良好的單株盆栽苗木——印楝Azadirachta indica,山合歡Albizia kalkora,新銀合歡 Leucaena glauca,蘇門答臘金合歡 Acacia glauca,木豆 Cajanus cajan,赤桉Eucalyptus camaldolensia,檸檬桉Eucalyptus citriodora,大葉相思Acacia auriculiformis,小桐子Jatropha curcas和藍桉Eucalyptus globulue,連續澆水3 d,使苗木葉片充分吸水至飽和含水量,帶回實驗室。種子采集地的自然環境條件見參考文獻[15]。
1.2 試驗方法
剪取各苗木長30 cm帶6~12片葉的枝條,包扎切口處,置放于實驗平臺上,在室溫條件下自然失水 ,保持4盞日光燈的持續照射,葉面處光強約為320 μmol·m-2·s-1。分別在失水0,2,4,6,8,10,12,24,26,28,30,32,34,36,48,50,52,54,56,58,60 h 時,采用精確度達0.000 1 g的天平稱取各樹種枝條的質量,為適時鮮質量Mt(g),0 h的質量即為各個樹種枝條的飽和質量Ms(g),并在每一個樹種的離體枝條上固定3個葉片,采用美國產的OS1-FL便攜式調制葉綠素熒光儀(Fv/Fm模式和Yield模式)測定葉綠素熒光參數[16]。Fv/Fm模式是用強作用光激發產生固定熒光Fo,最大熒光Fm,可變熒光Fv及PSⅡ最大光化學效率Fv/Fm,并計算電子傳遞效率Fm/Fo和PSⅡ潛在活性Fv/F[2]。測定前對每一葉片均暗適應8 min左右;Yield模式是用飽和閃光脈沖激發最大穩態熒光Fms,穩態熒光Fs及PSⅡ有效光化學量子產量Fmv/Fms(Fmv=Fms-Fs),測定時不需暗適應。當各樹種枝條葉片完全枯萎或含水量基本保持不變時即停止測定。最后實驗結束時,在WMK-02型電熱恒溫培養箱中進行烘干,測定枝條干質量Md(g),并計算整個自然失水過程中枝葉的相對含水量CRW(%)及失水率 RWL(%)的變化,各指標計算公式如下:RWC=(Mt-Md)/(Ms-Md),RWL=(Ms-Mt)/(Ms-Md)。另取各樹種苗木一定量葉片,使其室溫條件下自然失水。在失水過程中,隔0.5 h采用精確度達0.000 1 g的天平稱取質量,同時采用WP4露點水勢儀測定各樹種葉水勢,至少記錄7個數據點,繪制各苗木葉片相對含水量與水勢的關系圖。
