南方紅豆杉離體胚培養誘導不定芽研究
2010-12-05 02:50:08曾余力林新春桂仁意張翠萍黃麗春
浙江農林大學學報 2010年4期
曾余力,林新春,桂仁意,張翠萍,黃麗春
(浙江農林大學 浙江省現代森林培育重點實驗室,浙江 臨安 311300)
南方紅豆杉Taxus chinensis var.mairei屬國家一級瀕危保護植物之一,集材用、藥用、觀賞于一體,經濟價值極高。由于紅豆杉屬Taxus植物本身特殊的生物學特性,種子在天然條件下,需要兩冬一夏才能萌發并且植株生長緩慢[1],常規育種難以保證市場的需要,故通過離體胚培養技術克服紅豆杉種子難以萌發的缺點,以期加快植株生長。近年來,國內外學者對短葉紅豆杉Taxus brevifolia,歐洲紅豆杉Taxus baccata,雜種紅豆杉Taxus×media var.hicksii,東北紅豆杉Taxus cuspidata等進行了離體胚培養研究[2-3],從不同角度證實離體胚的萌發、成苗與培養基、種子來源、培養條件等有密切關系,但對南方紅豆杉離體胚培養的研究較少,僅見臧新等[4-5]對南方紅豆杉離體胚萌發影響因素的研究。本研究在前人的研究基礎上以南方紅豆杉的成熟胚為外植體,篩選外植體滅菌途徑,基本培養基、植物生長調節物質等因子對離體胚生長和分化不定芽的影響。
1 材料與方法
1.1 試驗材料
試驗材料購自浙江省嵊州市常青花木場,于4℃冰箱中冷藏備用。
1.2 試驗方法
1.2.1 外植體滅菌 選取均勻飽滿的種子,剝去外種殼,流水沖洗12 h,置于超凈工作臺內,分別置于 5.0,10.0,25.0 g·L-1的次氯酸鈉(NaClO)溶液中真空滅菌 5,10,15 min,無菌水分別沖洗 5遍。最后在解剖顯微鏡下切除胚乳挑取種胚,接入MS(Murashige and Skoog)培養基中,1個種胚·管-1,各接種100管。接種后先暗培養2 d,然后再光培養。
