蘇云金芽孢桿菌生物殺蟲劑發酵生產的影響因素及其工藝選擇
2010-08-29 06:56:12常明孫啟宏周順桂倪晉仁
生態環境學報 2010年6期
常明,孫啟宏,周順桂,倪晉仁
1. 中國環境科學研究院,北京 100012;2. 北京大學環境工程系//教育部水沙科學重點實驗室,北京 100871;3. 廣東省農業環境綜合治理重點實驗室//廣東省生態環境與土壤研究所,廣州 510650
蘇云金芽孢桿菌(Bacillus thuringiensis, Bt)簡稱蘇云金桿菌,是一類能在代謝過程中產生芽孢和伴孢晶體的革蘭氏陽性細菌,其制劑是目前世界上產量最大的微生物殺蟲劑[1]。自1901年日本學者石渡繁胤(Ishiwata)從病死家蠶中分離出第一株蘇云金芽孢桿菌以來,全世界已分離出了 71個血清型86個亞種。其殺蟲譜已由無脊椎動物節肢動物門中的鱗翅目擴大到雙翅目、鞘翅目、直翅目等9個目的昆蟲;同時還發現對螨類、線形動物門中的動植物寄生線蟲、原生動物門中的鞭毛蟲、變形蟲和草履蟲以及扁形動物門中的扁蟲、吸蟲、絳蟲等有特異毒性的菌株[2]。目前不少亞種已被成功應用于農業,用來防治糧食作物、蔬菜、棉花、大豆、煙草、果樹等多種害蟲。
Bt制劑的主要殺蟲活性成分是伴孢晶體(亦稱殺蟲晶體蛋白或 δ-內毒素)。殺蟲晶體蛋白(Insecticide Crystal Proteins, ICPs)可占培養物生物量總干重的 20%~30%,分子量在 27~150 kDa之間[3]。Bt的殺蟲機制為:伴孢晶體被昆蟲吞食后,在中腸的堿性環境和蛋白酶的作用下δ-內毒素被分解與激活,成為具有殺蟲活性的毒性肽。毒性肽與幼蟲腸上皮細胞膜的專一受體結合,導致膜穿孔,腸道內溶物滲入血腔,同時芽孢趁機侵入增殖,引起足以致死的敗血癥,導致昆蟲全身麻痹或痙攣而死[4]。近幾十年來,人們對Bt的發酵生產展開了大量研究,并已形成了較為成熟的生產工藝。本文提煉了國內外眾多研究者的實踐經驗,希望通過對發酵影響因素的分析與發酵工藝發展的探討為 Bt生物殺蟲劑的進一步開發與應用提供參考。
1 蘇云金桿菌發酵生產的影響因素
1.1 培養條件
1.1.1 溫度
Bt的生長代謝與溫度密切相關。溫度直接影響細胞內酶的形成與活性,關系到細胞對營養物質的吸收與利用,并影響到菌體數量與伴孢晶體質量。大量研究表明,Bt的最適生長溫度為28~32 ℃,發酵時溫度過高,會導致Bt毒素基因丟失或蛋白失活,毒效降低;溫度過低,則生長緩慢,發酵周期延長[5]。另外,細胞生長最適溫度與晶體蛋白中各基因合成的最適溫度有時會存在一定差異。Melek ?zkan等人考查了溫度對Bt subsp. israelensis HD500蛋白基因合成的影響,結果發現Cry4Ba在25 ℃時含量最高,而菌體生長及Cry11Aa合成的最適溫度分別是37 ℃和30 ℃[6]。不同蘇云金亞種甚至同一亞種的不同菌株,其最佳發酵溫度亦有所不同,發酵時需針對菌株特性選擇最具經濟效率的溫度。
1.1.2 pH值
pH不僅影響芽孢的萌發,也影響芽孢的形成。研究表明,初始pH值為7.0左右時,芽孢萌發率最高,當pH值<6.5或>8.0時,萌發率均在45%以下[1]。發酵過程中,Bt先利用碳源增殖,糖代謝產生大量有機酸,pH下降,隨后在酶的作用下,有機酸進一步被異化,pH值逐漸回升,芽孢與晶體開始形成。伴孢晶體的蛋白質轉化合成過程需要酶的參與,而這種酶的合成與酶活最適pH均為中性,pH值在6.5以下或9.1以上該酶的活性急劇下降。若pH不能回升,芽孢和晶體難于形成[7],若pH值過早升高,又會使營養體增殖提前結束,影響到芽孢和晶體的產量。另外,過高的堿性對伴孢晶體還有一定的溶解作用,會降低發酵液的毒效。發酵過程中可以流加緩沖液,或采用酸堿進行調解,以控制最佳pH值。Khanh Dang Vu等人采用NaOH/H2SO4、NaOH/CH3COOH、NH4OH/CH3COOH和NH4OH/H2SO44種酸堿組合對淀粉廢水培養Bt var.kurstaki HD-1的發酵過程進行調節,使pH值保持在7.0±0.1,結果發現以NaOH/H2SO4和NH4OH/H2SO4為pH調節劑對堿性蛋白酶和淀粉酶的產生十分有利,而以NaOH/CH3COOH為pH調節劑可顯著提高晶體蛋白產量和毒效[8]。由此可見,采用適宜的pH調節劑,對Bt的發酵過程進行有效調控,是Bt發酵生產的關鍵因素。