2 結果與討論
2.1 10種苗木葉片保水力比較
葉片保水力通常用來表示植物組織抗脫水的能力大小,與維持體內水分平衡的能力相關[17]。從表1可知,10種苗木離體葉片失水率隨失水時間的發展進程具有明顯差異。在失水2 h時,新銀合歡和木豆失水率已在20%以上,山合歡、大葉相思及檸檬桉失水率為15%~20%,大于其余5種苗木。這說明這幾個樹種早期失水控制能力較弱。在失水12 h時,10種苗木失水率大小排序為:小桐子<印楝<藍桉<大葉相思<赤桉<檸檬桉<木豆<蘇門答臘金合歡<新銀合歡<山合歡,其中,小桐子等前5種苗木的失水率控制在50%以下,后5種失水率均在50%以上,且差距較大。失水率越低,說明苗木保水能力越強。在失水24 h時,失水率的大小排序與失水12 h時基本一致。此后,小桐子與印楝的失水率一直低于大葉相思和3種桉樹。溫室播種苗葉片保水力分析結果表明,小桐子與印楝的葉片保水力高于另外8個樹種,大葉相思與3種桉樹保水力其次,3種合歡的葉片保水力最差,木豆保水力居于3種桉樹與3種合歡之間。
2.2 苗木葉片相對含水量與葉水勢的關系
葉片相對含水量與葉水勢為描述植物葉片水分狀況的2個特征參數,但由于葉片構造的不同,相同葉片含水量的不同樹種苗木的葉水勢往往具有很大差異。圖1描述了10種苗木葉片含水量與葉水勢的關系。從圖1可以看出,苗木葉水勢均隨葉片含水量的降低而下降,苗木葉水勢與含水量的關系可由2次拋物線予以準確擬合,相關系數相當高,均為極顯著相關(P<0.01),除藍桉為0.968 1外,其余9種苗木均達0.990 0以上。這表明通過相對簡單的葉片含水量的測定即可對苗木葉水勢作出有效的預測。從拋物線的形狀分析,赤桉、木豆、檸檬桉、大葉相思、小桐子、山合歡、新銀合歡和藍桉等8種苗木均表現為上 “凸”狀,這說明這幾種苗木葉水勢在葉片含水量下降初始階段的下降速度相對較慢,而在葉片含水量下降后期的下降趨勢更為顯著。蘇門答臘金合歡的葉片含水量與葉水勢關系曲線表現為下 “凸”狀,即其葉水勢隨葉片含水量的下降呈現先快后慢的下降趨勢。印楝葉水勢與葉片含水量的關系居于上述2種類型之間,其葉水勢隨葉含水量以0.668 9為斜率呈近乎直線狀下降。由于植物葉水勢測量往往是破壞性的,故以往探討兩者間連續相關關系的研究較少,但為便于不同研究間的比較分析,建立苗木葉片含水量(RWC)與葉水勢(ψwp)的關系深為必要。