1.2.2 不同基本培養基與植物生長調節物質對離體胚培養的影響 試驗一,挑取種胚分別接種于MS,1/2MS,1/3MS,1/4MS,WPM(woody plant medium)和 B5(Gamborg)等 6種基本培養基中,測試基本培養基的影響;試驗二,以WPM為基本培養基,測試不同種類和質量濃度的植物生長調節物質的影響:噻苯隆(TDZ)為 0,0.000 1,0.001 0,0.010 0,0.100 0 mg·L-1,細胞激動素(KT),6-芐氨基腺嘌呤(6-BA),異戊烯基腺嘌呤(2iP)和玉米素(ZT)為 0,0.030 0,0.300 0,1.000 0,3.000 0,10.000 0 mg·L-1。
1.2.3 種胚萌動對離體胚培養的影響 根據種胚大小、顏色分別選取萌動(長度為0.7~0.8 cm,顏色轉微黃色)和未萌動(長度為0.4~0.5 cm,顏色白色)的種胚,測試不同發育時期對胚培養的影響。以WPM為基本培養基,附加 0,0.010 0,0.100 0,1.000 0 mg·L-1的 6-BA。以上試驗培養條件:溫度為(25±2)℃,光照度為2 500~2 800 lx,光周期(晝/夜)16/8 h。對照組(ck)為WPM基本培養基。
1.2.4 馴化移栽 將帶根的試管苗置于馴化室中,在強光(2萬 lx)下練苗4周,然后取出試管苗,沖凈根部培養基,將植株移至泥炭 ∶珍珠巖 ∶蛭石為1∶1∶1的人工基質中。
1.3 統計分析
以上各試驗設計3個重復,各試驗的結果分析均采用生物統計法將平均值及其標準誤差[6]和95%的可信度進行差異顯著性分析[7]。
2 結果與分析
2.1 外植體滅菌