1.1.3 攪拌速度與通氣量
Bt是一種好氣性芽孢桿菌,在分解和利用發酵基質以及合成自身物質時要消耗大量氧。如:1摩爾葡萄糖完全氧化為CO2和H2O需要6摩爾氧。發酵培養基中糖的含量可達1%,但氧的含量卻很低,飽和濃度也只有0.007%。因此隨著細菌的生長,必須不斷補充大量的氧才能滿足菌體的正常生長。同時氧的存在也可提供一個較高的氧化還原電位,維持細胞內氧化酶系的活性[1]。毒素合成機制也受供氧影響[9],Foda等人通過改變裝液量考查了供氧對Bt subsp. entomocidus HD-635的影響。發現供氧受限時(空氣:培養基=3∶2)菌體不易存活且難于產生芽孢[10]。工業發酵通常以每分鐘通入的空氣體積來衡量通氣量。在液體深層發酵過程中,攪拌速度和通氣量顯得尤為重要,攪拌速度通常為400~600 r·min-1,通氣量一般控制在 1∶0.6~1.2(發酵培養基體積與每分鐘通入空氣的體積之比)。通氣量過大易產生大量泡沫,從排氣口沖出,導致污染,同時也影響氧傳遞,因此在發酵液中添加消泡劑是十分必要的。在發酵的不同階段,通氣量可以適當調節。發酵前期,菌數低,耗氧少,通氣量為1∶0.6~1.0即可;進入對數期,耗氧增大,通氣量需提高為1∶1.0~1.2[1]。同時配合攪拌以強化供氧,為防止細胞受損,不宜采用剪切式攪拌。當芽孢形成以后,雖然菌體代謝活性降低,但氧對芽孢的成熟和伴孢晶體的形成有重要影響,因此后期仍應保持較大的通氣量,可略低于對數期通氣量[9]。
1.1.4 接種
工業生產過程中通常采用3級發酵培養,即搖瓶→種子罐→發酵罐。從種子罐到發酵罐通常有 2種接種方式:芽孢接種和營養體接種。芽孢接種可殺滅雜菌及噬菌體,獲得較高的同步率,但芽孢萌發慢,延遲期較長。嚴格控制滅菌條件,避免污染,采用活性營養體接種芽孢產率可能更高。Pearson和Ward[11]在研究Bt subsp. israelensis的接種條件時,選擇有抑制芽孢形成能力的培養基作為種子培養基,這種營養體接種方式能有效提高同步率與細菌總數,芽孢產率在90%以上。Bt對培養基的適應能力不同,在發酵培養基中馴化后再作為種子接種往往能縮短延遲期[12]。另外 Wu和 Chen[13]在研究中還發現接種時機能明顯影響毒素產量,在菌體活性最強時(延遲期末,對數期初)接種,Bt subsp.darmstadiensis HD-199的外毒素產量比指數末期接種提高了3倍。
1.2 培養基組分
1.2.1 碳源
Bt是一類化能有機營養型細菌,只能利用有機含碳化合物作為碳源和能源。自然界的含碳化合物種類繁多,并非都能被 Bt利用,常用作碳源培養Bt的主要是糖類[1]。單糖如葡萄糖可直接被 Bt吸收,多糖需經菌體產生的胞外淀粉酶水解成葡萄糖等單糖再被吸收。培養基中含有適量葡萄糖,可以促進菌體生長,縮短發酵周期。但在工業生產中培養基滅菌時,受Fe2+、PO43-的影響,葡萄糖易焦糖化,需單獨滅菌,使操作過程變得復雜[14];而且由于葡萄糖代謝產酸,大量葡萄糖會使pH值迅速下降,產生碳源抑制,使代謝進程受阻[15]。因此淀粉、糊精、糖蜜和甘油等多糖都被用作 Bt發酵的常用碳源,它們可以克服葡萄糖代謝過快的弊病,保持發酵后期有一定的糖源。Yasemin Icgen等人考查了各種碳源對 Bt的影響,研究發現蔗糖、乳糖和菊粉作碳源時毒效較高,同時也支持芽孢形成,乳清及糖蜜對毒素合成有促進作用,而其他碳源包括葡萄糖、甘油、麥芽糖、淀粉和糊精都得到較低的毒素量[16];Melek ?zkan等人則發現菊粉、糊精、麥芽糖、乳糖、蔗糖、乳清、甘油等對 Bt subsp. israelensis HD500的毒效有促進作用,而葡萄糖、淀粉和糖蜜用作碳源時毒素產量低[6]。可見不同菌株對碳源種類及營養需求存在差異。