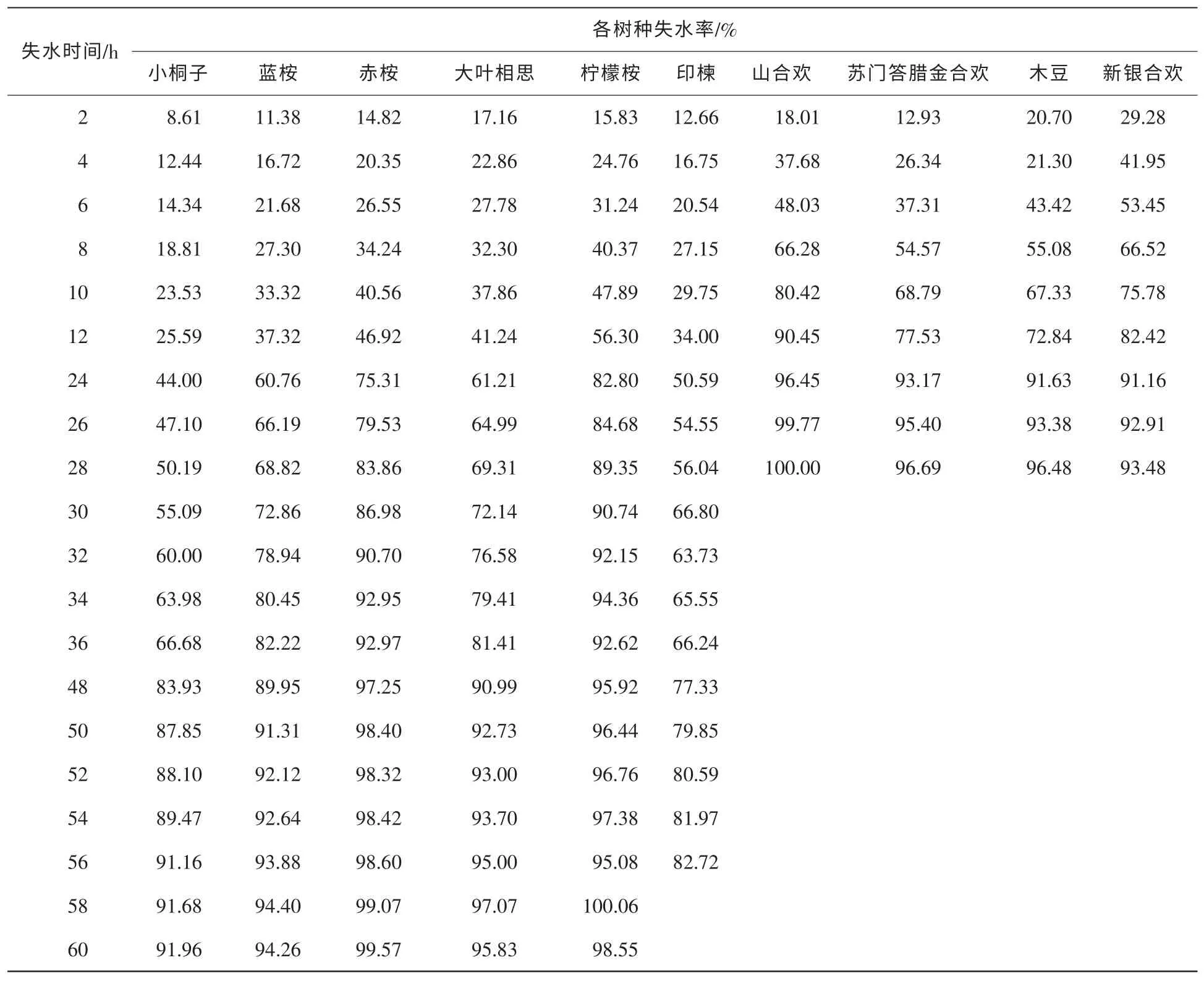
表1 10種苗木離體葉片失水率隨失水時間的變化Table 1 Change of water losing rate with water losing timeabout body-leaved leaves of ten species of seedlings

圖1 10種苗木葉片相對含水量與葉水勢的關系Figure 1 Relationship between relative water content and leaf potential of ten species of seedlings
2.3 持續失水條件下苗木葉片熒光參數的響應
2.3.1 Fo對水分脅迫的響應 固定熒光參數Fo的降低與上升可分別作為非光化學能量耗散和光合機構被破壞的重要指示性指標[15]。從圖2可以看到,Fo隨失水率(RWL)的變化規律可大致分為2種類型。第1種類型表現為Fo隨RWL的增大呈先下降后上升的變化趨勢,即在失水初期,處于安全含水量范圍的葉片以增加熱耗散的方式來保護光合機構免受傷害,而在失水一定程度后,葉片Fo上升,表明光合機構開始受到破壞。小桐子、印楝、大葉相思及赤桉屬于此一類型,藍桉與檸檬桉在失水2 h時Fo較為穩定,而在失水4 h時存在一個明顯下降點,此后Fo上升,總體上亦屬于第1種類型。第2種類型表現為Fo隨RWL的增大先維持不變而后上升的發展規律。屬于這一類型的樹種有新銀合歡、山合歡、蘇門答臘金合歡及木豆,它們在失水初期光合機構未啟動熱耗散機制亦未發生明顯可逆性失活現象,而是表現為一種忍耐干旱的適應特征。當失水至某一限度,光合機構發生失活現象。不同樹種光合機構失活的時刻不同,如新銀合歡出現在失水4 h,失水率達41.95%時刻,山合歡則出現在失水8 h,失水率達66.28%時刻。由于山合歡Fo在8 h時上升的速度非常迅速且失水率過大,不能肯定說其光合機構發生了可逆性失活,而很可能產生了不可逆性失活現象。第1種類型可謂主動防御型,而第2種類型則屬被動抵抗型。