培養28 d后調查,結果見表1。最優滅菌處理為25.0 g·L-1次氯酸鈉溶液真空滅菌10 min,其污染率僅6%,出苗率達86%。試驗結果表明,隨次氯酸鈉質量濃度升高,污染率顯著降低,而滅菌時間加長也能有效降低污染。由于南方紅豆杉胚乳較厚,高質量濃度次氯酸鈉浸泡不會損傷到離體胚的萌發生長,各處理的萌發率為93%~98%。
2.2 不同基本培養基與植物生長調節劑對離體胚培養的影響
2.2.1 不同基本培養基對離體胚培養的影響 將南方紅豆杉離體種胚接于不同基本培養基中,培養2個月后,MS培養基中的胚,葉色淡綠,莖較長較粗,但根較短無側根,根長約2.0 cm。在不同MS質量濃度(1/3MS,1/2MS,MS)培養基的胚,隨無機鹽質量濃度升高,芽長增長;在1/3MS中根長最長,根數最多。可能是南方紅豆杉莖伸長生長需要高質量濃度的無機鹽離子,根生長則以低質量濃度無機鹽離子為宜,而莖葉和根的平衡生長則需要大量元素的協調配比。對應MS培養基,WPM培養基中離體胚莖段細長,葉色深綠,并有許多側根生成,根系好,主根長度達3.8 cm,平均根數達4.3條,是南方紅豆杉胚培養的最佳基本培養基;B5培養基由于莖細弱,葉色發黃,不適于南方紅豆杉的胚培養(圖1~3)。
2.2.2 不同植物生長調節物質對離體胚培養的影響 將離體種胚接種于含有不同質量濃度的6-BA的培養基中,50 d調查結果如表2。與對照組相比,6-BA質量濃度遞增時,離體胚抽出莖葉的長度遞降,胚根長度變短甚至不萌發,胚軸上有突起發生。在6-BA 0.03 mg·L-1時,胚軸有突起形成,少數有不定芽分化;質量濃度增加至0.10 mg·L-1,不定芽數量增加;但6-BA質量濃度增高到1.00 mg·L-1時,突起變少,且突起不能分化為不定芽,并隨之褐化。將離體種胚接種于含有不同質量濃度的KT