1.2.2 氮源
工業上常用的氮源有黃豆粉、棉籽餅粉、花生餅粉、玉米漿等,其中玉米漿還有較強的緩沖能力,對Bt的生長有利。Salama等人采用棉籽餅粉、魚粉、牛血等農副產品以及一些當地可得的廉價高蛋白廢渣,如飼料酵母、雞舍墊料、各類豆渣等,采用 Bt subsp. kurstaki HD-1、Bt subsp. kurstaki HD-73和Bt subsp. entomocidus等菌株進行發酵[17],結果發現飼料酵母為 Bt生長的最適氮源,三株菌的芽孢數都可達到1×109CFU·mL-1以上,其次為牛血;而棉籽餅粉更適合培養Bt subsp. kurstaki HD-73,雞舍墊料對 Bt subsp. kurstaki HD-1和 Bt subsp.entomocidus生長較為有利。當制劑濃度為500μg/ml時,采用飼料酵母、牛血、雞舍墊料作為氮源的Bt subsp. kurstaki HD-73與Bt subsp. kurstaki HD-1制劑對棉鈴蟲(H. armigera)的致死率均可達80%~100%,以各種豆渣作為氮源培養的Bt subsp.entomocidus對棉貪夜蛾(S. littoralis)的致死率也可達90%左右。可見不同菌株對氮源種類具有選擇性,而不同氮源又影響同一菌株對各類昆蟲的毒效。因此在每一個新菌株用于生產之前,都需要進行最適培養基的選擇。無機氮源通常難以被利用,但新近研究表明,無機氮能增加蛋白水解活性,蛋白水解活性可進一步影響晶體蛋白的穩定性。Yasemin Icgen等人的研究還表明無機氮源能影響Bt的蛋白組成(該菌株可合成135 kDa Cry I及65 kDa Cry II兩種有效蛋白)。當(NH4)2SO4質量濃度從0~2 g·L-1時,65 kDa蛋白隨之濃度增加而提高,用NH4Cl替換(NH4)2SO4則導致135 kDa蛋白含量降低,而改用KNO3可導致65 kDa蛋白消失[18];Melek ?zkan[6]等人也發現(NH4)2HPO4用作無機氮源時能使Cry11Aa和Cry4Ba蛋白產量提高。目前對無機氮源的研究還在不斷深入。
1.2.3 碳氮比
作為微生物生長最重要的營養源,碳、氮源充足是微生物正常生長代謝的必要條件,但同步增加碳、氮源并不能有效提高生物量與毒效,濃度過高還會產生底物抑制,研究發現碳氮比在其中起關鍵作用。首先,碳氮比會明顯影響發酵體系的 pH,高碳氮比培養基的pH在整個代謝過程中不會上升太高,最終停留在7.5左右;而低碳氮比時發酵終點的pH可上升至8.5以上[19]。通常低碳氮比培養基毒素產量略高,但毒素產量還不能完全理解為毒效。Mummigatti和Raghunathan 研究時就遇到了這種情況,盡管低碳氮比時芽孢數及毒素產量略高,但毒效較低;而 Lachhab[21]等人在研究污泥發酵培養Bt HD-1時也發現較高的碳氮比對提高毒效有利,當碳氮比在 8~10左右時,殺蟲毒效較好。因此在選擇碳、氮源時,不可一味追求生物量與毒素產量,需結合生物毒效確定合理的碳氮比。
1.2.4 礦質元素
Bt需要的礦質養料可分為常量元素和微量元素兩大類。常量元素包括磷、鎂、鈉、鉀和鈣,主要參與細胞組成、能量轉移、物質代謝以及調節細胞原生質狀態或細胞通透性等[1]。磷是組成核酸和磷脂的成分,也是蘇云金素的重要組成部分,許多酶的活性基中都有磷的參與。常用的磷鹽主要是K2HPO4和KH2PO4,在提供磷的同時也補充了鉀。Melek ?zkan等人報道較高濃度的無機磷對合成Cry4Ba很有效,當培養基中含50~100 m M K2HPO4時,Cry4Ba和Cry11Aa以及芽孢產率均顯著提高。磷酸鹽對細胞和環境中的酸堿度還起到緩沖作用[6],常用作生測時細胞及晶體的保護劑。鎂是核糖體和膜結構的穩定劑,能激活酶反應,對重金屬引起的細胞毒害還有一定的拮抗作用。Yasemin Icgen等發現鎂影響135 kDa和65 kDa毒性蛋白組分的生物合成,當Mg2+濃度在8×10-5~4×10-3M時,生長、芽孢形成及晶體蛋白合成量都很高[18]。鉀的主要作用是維持滲透壓和離子平衡。Wakisaka等證實在Bt subsp. kurstaki和aizawai培養基中加入鉀鹽比鈉鹽有助于δ-內毒素的形成,當K+濃度達0.20 M時,產量及內毒素活性較高[22]。鈣不參與細胞結構組成,而是以離子狀態控制細胞的生理狀態,如降低細胞質膜的通透性,調節酸堿度,并對一些陽離子的毒性有拮抗作用。與磷類似,鈣鹽也能起到緩沖作用,培養基中加入適量的CaCl2或CaCO3可緩和pH 變化[23]。