值得注意的是,第1種類型的樹種保水力較第2種類型大,這說明保水力越強的樹種,其啟動熱耗散機制保護光合機構免受干旱脅迫傷害的能力越強,且水分脅迫條件下發生可逆性失活的時刻越晚。可以很明顯地看到,在失水最后階段,各樹種熒光參數Fo均出現下降現象,此時,Fo的下降應已與熱耗散機制的啟動無關,因Fo反映了PSⅡ天線色素受激發后的電子密度,它與葉片葉綠素濃度有關[16],故而很可能是因為離體葉片過度失水后葉綠素含量降低的緣故。除定性方面的緊密關系外,熒光參數Fo與失水率間亦具有很強的數量關系,除藍桉外,其余9種苗木Fo隨失水率的變化曲線均可由二次拋物線予以恰當的描述,相關系數均達顯著或極顯著相關水平。
2.3.2 Fm,Fv及Fs對水分脅迫的響應 從圖3可以明顯看到,穩態熒光產量參數Fs隨失水率RWL的變化表現較為穩定,而熒光參數Fm和Fv隨失水率的增大總體上呈顯著下降趨勢,由于Fm和Fv分別指示最大熒光產量與PSⅡ原初電子受體QA的還原狀態,故這一結果表明PSⅡ反應中心處于完全關閉時的最大熒光產量與QA的可還原強度均隨葉片失水率的增大而下降。當Fm下降且逐漸接近Fo時,Fv逐漸下降至0。與Fo相同,Fm和Fv隨失水率的變化亦可分為2種類型。一種類型為Fm和Fv隨失水率的增大而一直呈明顯下降趨勢,另一類型為Fm和Fv隨失水率的增大呈先維持穩定而后下降的規律。2種類型所包含的樹種與Fo隨失水率的變動類型基本一致。這說明水分脅迫條件下,保水力強的樹種具有更靈敏的Fm和Fv響應機制,而保水力弱的樹種在脅迫初期具有更高的最大熒光產量和QA還原狀態的持續能力。
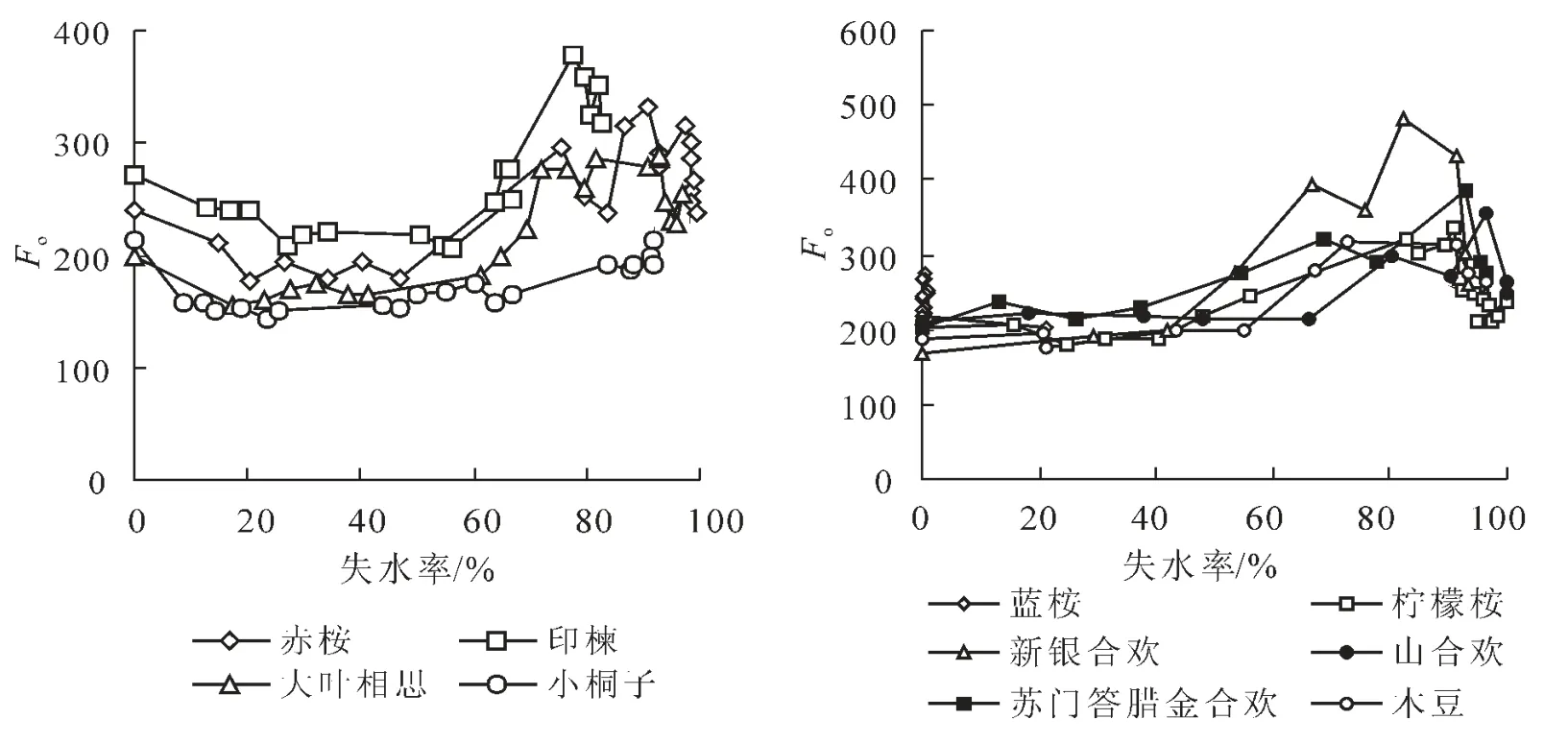
圖2 10種苗木離體葉片熒光參數Fo隨失水率的動態變化規律Figure 2 Dynamic change of fluorescence parameter Fowith water losing rate about body-leaved leaves of ten species of seedlings