的培養基中,50 d調查結果如表3。由表3可知,0.10 mg·L-1KT的培養基中離體胚無莖葉抽出和胚根伸長,但可見叢生不定芽的分化,誘導率達30%。與6-BA相比,KT更優于不定芽的分化,但對莖葉和胚根生長的抑制較明顯;當KT質量濃度升高至0.30 mg·L-1時不定芽的誘導率降低,而且隨KT質量濃度的升高,對胚根發育的抑制越強;當質量濃度提高至10.00 mg·L-1時,離體胚無生長、褐化。試驗發現,接種于含6-BA的培養基中的離體胚僅在胚軸上有突起產生,分化少量不定芽,而接種于含KT培養基中的離體胚在胚軸、子葉和生長點均有不定芽的分化。將離體種胚接種于含不同濃度ZT的培養基中,50 d調查結果如表4。由表4可知,ZT也能有效誘導不定芽的分化。當ZT的質量濃度在0.10~1.00 mg·L-1時,胚軸、生長點和子葉均有不定芽分化,且多為叢生芽,當ZT質量濃度為1.00 mg·L-1時誘導率高達50%,與KT不同的是,ZT對莖葉抽出抑制不明顯,KT質量濃度為0.10 mg·L-1時無莖葉抽出,而ZT的質量濃度為1.00 mg·L-1時仍有莖葉抽出,但節間較短,并且有輕微水化現象;當ZT升至3.00 mg·L-1時,水化更為明顯。在含不同質量濃度2ip的培養基中,2iP對不定芽的誘導較6-BA強(表5),對南方紅豆杉離體胚不定芽的誘導率最高,質量濃度為0.30 mg·L-1時,誘導率高達70%,但僅有1~2個單芽·胚-1,而KT和ZT的不定芽多為叢芽。對莖葉抽出抑制效果低于6-BA,當質量濃度為3.00 mg·L-1莖葉仍有抽出,葉色較綠。添加TDZ的試驗處理,都無不定芽分化。低質量濃度TDZ中試管苗生長與對照組相似,質量濃度高于0.100 0 mg·L-1,胚軸膨脹,試管苗有水化現象;當質量濃度增加到1.000 0 mg·L-1,種胚呈黃褐色愈合組織(表6)。
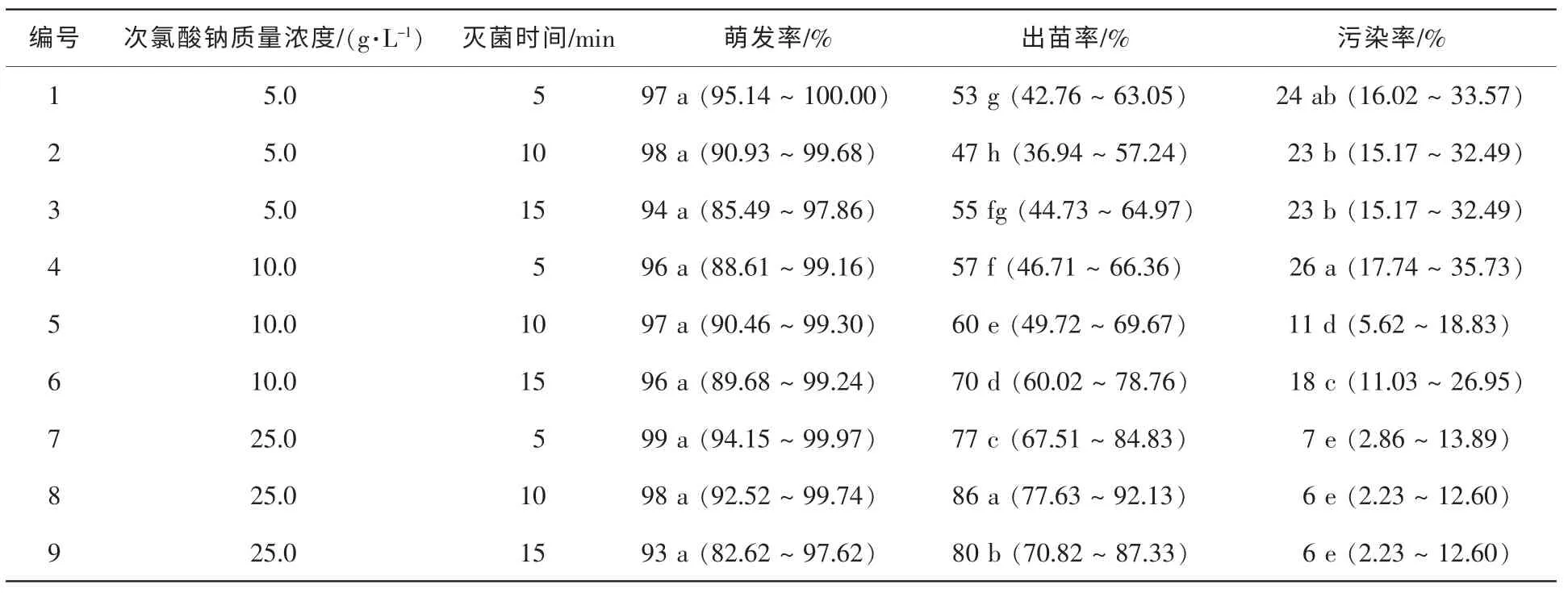