微量元素有錳、鋅、硅、銅和鐵,它們多為輔酶或輔基。Bt對微量元素的需要量很少,通常在水和其他養料成分中作為雜質存在的含量足以滿足其需要。鐵是細胞色素、細胞色素氧化酶和過氧化氫酶的活性組成成分;鋅是乙醇脫氫酶的輔基,許多酶的活性也靠鋅激活[1];錳也是多種酶的激活劑,濃度達到10-6M時對Cry4Ba和Cry11Aa毒素產量即有明顯影響[6],Yousten’s培養基中含3×10-4M錳對生長及毒素合成最有利。銅也有一定激活作用,加入10-7M時晶體蛋白合成明顯受到激發,增加至10-6和 10-5之間時,可導致晶體蛋白產量降低,適宜控制銅濃度可使芽孢數比不加銅時提高3倍。此外,Abrosimova等人發現增加礦物鹽的培養基有更好的耐熱性能,這可能是由于鹽組分(尤其是鉀鹽)能明顯提高細胞的護溫屏障[24]。
總之,在 Bt發酵生產中,這些礦質元素的加入量一般為:KH2PO4或K2HPO4為0.075%~0.2%;MgSO4·7H2O 為 0.075%~0.3%;CaCO3為0.075%~0.15% ; MnSO4·H2O 、 FeSO4·7H2O 均 為0.002%。
1.2.5 其它
發酵過程中添加消泡劑等表面活性劑對發酵也有促進作用,Zouari和 Jaoua[25]在培養基中加入了0.5% NaCl和0.1%的Tween-80,發現含固率<59 g·L-1時,Tween-80有助于提高δ-內毒素產量,高于75 g·L-1時,Tween-80在產量提高方面作用不大,但仍能顯著降低蛋白水解活性。總之,Bt的微生物發酵是一個復雜的物質及能量轉化過程,影響因素較多,且各因素間往往相互影響,需綜合調配。表1列出了優化培養基組分及培養條件所得部分代表性成果。
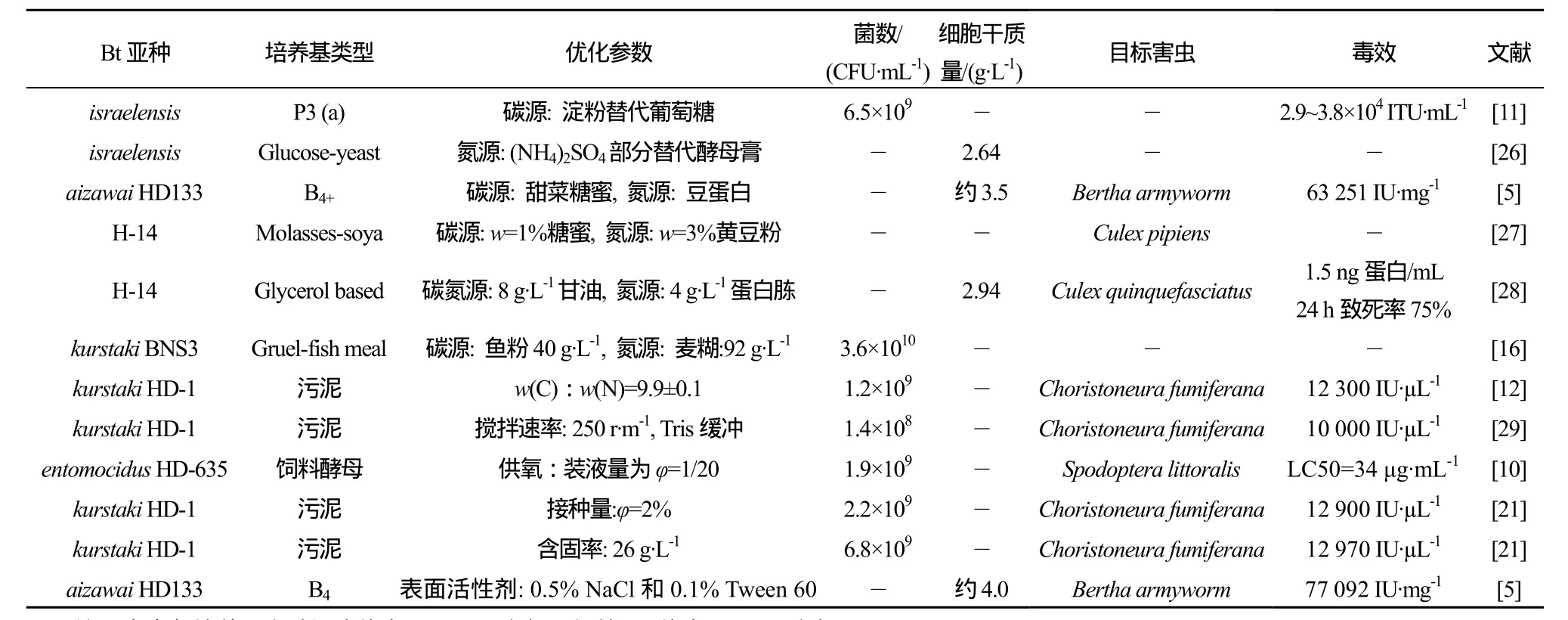
表1 Bt培養條件與培養基組分的優化Table 1 Typical achievements of optimized culture media and culture conditions on Bt fermentation

圖1 液體深層發酵基本工藝流程Fig.1 Schematic diagram showing submerged fermentation process of Bt
2 蘇云金桿菌的發酵工藝選擇
2.1 液體深層發酵:
1956年前蘇聯發表了用液體培養基搖瓶培養Bt,并用于防治菜青蟲的報道[1],從而揭開了Bt液體培養的序幕,Bt制劑之所以能廣泛應用,關鍵在于能通過液體深層發酵大規模生產。其工藝流程如圖1所示。
液體發酵主要有分批發酵、補料分批發酵和連續發酵三種方式[30]。分批發酵一次性投料,工藝簡單,但若要達到較高的發酵水平,需要較高的基質濃度,這種情況下很容易產生基質和代謝產物抑制[31],同時培養基的粘度增加后,由于影響混合和流動而不利于氧的傳遞,最終可能使毒效大打折扣。為此人們從反應器和工藝角度進行了改進。采用外環流氣升式反應器,通過氣體噴射推動液體循環流動以取代傳統的機械攪拌方式[32],由于能耗低、結構簡單、傳質效果好、換熱面積大、剪切力低等優點,對Bt毒效的提高有很大幫助,但目前還缺乏大型生產的經驗,尚處于研究階段;提高攪拌速度或增大通風量以改善供氧環境,有助于毒效的提高[33],但勢必以增加能耗為代價。于是有人提出了流加工藝,逐漸提高基質濃度以削弱抑制,也因此實現了Bt的連續發酵,但長時間的連續發酵培養基很容易染菌,菌種也易發生退化或產生無孢突變株[34]。綜合兩者的優點,補料分批發酵被認為有較好的發展前景,即逐漸補料,一次出料。補料方式又可分為連續式和間歇式,Kang等研究發現間歇式補料比連續式細菌增殖快,芽孢密度大;連續式補料在補料過程中即使細胞增殖緩慢也不會像分批發酵那樣很快轉入芽孢期,補料濃度過高則不能形成芽孢[35]。Zhou等通過控制pH值來調節補料,補料過程中pH保持在7.0左右,避免了營養過剩問題,蘇云金素產量比分批發酵提高了89.51%[35]。