2.3.3 Fv/Fm和Fmv/Fms與水分參數的定性與定量關系 從圖4可知,熒光參數Fv/Fm和Fmv/Fms隨失水率RWL的增大總體上呈逐漸下降的趨勢,但不同于Fm/Fo和Fv/Fo,其下降軌跡近似于一條水平拋物線,即在失水初期,Fv/Fm和Fmv/Fms變化較為平緩,而在失水后期,Fv/Fm和Fmv/Fms下降較為迅速。2個熒光參數在失水初期變化不明顯,表明一定范圍內的持續失水不能夠立即引起PSⅡ反應中心光化學效率的下降,這與輕度水分脅迫條件下植物葉片凈光合速率一般變化較小的現象相符[14]。Fv/Fm和Fmv/Fms的下降表明水分脅迫降低了PSⅡ反應中心的光化學效率和有效光化學量子產量[15]。當然,樹種不同,快速下降時刻與下降幅度亦不同。在整個失水過程中,熒光參數Fv/Fm一直略高于Fmv/Fms,生動地表明了PSⅡ有效光化學量子產量是PSⅡ最大光化學效率的主要組成部分。可以發現,在Fv/Fm和Fmv/Fms下降的過程中,各樹種均存在一個突降點,即熒光參數隨失水率的下降而快速下降的時刻,該時刻為PSⅡ反應中心失活過程中的關鍵點。不同樹種突降點出現的時刻不同,光化學效率Fv/Fm第1次降幅在0.10以上時,小桐子、大葉相思、印楝、新銀合歡、山合歡、赤桉、木豆、檸檬桉、蘇門答臘金合歡和藍桉等10樹種突降點對應的失水率依次降低,分別為83.93%,64.99%,63.73%,53.45%,48.03%,46.92%,43.42%,40.37%,37.31%和33.32%。且PSⅡ有效光化學量子產量參數Fmv/Fms的突降點出現的時刻較PSⅡ最大光化學效率參數Fv/Fm的突降點出現的時刻早,這意味著熒光參數Fmv/Fms比Fv/Fm能更靈敏地指示葉片光合機構遭受水分脅迫的重大傷害狀況,也就是說,極度的水分脅迫首先引起了PSⅡ有效光化學量子產量的嚴重下降。對于水分脅迫條件下Fv/Fm存在突降點的問題,以前的研究亦有發現[8],但由于熒光測定時間間隔過長,且不夠均勻,致使未能得到可靠結論,而本研究無疑再一次證明了該現象的存在。而突降點出現時刻所對應的失水率越大是否意味著樹種葉片光合機構忍耐干旱的能力愈強,則需要得到進一步的深入探討。表2定量地描述了離體葉片熒光參數Fv/Fm和Fmv/Fms隨失水率的數量變化關系。從表2及圖4可知,熒光參數Fv/Fm和Fmv/Fms與失水率存在極顯著的相關關系(P<0.01),二次拋物線方程的相關指數均在0.85以上,絕大部分超過了0.95。