表1 種子表面滅菌對離體種胚培養的影響Table 1 Effect of factorial tests among various concentrations of NaClO vs.its durations on germination of excised embryos
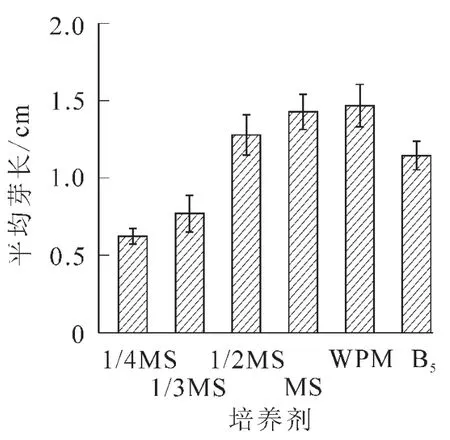
圖1 不同基本培養基對芽體生長的影響Figure 1 Effects of basal media on shoot growth of Taxus chinensis var.mairei
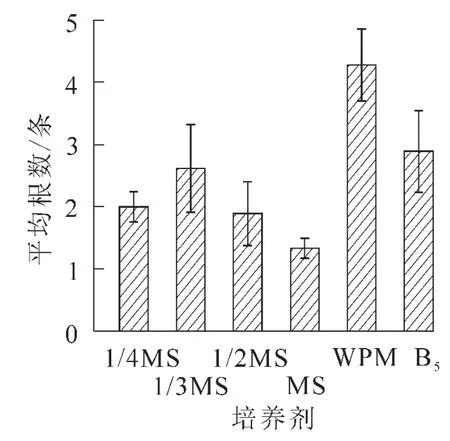
圖2 不同基本培養基對根數的影響Figure 2 Effects of basal media on root number of Taxus chinensis var.mairei
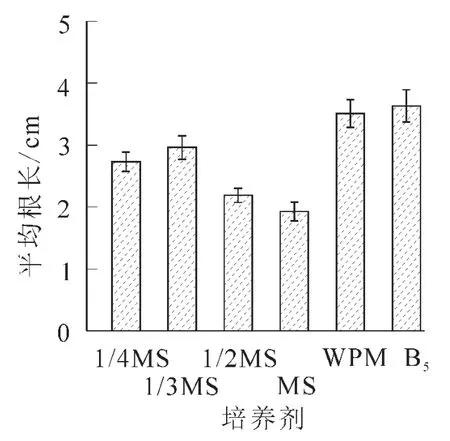
圖3 不同基本培養基對根長生長的影響Figure 3 Effect of basal media on root growth of Taxus chinensis var.mairei
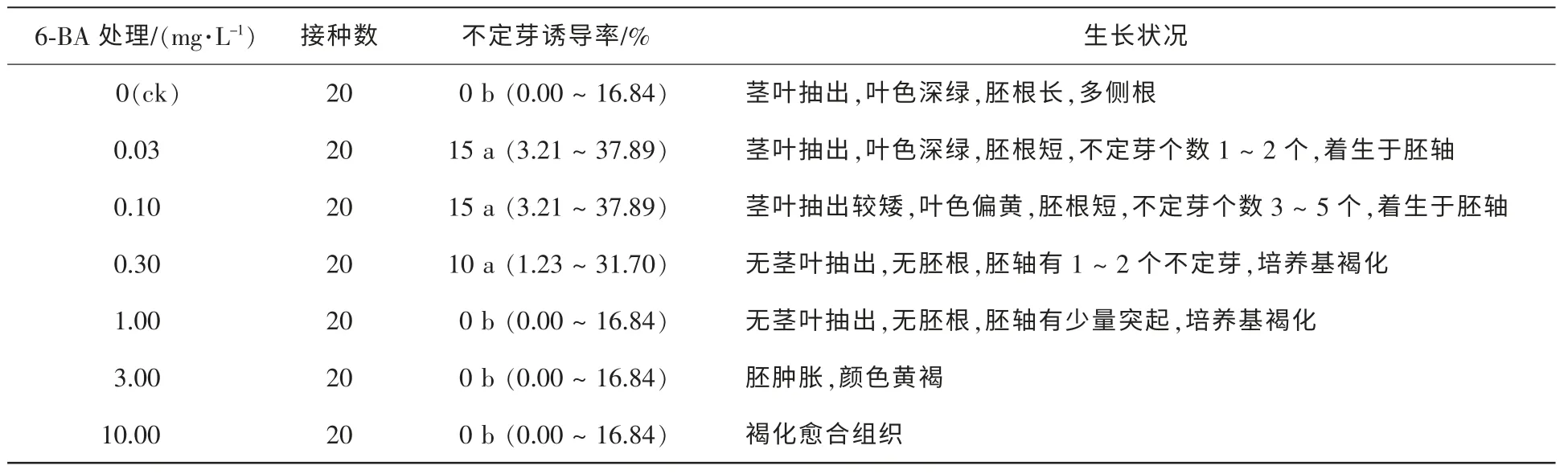
表2 6-BA對種胚培養誘導不定芽分化的影響Table 2 Effect of 6-BA on induction of adventitious buds by embryo culture
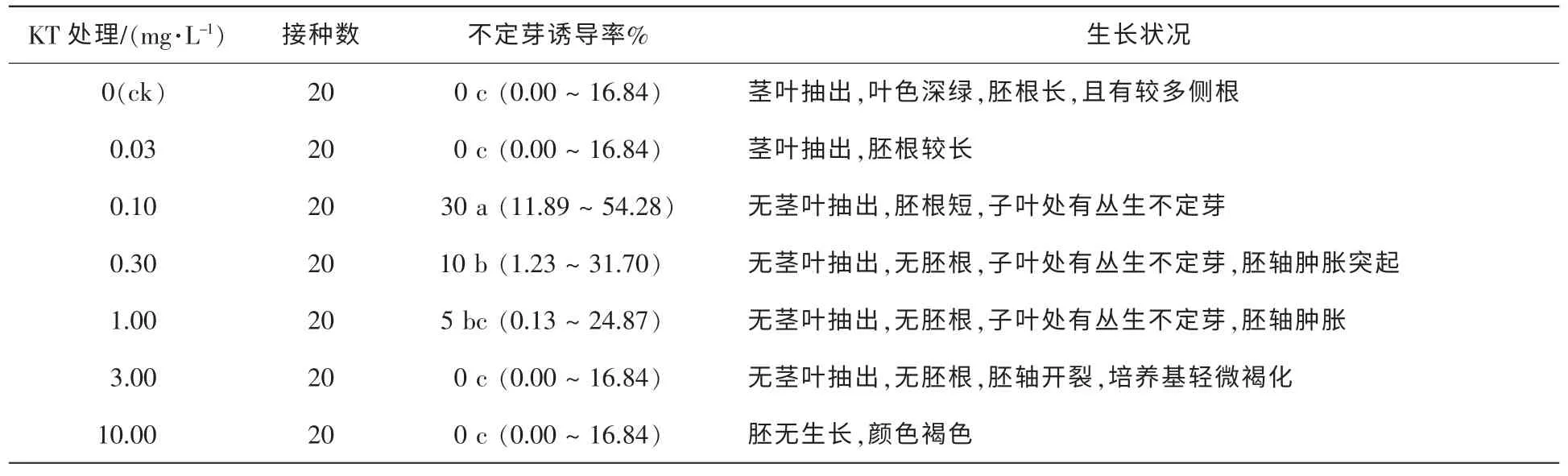
表3 KT對種胚培養誘導不定芽分化的影響Table 3 Effect of KT on induction of adventitious buds by embryo culture
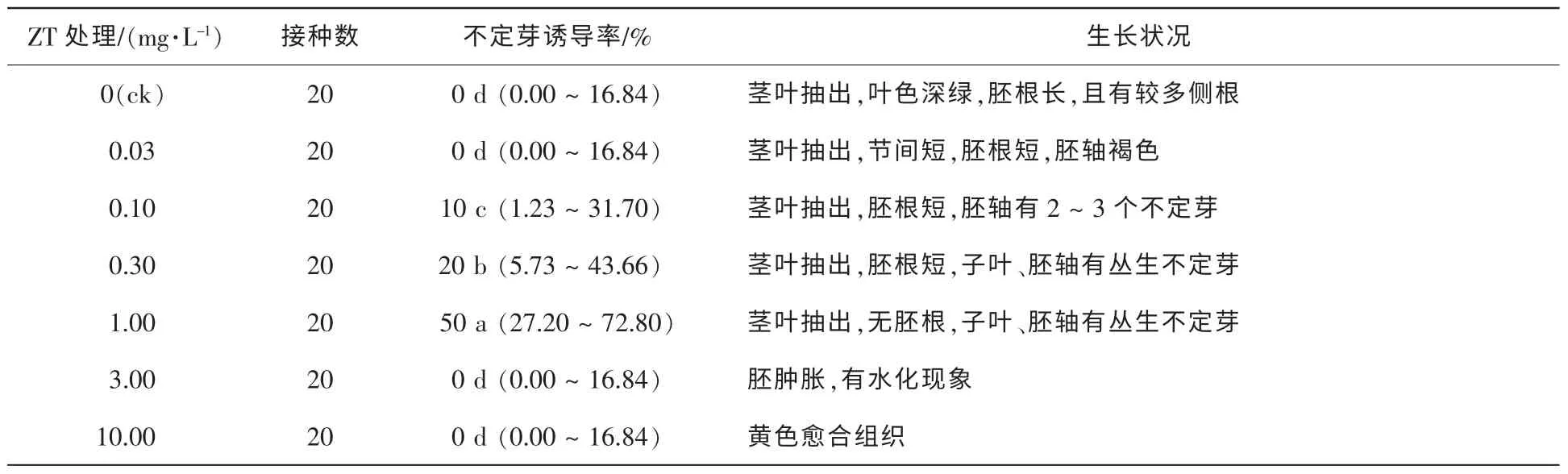