2.2 固態發酵:
固態發酵起源于我國傳統的“制曲”技術,是利用顆粒載體表面所吸附的營養物質或顆粒本身提供的營養來培養微生物[1]。在相對小的空間內,這些顆粒載體可提供相當大的氣液界面,從而滿足好氣微生物增殖所需要的水份、氧氣和營養。20世紀50年代,國外開始將這項技術用于Bt的發酵生產。70年代,我國許多地區與單位都進行了Bt的固態發酵研究,直到上世紀80年代,其生產工藝才逐漸完善。傳統的固態發酵按設計規模可分為網盤薄層法、皿箱式、大池通風法以及地坪式等發酵方式[1],如圖2所示。
可用于蘇云金固態發酵的原料很廣泛,但選擇時既要考慮物料的營養性,也要考慮載體的通氣性。通常使用的載體可分為有機載體和無機載體兩類,有機載體如麥麩、米糠、黃豆餅粉、花生餅粉等,既可作為營養源,又兼有載體作用;無機載體如多孔珍珠巖、細沙等,這些物質通氣性能較好,但需要另外添加營養成分[31]。楊淑蘭等[36]利用麩皮、棉籽餅粉、米糠、草木灰等為發酵原料,進行了百公斤級的Bt subsp. HD-1固態發酵試驗,研究發現種齡、發酵溫度、初始pH值、基質含水量與草木灰含量等是影響芽孢形成和毒效的重要因素,選擇最佳培養條件可使發酵芽孢數穩定在2.0×10 CFU·g-1,在2000倍稀釋度條件下對菜青蟲的致死率為100%。基質的通氣性主要用含水量來控制。適宜的初始含水量,使得培養基有合適的疏松度,顆粒間存在一定空隙,有助于菌體從培養基獲得營養成分和氧的傳遞,從而促進生長繁殖,而過高的含水量會導致培養基粘結成團,多孔性降低,影響氧的傳遞;含水量過低,則使培養基膨脹程度降低,水的活度低,抑制菌體生長。Capalbo等人將潮濕的稻谷裝入聚丙烯袋接種Bt subsp. tolworthi進行了固態發酵實驗室研究,濕度控制在50%~60%,產物田間毒效良好,48 h死亡率可達100%[37]。固態發酵具有低投資、低成本、低排污等優點[38],但因輸送、攪拌、溫度、濕度、pH值和供氧等諸多問題缺少工程解決手段而使其發展受到了限制。陳洪章等人首先提出了壓力脈動固態發酵技術,利用壓力脈動周期刺激強化生物反應和細胞膜傳質速率。壓力脈動避免了機械攪拌的缺陷,提高了傳質傳熱效率,降低溫度、O2和CO2濃度梯度,促進了毒效的提高。目前,壓力脈動固態發酵反應器已成功放大到70 m3的工業級生產規模[39]。
在發酵過程中,液態發酵流動性好,傳質、傳熱性能優于固態發酵,也便于控制;但在后處理過程中,液態發酵通常需要碳酸鈣助濾或離心濃縮,操作復雜且有效成分易流失,而固態發酵可以直接進行干燥、粉碎,能源消耗小,但可濕潤性較差。可見,工藝都存在各自的優缺點,選用何種發酵工藝還應依據培養基組分與發酵水平,以進行綜合調控。
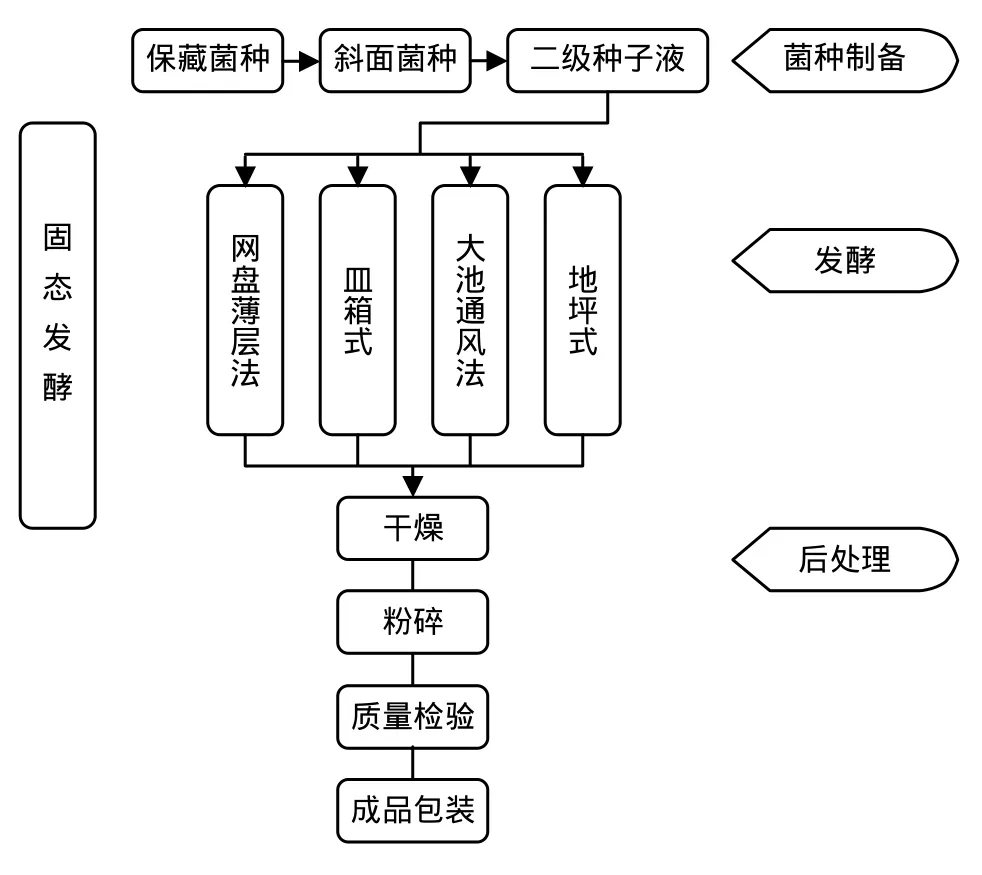
圖2 固態發酵基本工藝流程Fig.2 Schematic diagram solid-state fermentation process of Bt
3 展望
化學殺蟲劑的弊端引發了人們對生物殺蟲劑的關注,而 Bt的研究與應用推動了生物殺蟲劑的發展。據專家預測,今后10年內,由于有機農業、森林防治與生物殺蚊劑市場的需求驅動,生物殺蟲劑產量將以10%~15%的速率遞增[40],Bt生物殺蟲劑的研究還有待進一步深入。