這表明熒光參數Fv/Fm和Fmv/Fms與葉片含水量具有很強的數量相關關系,且由于葉片含水量與葉水勢相關關系緊密,故Fv/Fm和Fmv/Fms與葉水勢亦應具有很強的相關關系。很明顯,熒光參數Fv/Fm和Fmv/Fms具有量化診斷葉片水分狀況的潛在功能,因此,除遙感方法外[17],葉綠素熒光參數亦可望成為植物葉片失水率無損傷測定的指示性指標。根據圖1中葉水勢與相對含水量的關系以及表2可建立熒光參數Fv/Fm和Fmv/Fms與葉水勢的關系式。當葉水勢為-1.5 MPa時,10種苗木離體葉片PSⅡ有效光化學量子產量Fmv/Fms由大到小依次為:赤桉(0.79)>小桐子(0.77)=大葉相思(0.77)>新銀合歡(0.68)>印楝(0.67)和山合歡(0.63)>木豆(0.62)>蘇門答臘金合歡(0.61)>檸檬桉(0.58)>藍桉(0.53)。Fmv/Fms越大,表明水分脅迫條件下苗木葉片光合機構實際光能捕獲效率越高,苗木潛在耐旱生產能力越強。
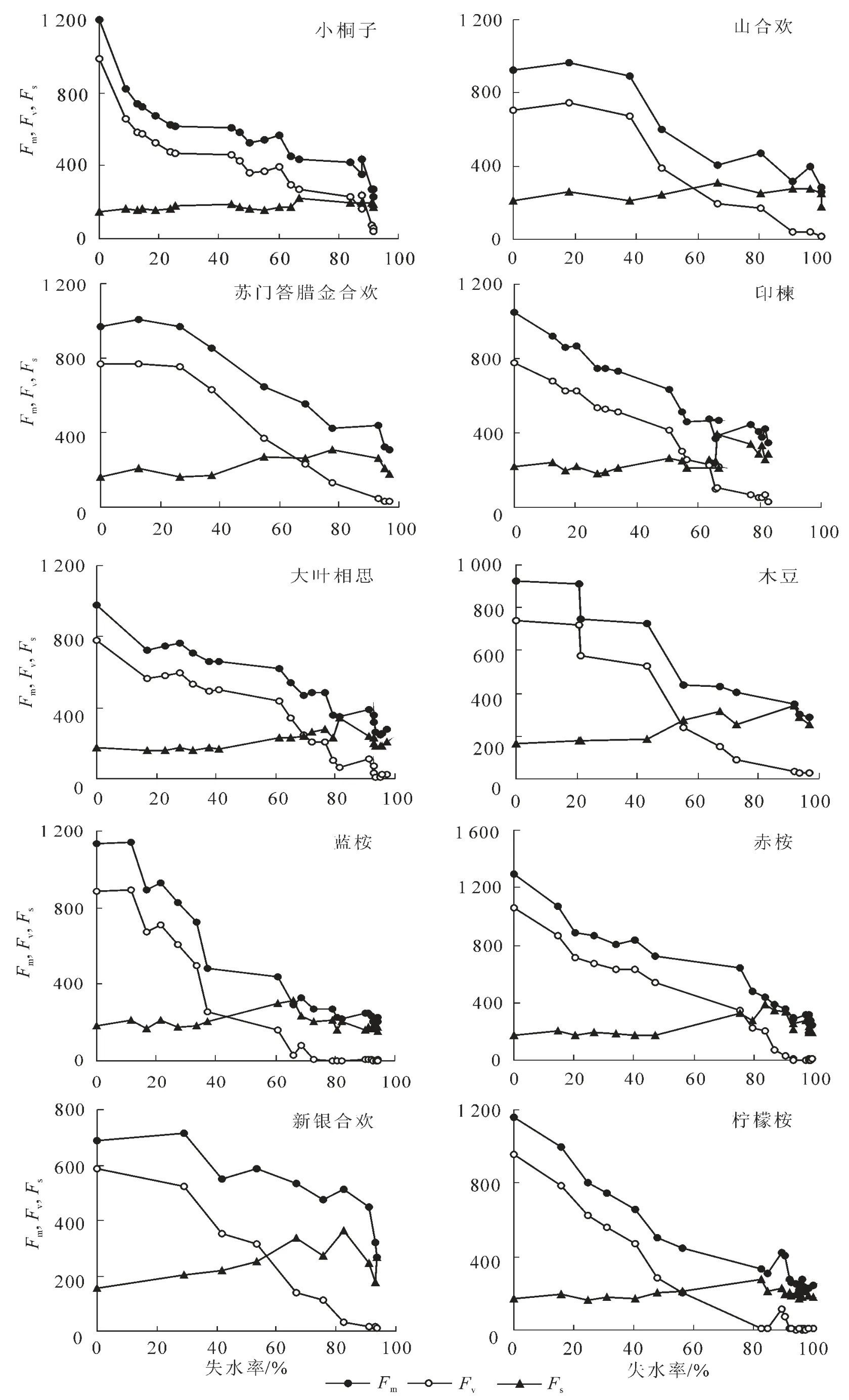
圖3 10種苗木離體葉片熒光參數Fm,Fv,Fs隨失水率的動態變化規律Figure 3 Dynamic change of fluorescence parameter Fm,Fv,Fswith water losing rate about body-leaved leaves of ten species of seedlings
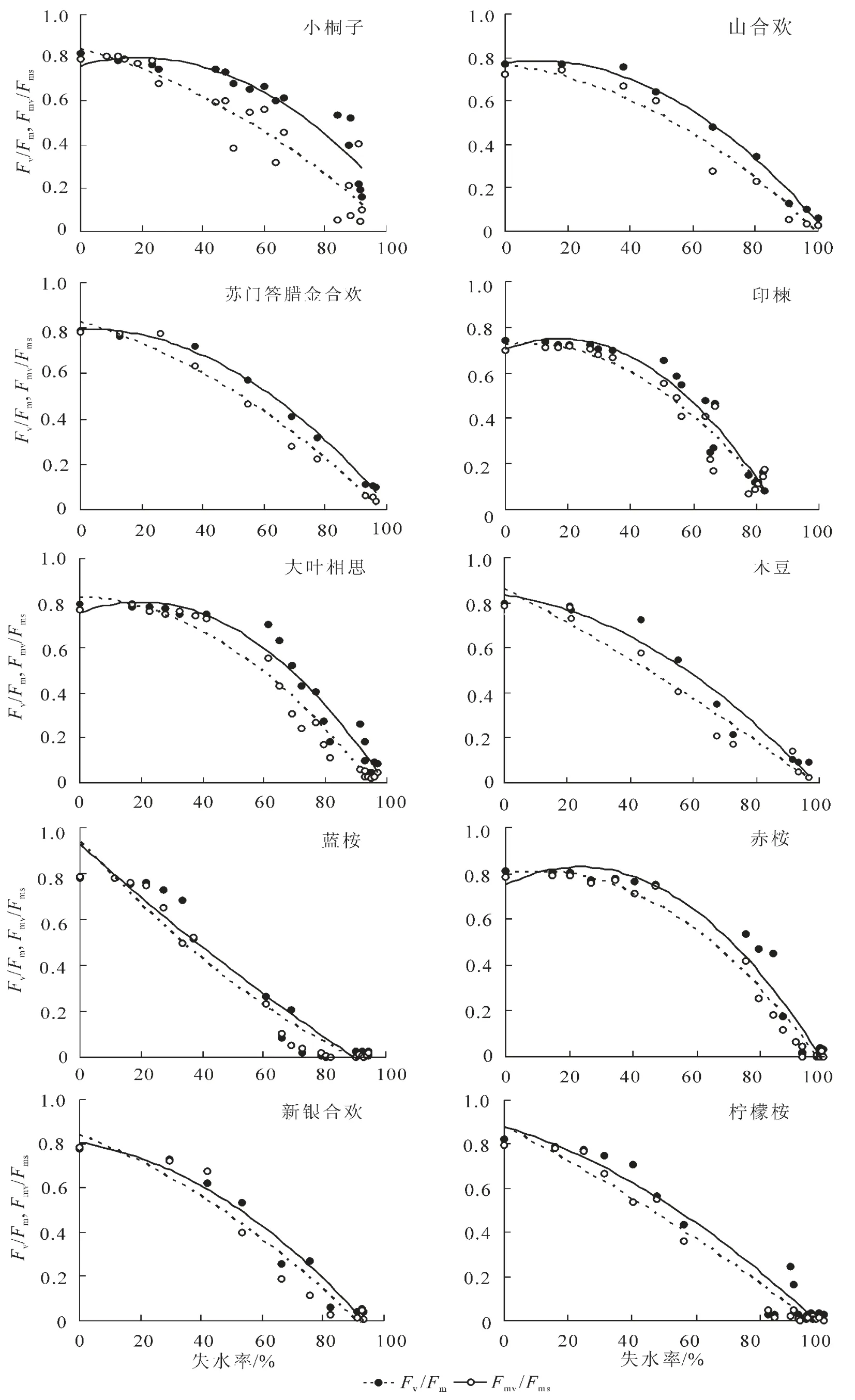
圖4 10種苗木離體葉片熒光參數Fv/Fm和Fmv/Fms隨失水率的動態變化規律Figure 4 Dynamic change of fluorescence parameter Fv/Fm,Fmv/Fmswith water losing rate about body-leaved leaves of ten species of seedlings
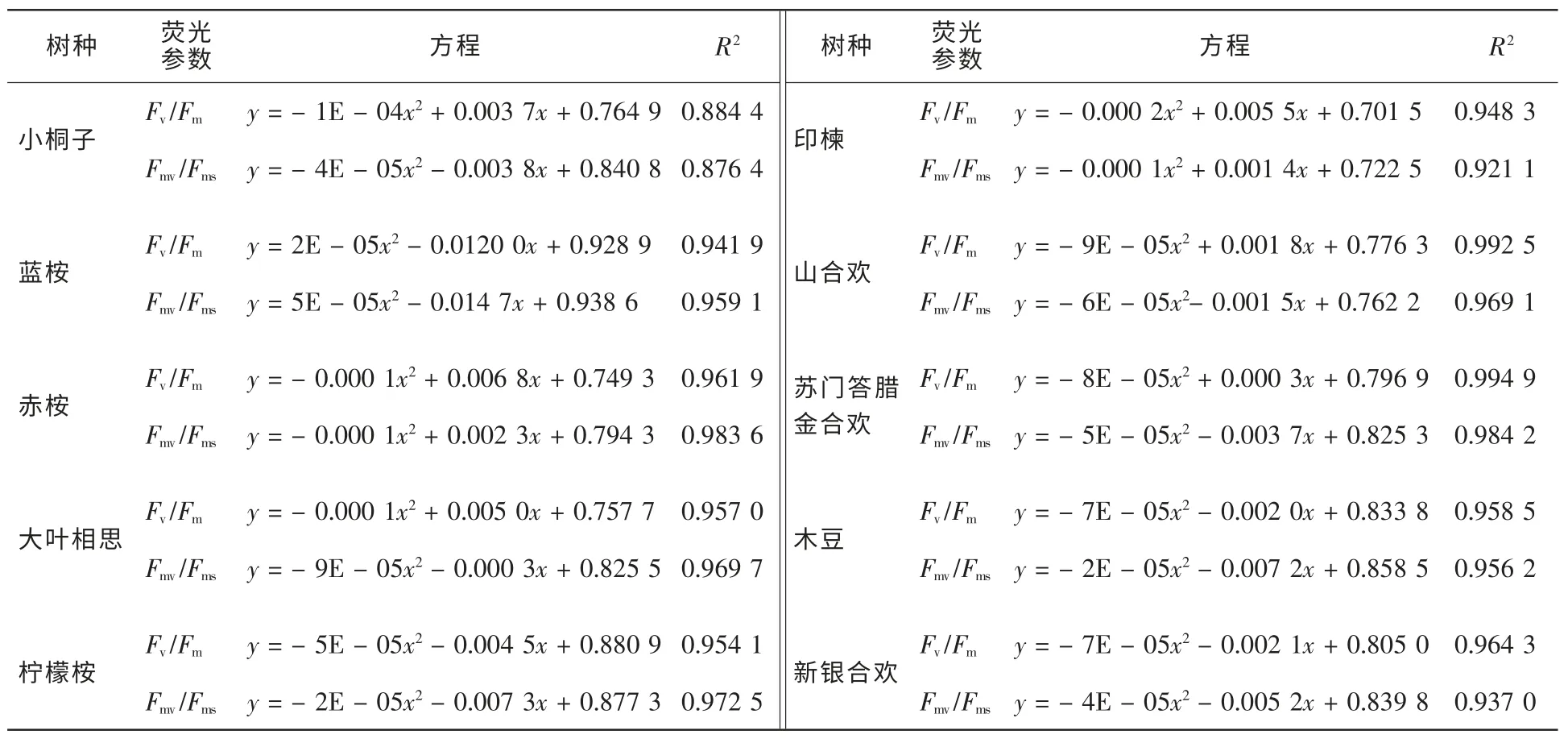
表2 10種苗木離體葉片熒光參數Fv/Fm和Fmv/Fms與失水率的數量關系式Table 2 Mathematical model of fluorescence parameters Fv/Fm,Fmv/Fmsand water losing rate about body-leaved leaves of ten species of seedlings