表4 ZT對種胚培養誘導不定芽分化的影響Table 4 Effect of ZT on induction of adventitious buds by embryo culture
2.3 種胚萌動對離體胚培養的影響
由表7可知,將萌動的種胚接種到含0.10~1.00 mg·L-16-BA的培養基上,離體胚分化出大量不定芽;而且隨6-BA質量濃度增高,胚軸分化不定芽的數量呈現先升高后下降的趨勢;但是將未萌動的種胚接種到相同的培養基中,僅在胚軸處產生突起,沒有不定芽的分化。
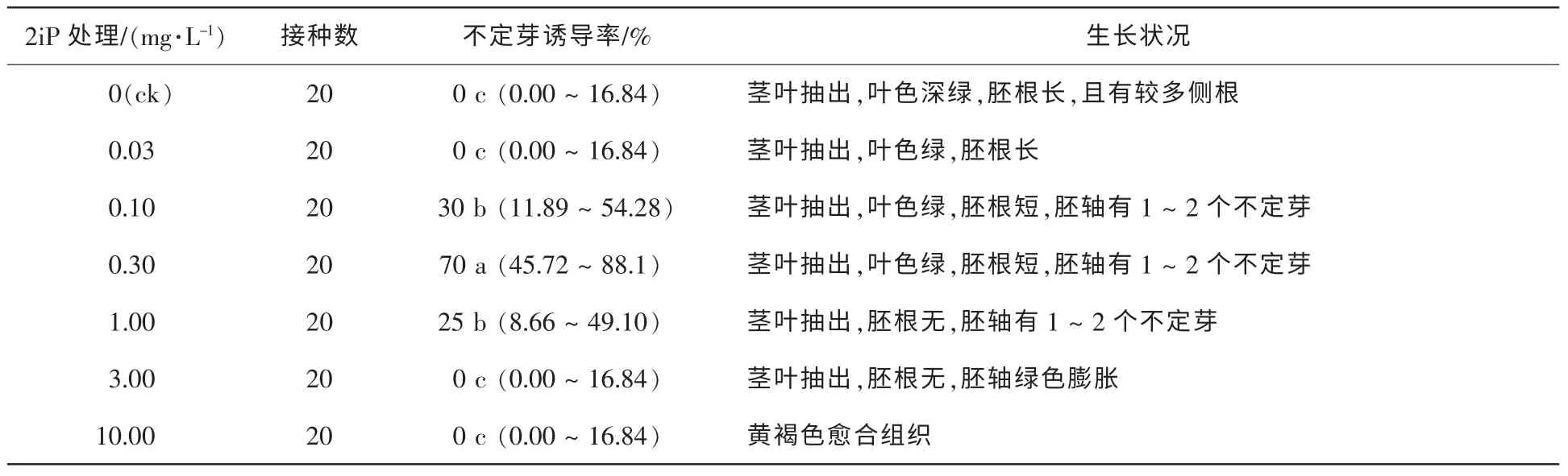
表5 2iP對種胚培養誘導不定芽分化的影響Table 5 Effect of 2iP on induction of adventitious buds by embryo culture

表6 TDZ對種胚培養誘導不定芽分化的影響Table 6 Effect of TDZ on induction of adventitious buds by embryo culture

表7 種胚萌動前后對胚培養的影響Table 7 Effect of with or without breaking seed dormancy on germination of excised embryos
2.4 馴化移栽
將試管苗置于模擬自然環境中,經1個月練苗馴化后,移至混合人工基質中,成活率達到90%。
3 結論與討論
木本植物組織培養的困難之一是建立無菌材料。在建立無菌材料時,外植體表面滅菌將影響試驗成敗。本研究采用25.0 g·L-1次氯酸鈉溶液真空滅菌10 min,污染率僅為6%,出苗率達86%。而且由于南方紅豆杉胚乳較厚,高質量濃度的次氯酸鈉溶液不會傷害到內部的胚,對其萌發無影響。試驗還發現南方紅豆杉種胚在MS培養基上能夠較好的萌發生長,萌發率在95%左右,而陳永勤等[8]曾報道紅豆杉在MS培養基上的萌發較差,這與本研究結果不一致。