(1)原料價格是 Bt生物殺蟲劑規模化的制約因素,傳統發酵原料價格約占總生產成本的35%~59%[41],因此應致力于開發低成本、當地可得的工農業廢棄物為原料,已有學者曾嘗試以污泥[42,43]、味精廢水[44]、廢啤酒酵母浸出液[45]等為發酵原料,這不僅為生物殺蟲劑的推廣創造了有利條件,還充分利用了廢棄資源。目前,低成本培養原料選擇與優化還處于起步階段,如何放大投產是亟待解決的問題。
(2)傳統生產工藝已不能滿足日益增長的應用需求,高效生產工藝的開發十分迫切。已有研究表明,以提高毒蛋白為目的的高密度補料發酵工藝和壓力脈動固態發酵工藝將有廣闊的應用前景。
(3)為了進一步提高 Bt的殺蟲效果,如延長持效期、擴大殺蟲譜等,可以通過對殺蟲基因的修飾、改造、轉移等基因工程手段構建新型的工程菌[46]。在國外,已有Conder、MVP等10余種Bt工程菌制劑投人了商業應用[47]。在我國,Bt工程菌劑的研發也有擴大趨勢,如高效 Bt和熒光假單胞菌的組合基因工程菌劑WG001已于2000年通過安全性審批,更多新型Bt工程菌劑正有待開發與應用。
[1] 喻子牛. 蘇云金芽孢桿菌制劑的生產與應用[M]. 北京:農業出版社, 1993.YU Ziniu. Production and application of Bacillus thuringiensis preparations[M]. Agricultural Publishing House, Beijing, 1993.
[2] 魯松清, 孫明, 喻子牛, 等. 蘇云金芽孢桿菌殺重晶體蛋白基因的分類[J]. 生物工程進展, 1998, 18(5): 57-62.LU Songqing, SUN Ming, YU Ziniu, et al. Nomenclature of Bacillus thuringiensis Insecticide Crystal Protein Genes[J]. Chinese Progress of Biotechnology, 1998, 18(5): 57-62.
[3] CHATTOPADHYAY A, BHATNAGAR N B, BHATNAGAR R. Bacterial Insecticidal Toxins[J]. Critical Reviews in Microbiology, 2004,30(1): 33-54.
[4] 格拉澤 AN, 二介堂弘. 微生物生物技術: 應用微生物學基礎原理[M]. 陳守文, 喻子牛, 譯. 北京: 科學出版社, 2002.GLAZER A N, NIKAIDO H. Microbial biotechnology: fundamentals of applied microbiology[M]. Chen Shouwen, Yu Ziniu, translators.Beijing: Science Press, 2002.
[5] MORRIS O N, CONVERSE V, KANAGARATNAM P, et al. Effect of cultural conditions on spore-Crystal yield and toxicity of Bacillus thutingiensis subap. azawai (HD133)[J]. Journal of Invertebrate Pathology, 1996, 67: 129-136.
[6] MELEK ?, FILIZ B D, YETIS ü, et al. Nutritional and cultural parameters influencing antidipteran delta-endotoxin production[J]. Research in Microbiology, 2003, 154: 49-53.
[7] 和致中. 碳、氮、磷三因素對蘇云金桿菌的影響[J]. 微生物學通報,1980(7): 7-10.HE Zhizhong. Effects of carbon, nitrogen and phosphorus on Bacillus thuringiensis[J]. Microbiology, 1980(7): 7-10.