3 結論
離體葉片水分狀況與葉綠素熒光參數的數量關系研究對于揭示供試樹種干旱逆境條件下的內在光合作用機制和活體診斷葉片水分狀況具有重要參考價值。筆者以干熱河谷區10種主要植被恢復樹種為材料,探討了苗木離體葉片室內自然失水條件下葉綠素熒光參數的響應特征,主要結論有:①小桐子與印楝的葉片保水力最高,大葉相思與3種桉樹保水力其次,3種合歡的葉片保水力最差,木豆保水力居于3種桉樹與3種合歡之間。②不同樹種苗木葉水勢均隨葉片含水量的降低而下降,兩者的關系可由二次拋物線予以準確擬合,相關指數均在0.95以上,可通過相對簡單的葉片含水量的測定對苗木葉水勢作出有效的預測。③熒光參數Fo隨失水率的變化規律大致可分為主動防御型和被動抵抗型等2種類型,主動防御型表現為Fo隨失水率的增大呈先下降后上升的變化趨勢,被動抵抗型表現為Fo隨失水率的增大先維持基本不變而后上升的發展規律,主動防御型樹種的葉片保水力較被動抵抗型大。這說明保水力越強的樹種,其啟動熱耗散機制保護光合機構免受干旱脅迫傷害的能力越強,且水分脅迫條件下發生可逆性失活的時刻越晚。④熒光參數Fm和Fv隨失水率的增大總體上呈顯著下降趨勢,Fv/Fm和Fmv/Fms隨失水率的增大總體上呈逐漸下降的趨勢,下降軌跡近似于一條水平拋物線,在Fv/Fm和Fmv/Fms下降的過程中,各樹種均存在一個突降點,即熒光參數隨失水率的下降而快速下降的時刻,該時刻為PSⅡ反應中心失活過程中的關鍵點。⑤熒光參數Fmv/Fms比Fv/Fm能更靈敏地指示葉片光合機構遭受水分脅迫的重大傷害狀況,熒光參數Fv/Fm和Fmv/Fms與葉片水分參數呈現很強的數量相關關系,具有作為植物葉片水分狀況診斷指標的廣闊前景。
[1]HORTON P,RUBSN A V,WALTERS R G.Regulation of light harvesting in green plants:indication by nonphotochemical quenching of chlorophyll fluorescence[J].Plant Physiol,1994,106:415-420.