臧新等[5]對MS和B5等培養基進行研究,認為這幾種培養基對南方紅豆杉的離體胚培養影響差別不大,而我們在研究中發現不同基本培養基對離體胚培養影響較大。WPM培養基能促進根系產生和莖葉生長,是南方紅豆杉離體種胚培養最適宜的基本培養基。MS培養基對南方紅豆杉莖段生長亦較佳,但不利其根系的生長。B5培養基也不適于南方紅豆杉的胚培養。
在不同細胞分裂素對離體種胚培養的影響試驗中發現,不加細胞分裂素時試管苗生長最好,添加細胞分裂素不利于種胚的伸長生長,但可誘導不定芽的產生且不需經過愈合組織而直接分化產生不定芽,其中KT和ZT最適于南方紅豆杉的不定芽誘導。對于大多數樹種,誘導不定芽時,外植體需接種在細胞分裂素與生長素配比的培養基中,但在許多針葉樹培養中,單獨使用細胞分裂素,足以誘導不定芽發生[9-10],這與本試驗研究結果相一致。
萌動后的南方紅豆杉種子具有較強的生長勢和生理活性,而且此時內源激素含量高于其他時期,而未萌動的種子中發芽抑制物的含量較高[11],抑制種子的萌發生長。南方紅豆杉種胚萌動對胚培養的影響,試驗得知以萌動后的種胚為外植體更容易誘導出不定芽,這可能與南方紅豆杉種胚內源激素隨種胚成熟度而發生變化有關。
[1]程廣有,唐曉杰,高紅兵,等.東北紅豆杉種子休眠機理與解除技術探討[J].北京林業大學學報,2004,26(1):5-10.CHENG Guangyou,TANG Xiaojie,GAO Hongbing,et al.Dormancy mechanism and relieving techniques of seeds of Taxus cuspidata Sieb.et Zucc.[J].J Beijing For Univ,2004,26(1):5-10.