[8] VU K D, TYAGI R D, VALéRO J R, et al. Impact of different pH control agents on biopesticidal activity of Bacillus thuringiensis during the fermentation of starch industry wastewater[J]. Bioprocess and Biosystem Engineering, 2009, 32: 511-519.
[9] AVIGNONE-ROSSA C A, ARCAS C, MIGNONE C. Bacillus thuringiensis growth, sporulation and δ-endotoxin production in oxygen limited and non-limited cultures[J]. World Journal of Microbiology and Biotechnology, 1992, 8: 301-304.
[10] FODA M S, SALAMA H S, SELIM M. Factors affecting growth physiology of Bacillus thuringiensis[J]. Applied Microbiology and Biotechnology, 1985, 22(1): 50-52.
[11] PEARSON D, WARD O P. Effect of culture conditions on growth and sporulation of Bacillus thuringiensis subsp israelensis and development of media for production of the protein Crystal endotoxin[J]. Biotechnology Letters, 1988, 10(7): 451-456.
[12] VIDYARTHI A S, TYAGI R D, VALéRO J R, et al. Studies on the production of B. thuringiensis based biopesticides using wastewater sludge as a raw material[J]. Water research, 2002, 36: 4850-4860.
[13] WU W T, CHEN S. An index for determining the culture time of inoculum in cultivation of Bacillus thuringiensis[J]. Biotechnology Letters, 2001, 23: 1201-1204.
[14] 余健秀, 湯慕謹, 徐建敏, 等. 蘇云金桿菌小試生產發酵影響因子的研究[J]. 中國生物防治, 2002, 18 (1): 17-20.YU Jianxiu, TANG Mujin, XU Jianmin, et al. The Factors Affecting the Fermentation of Bacillus thuringiensis[J]. Chinese Journal of Biology Control, 2002, 18 (1): 17-20.
[15] SAKHAROVA Z V, RABOTNOVA I L, KHOVRYCHEV M P. Growth and spore formation in Bacillus thuringiensis var. israelensis and development of media for production of protein Crystal endo-toxin[J].Biotechnology Letters, 1988, 10(7): 451-456.
[16] YASEMIN I, BüLENT I, GüLAY ?. Regulation of Crystal protein biosynthesis by Bacillus thuringiensis: Ⅱ . Effects of carbon and n itrogen sources[J]. Research in Microbiology, 2002, 153: 605-609.
[17] SALAMA H S, FODA M S, Dulmage H T. Novel fermentation media for production of δ-endotoxins from Bacillus thuringiensis[J]. J Invert Pathol, 1983, 41: 8-19.
[18] YASEMIN I, BüLENT I, GüLAY ?. Regulation of Crystal protein biosynthesis by Bacillus thuringiensis: Ⅰ . Effect of mineral elements and pH[J]. Research in Microbiology, 2002, 153: 599-604.
[19] SACHDEVA V, TYAGI R D, VALéRO J R. Factors affecting the production of Bacillus thuringiensis biopesticides[J]. Recent Research and Development of Microbiology, 1999 (3): 363-375.
[20] MUMMIGATTI S G, RAGHUNATHAN N. Influence of media composition on the production of δ-endotoxin by Bacillus thuringiensis var.thuringiensis[J]. Journal of Invertebrate Pathology, 1990, 55: 147-151.
[21] LACHHAB K, TYAGI R D, VALéRO J R. Production of Bacillus thuringiensis biopesticides using wastewater sludge as a raw material:effect of inoculum and sludge solids concentration[J]. Process Biochemistry, 2001, 37: 197-208.
[22] WAKISAKA W, MASAKI E, NISHIMOTO Y. Formation of Crystalline δ-endotoxin or poly-β-hydroxybutyric acid granules by asporogenous mutants of Bacillus thuringiensis. Applied and Environmental Microbiology, 1982: 43.
[23] SIKDAR D P, MAJUMDAR M K, MAJUMDAR S K. Effect of minerals on the production of the delta endotoxin by Bacillus thuringiensis subsp. israelensis[J]. Biotechnology Letters, 1991, 13(7): 511-514.
[24] ABROSIMOVA L I, AVA P V, BREVA G M, et al. Influence of mineral salts on the level of exotoxin production and productivity of a culture of Bacillus thuringiensis[J]. Microbiology, 1986, 55(3): 337-341.
[25] ZOUARI N, JAOUA S. The effect of complex carbon and nitrogen, salt,Tween-80 and acetate on delta-endotoxin production by a Bacillus thuringiensis var. kurstaki[J]. Journal of Industrial Microbiology and Biotechnology, 1999, 23: 497-502.
[26] AVIGNONE-ROSSA C A, YANTORNO O M, ARCAS J A, et al.Organic and inorganic nitrogen source ratio effects on Bacillus thuringiensis var. israelensis delta-endotoxin production[J]. World Journal of Microbiology and Biotechnology, 1990, 6: 27-31.
[27] ABDEL-HAMEED A, CARLBERG G, ELTAYEB O M. Studies on Bacillus thuringiensis H-14 strains isolated in Egypt-Ⅳ . Characteriz ation of fermentation conditions for delta-endotoxin production. World Journal of Microbiology and Biotechnology[J]. 1991, 7(2): 231-236.