[2]許大全,張玉全.植物光合作用的光抑制[J].植物生理學通訊,1992,28(4):237-243.XU Daquan,ZHANG Yuquan.Photoinhibition of photosynthesis in plants[J].Plant Phys Commun,1992,28(4):237-243.
[3]GOVINDJEE D,FORK D C,ARMOND P A.Chlorophyll afluorescence transient as an indicator of water stress in maize plants[J].Plant Sci Lett,1981,20:191-194.
[4]譚新星,許大全.葉綠素缺乏的大麥突變體的光合作用和葉綠素熒光[J].植物生理學報,1996,22(1):51-57.TAN Xinxing,XU Daquan.Leaf photosynthesis and chlorophyll fluorescence in a chlorophyll deficient mutant of barley[J].Acta Phytophys Sin,1996,22(1):51-57.
[5]王可玢,趙福洪,王孝宣,等.用體內葉綠素a熒光誘導動力學鑒定番茄的抗冷性[J].植物學通報,1996,13(2):29-33.WANG Kefen,ZHAO Fuhong,WANG Xiaoxuan,et al.In vivo chlorophyll a fluorescence induction kinetics as a tool for chilling-tolerance detect in tomato[J].Chin Bull Bot,1996,13(2):29-33.
[6]趙會杰,鄒琦,于振文.葉綠素熒光分析技術及其在植物光合機理研究中的應用[J].河南農業大學學報,2000,34(3):248-251.ZHAO Huijie,ZOU Qi,YU Zhenwen.Chlorophyll fluoresence analysis technique and its application to photosynthesis of plant[J].J Henan Agric Univ,2000,34(3):248-251.
[7]李樹華,許興,鄭國琦.牛心樸子葉綠素熒光參數日變化及其與氣象因子的關系[J].干旱地區農業研究,2003,21(2):91-94.LI Shuhua,XU Xin,ZHENG Guoqi.The daily change in chla fluorescence parameters and the relationship between it and weather factors[J].Agric Res Arid Areas,2003,21(2):91-94.
[8]段愛國,保爾江,張建國.水分脅迫下華北地區主要造林樹種離體枝條葉片的葉綠素熒光參數[J].林業科學研究,2005,18(5):578-584.DUAN Aiguo,BAO Erjiang,ZHANG Jianguo.Response of fluorescence parameters in detached leaves of several tree species in Huabei district to drought stress[J].For Res,2005,18(5):578-584.
[9]王利英,樓爐煥,王超,等.3種冬青屬植物氣體交換參數及葉綠素熒光特性[J].浙江林學院學報,2009,26(1):27-31.WANG Liying,LOU Luhuan,WANG Chao,et al.Leaf gas exchange and chlorophyll fluorescence parameters in three species of Ilex[J].J Zhejiang For Coll,2009,26(1):27-31.
[10]林世青,許春輝,張其德.葉綠素熒光動力學在植物抗性生理學、生態學和農業現代化中的應用[J].植物學通報,1992,9(1):1-16.LIN Shiqing,XU Chunhui,ZHANG Qide.Some application of chlorophyll fluorescence kinetics to plant stress physiology phytoecology and agricultural modernization[J].Chin bull Bot,1992,9(1):1-16.
[11]蒲光蘭,周蘭英,胡學華,等.干旱脅迫對金太陽杏葉綠素熒光動力學參數的影響[J].干旱地區農業研究,2005,23(3):44-48.PU Guanglan,ZHOU Lanying,HU Xuehua,et al.Effect of soil drought stress on characteristics of chlorophyll fluorescence in Jintaiyang apricot variety[J].Agric Res Arid Areas,2005,23(3):44-48.
[12]段愛國,張建國,張俊佩,等.金沙江干熱河谷主要植被恢復樹種葉水勢的時空變化規律[J].林業科學研究,2007,20(2):151-159.DUAN Aiguo,ZHANG Jianguo,ZHANG Junpei,et al.Spatial and temporal variations of water potential of the main tree species for vegetation restoration in the dry-hot valleys of the Jinsha River[J].For Res,2007,20(2):151-159.
[13]HAVAUX M,LANNOYE R.Chlorophyll afluorescence induction:a sensitive indicator of water stress in maize plants[J].Irrig Sci,1983,4:147-151.
[14]張建國,李吉躍,沈國舫.樹木耐旱特性及其機理研究[M].北京:中國林業出版社,2000:31-54.
[15]張守仁.葉綠素熒光動力學參數的意義及討論[J].植物學通報,1999,16(4):444-448.ZHANG Shouren.A discussion oil chlorophyll fluorescence kinetics parameters and their significance[J].Chin Bull Bot,1999,16(4):444-448.
[16]KRAUSE G H,WEIS E.Chlorophyll fluorescence as a tool in plant physiology(Ⅱ)interpretation of fluorescence signals[J].Photosynth Res,1984,5:139-157.
[17]王紀華,趙春江,郭曉維,等.利用遙感方法診斷小麥葉片含水量的研究[J].華北農學報,2000,15(4):68-72.WANG Jihua,ZHAO Chunjiang,GUO Xiaowei,et al.Study on the water content of wheat leaves by the remote sensing[J].Acta Agric Bor Sin,2000,15(4):68-72.