[2]陳永勤,朱蔚華.紅豆杉屬植物的組織、細胞及胚培養[J].植物生理學通訊,1997,33(3):213-219.CHEN Yongqin,ZHU Weihua.Tissue,cell and embryo culture of Taxus[J].Plant Physiol Commun,1997,33(3):213-219.
[3]王水,賈勇炯,魏峰,等.云南紅豆杉的組織培養及植株再生[J].云南植物研究,1997,19(4):407-410.WANG Shui,JIA Yongjiong,WEI Feng,et al.Tissue culture and plant regeneration of Taxus yunnanensis[J].Acta Bot Yunnan,1997,19(4):407-410.
[4]臧新,呂曉輝,楊冬之,等.激素對2種紅豆杉離體胚培養的影響[J].華中農業大學學報:自然科學版,2006,25(3):310-312.ZANG Xin,Lü Xiaohui,YANG Dongzhi,et al.Effects of hormones on the embryo culture of two Taxus species in vitro[J].J Huazhong Agric Univ,2006,25(3):310-312.
[5]臧新,呂曉輝,楊冬之,等.2種紅豆杉的離體胚培養[J].鄭州大學學報:理學版,2006,38(2):107-112.ZANG Xin,Lü Xiaohui,YANG Dongzhi,et al.The embryo culture in vitro of two Taxus species[J].J Zhengzhou Univ Nat Sci Ed,2006,38(2):107-112.
[6]COLQUHOUN D.Lectures on Biostatistics[M].Oxford:Clarendon Press,1971.
[7]SNEDECOR G W.Statistical Methods[M].4th Ed.Iowa:Iowa State College Press,1946.
[8]陳永勤,戴均貴,朱蔚華.紅豆杉成熟胚的離體培養[J].植物生理學通訊,1998,34(3):191-193.CHEN Yongqin,DAI Jungui,ZHU Weihua.In vitro culture of mature embryos of Taxus chinensis[J].Plant Physiol Commun,1998,34(3):191-193.
[9]成小飛,花曉梅,李文鈿.馬尾松離體培養條件下的微繁殖和菌根的形成[J].林業科學研究,1995,8(3):241-246.CHENG Xiaofei,HUA Xiaomei,LI Wendian.Micropropagation and mycorrhizae formation of Pinus massoniana Lamb.in vitro[J].For Res,1995,8(3):241-246.
[10]AITEKEN J,HORGAN K J,THORPE T A.Influence of explant selection on the shoot forming capacity of juvenile tissue of Pinus radiata[J].Can J For Res,1981,11(1):112-117.
[11]張艷杰,高捍東,魯順保.南方紅豆杉種子中發芽抑制物的研究[J].南京林業大學學報:自然科學版,2007,31(4):52-56.ZHANG Yanjie,GAO Handong,LU Shunbao.Germination inhibitors in methanol extract from Taxus chinensis var.mairei seed[J].J Nanjing For Univ Nat Sci Ed,2007,31(4):52-56.
猜你喜歡
中學生數理化·八年級物理人教版(2022年3期)2022-03-16 05:55:08
中學生數理化·八年級物理人教版(2021年12期)2021-12-31 03:23:08
當代陜西(2021年2期)2021-03-29 07:41:24
小讀者(2021年2期)2021-03-29 05:03:48
中學生數理化·中考版(2020年10期)2020-11-27 01:59:48
華人時刊(2019年13期)2019-11-17 14:59:54
中國生殖健康(2019年2期)2019-08-23 08:12:08
文苑(2018年22期)2018-11-19 02:54:14
紅領巾·萌芽(2016年1期)2016-09-10 07:22:44
中國塑料(2016年3期)2016-06-15 20:30:00