[28] MIRTA M F, OSVALDO M Y, HORACIO A, et al. Effect of the media composition on the growth parameters and biological properties of Bacillus thuringiensis var. israelensis delta-endotoxin[J]. World Journal of Microbiology and Biotechnology, 1990, 6: 32-38.
[29] TIRADO M M L, TYAGI R D, VALéRO J R, et al. Production of biopesticides using wastewater sludge as a raw material-effect of process parameters[J]. Water Science and Technology, 2003, 48(8):239-246.
[30] KANG B C, LEE S Y, CHANG H N. Enhanced Spore Production Of Bacillus thuringiensis By Fed-Batch Culture[J]. Biotechnology Letters,1992, 14 (8): 721-726.
[31] 朱瑋, 趙兵, 王曉東, 等. 生物農藥蘇云金芽孢桿菌的研究進展[J].過程工程學報, 2004, 4(3): 282-288.ZHU Wei, ZHAO Bing, WANG Xiaodong, et al. Progress of the Study on Bacillus thuringiensis as Biopesticides[J]. The Chinese Journal of Process Engineering, 2004, 4(3): 282-288.
[32] HUANG T K, WANG P M, WU W T. Cultivation of Bacillus thuringiensis in an Airlift Reactor with Wire Mesh Draft Tubes[J]. Biochemical Engineering Journal, 2001, 7: 35-39.
[33] DHOUHA G, NABIL Z, HASSEN T. Improvement of Bacillus thuringiensis delta-endotoxin production by overcome of carbon catabolite repression through adequate control of aeration[J]. Enzyme and Microbial Technology, 2007, 40(4): 614-622.
[34] ACRAS J, YANTORNO O, ERTOLA R. Effect of High Concentration of Nutrients on Bacillus thuringiensis Culture[J]. Biotechnology Letters, 1987, 9(2): 105-110.
[35] ZHOU Jingwen, CHANG Yafei, XU Zhenghong. Production of thuringiensin by fed-batch culture of Bacillus thuringiensis subsp. darmstadiensis 032 with an improved pH-control glucose feeding strategy[J]. Process Biochemistry, 2007, 42 (1): 52-56.
[36] 楊淑蘭,張怡,梁淑蘭,等. 生物殺蟲劑蘇云金桿菌的固態發酵[J].化工冶金, 1993, 14(2): 162-167.YANG Shulan, ZHANG Yi, LIANG Shulan, et al. Solid fermentation of Bacillus thuringiensis based biopesticide[J]. Journal of Chemical Metallurgy, 1993, 14(2): 162-167.
[37] CAPALBO D M F, MORAES I O, PELIZER L H. Solid-State fermentation of Bacillus thuringiensis tolworthi to control fall armyworm in maize. Electronic Journal of Biotechnology, 2001, 4(2): 112-115.
[38] VIMALA D P S, RAVINDER T, JAIDEV C. Cost-effective production of Bacillus thuringiensis by solid-state fermentation[J] Journal of Invertebrate Pathology, 2005,88(2): 163-168.
[39] 陳洪章, 李佐虎. 固態發酵新技術及其反應器的研制[J]. 化工進展,2002, 21(1): 37-40.CHEN Hongzhang, LI Zuohu. Study on Solid-state Fermentation and Fermenter[J]. Chemical Industry and Engineering Progress, 2002,21(1): 37-40.
[40] TYAGI R D, SIKATI F V, BARNABE S, et al. Simultaneous production of biopesticide and alkaline proteases by Bacillus thuringiensis using sewage sludge as a raw material[J]. Water Science and Technology, 2002, 46(10): 247-254.
[41] The Bacillus thuringiensis Production Handbook[M]. CPL Scientifics Ltd., Newbury, UK: CPL Press, 1993.
[42] 常明, 周順桂, 盧娜, 等. 微生物轉化污泥制備蘇云金桿菌生物殺蟲劑[J]. 環境科學, 2006, 27(7): 1450-1454.CHANG Ming, ZHOU Shungui, LU Na, et al. Bioconversion of sewage sludge to biopesticide by Bacillus thuingiensis[J]. Environ Sci,2006, 27(7): 1450-1454.
[43] BRAR S K, VERMA M, TYAGI R D, et al. Bacillus Thuringiensis Fermentation of Primary and Mixed Sludge: Rheology and Process Performance[J]. Journal of Environmental Engineering, 2008, 134(8):659-670.
[44] 楊建州, 張松鵬. 利用味精廢水發酵生產蘇云金芽孢桿菌的發酵條件研究[J]. 食品與發酵工業, 2002, 28 (4): 28-32.YANG Jianzhou, ZHANG Songpeng. Study on the Conditions of Bacillus thuringiensis Fermentation in Monosodium Glutamate Wastewater[J]. Food and Fermentation Industry, 2002, 28 (4): 28-32.
[45] SAKSINCHAI S, SUPHANTHARIKA M, VERDUYN C. Application of a simple yeast extract from spent brewer's yeast for growth and sporulation of Bacillus thuringiensis: a physiological study[J]. World Journal of Microbiology and Biotechnology, 2001, 17: 307-316.
[46] BAIG D N, MEHNAZ S. Determination and distribution of cry-type genes in halophilc Bacillus thuringiensis isolates of Arabian Sea sedimentary rocks[J]. Microbiological Research, 2010, 165(5): 376-383.
[47] KRIEGER R. Hayes' Handbook of Pesticide Toxicology (Third Edition)[M]. UK: Academic Press, 2010: 119-229.
