產大環糊精4-α-糖基轉移酶的分離純化及其性質研究
2010-08-28 04:42:02冀雪霞王金鵬徐學明金征宇
食品與生物技術學報 2010年3期
關鍵詞:研究
冀雪霞, 王金鵬, 徐學明, 金征宇*
(1.食品科學與技術國家重點實驗室,江南大學,江蘇無錫 214122;2.江南大學食品學院,江蘇無錫 214122)
產大環糊精4-α-糖基轉移酶的分離純化及其性質研究
冀雪霞1,2, 王金鵬1,2, 徐學明1,2, 金征宇*1,2
(1.食品科學與技術國家重點實驗室,江南大學,江蘇無錫 214122;2.江南大學食品學院,江蘇無錫 214122)
4-α-糖基轉移酶能夠作用直鏈淀粉產生大環糊精。作者研究了重組E.coliDH-5α-TA(保藏編號:3093)產生的4-α-糖基轉移酶的分離純化及酶學性質。粗酶液經過65℃處理20 min、Ni -NTA親和層析、生物半透膜脫鹽得到目標酶,該酶經SDS-PAGE凝膠電泳呈單一蛋白條帶,其相對分子質量為57 000。該酶具有較高的轉糖基活性且最小作用底物為麥芽糖。酶學性質研究表明:該酶的最適反應溫度為75℃,最適p H值為7.5;該酶具有良好的耐熱性,在70~85℃酶活維持在80%以上;酶的p H穩定范圍為6.0~8.5。
4-α-糖基轉移酶;大環糊精;分離純化;酶學性質
環糊精(cyclodextrin,CD)是由芽孢桿菌屬(Bacillus)的某些種產生的葡萄糖基轉移酶(CGTase)作用于淀粉而生成的一類環狀低聚糖,其中最常見的是聚合度分別為6、7和8的α-、β-和γ-環糊精[1-2]。區別于上述環糊精的大環糊精是指聚合度從9到幾百不等的環狀葡聚糖[3-4],由于其復雜的空穴結構、高水溶性、低黏度和不回生等特性,可以廣泛地用于食品、化學、醫藥工業中,越來越多的學者開始關注并致力于大環糊精的研究[5-8]。
環糊精的生產通過生物酶法進行的,即由微生物來源的環糊精葡萄糖基轉移酶(簡稱CGTase, EC 2.4.1.19)轉化淀粉而得[2,9-11],其中用于制備大環糊精的酶是4-α-糖基轉移酶(4-α-GTase,EC 2.4.1.25)[12-13]。該酶是一個多功能性酶(Nakamura et al.1994),屬于α-淀粉酶家族[3,14],可以作用直鏈淀粉使其發生環化反應,生成α-1,4-葡聚糖,即大環糊精(Takaha et al.1996,1998)。盡管從眾多微生物或植物中都可以得到該酶[9,15-16,21],但是由于其在生物體內的含量很低,無論是作為研究來源還是用于制備大環糊精,都不能滿足需求[7,17]。國外對產大環糊精的糖基轉移酶的研究已經進行了幾十年,而國內僅近幾年才開始進行研究[4,19],對于產大環糊精酶的研究成果很少。
作者利用實驗室保存的一株基因工程菌E.coliDH-5α-TA發酵產4-α-CGTase,對該酶進行了分離、純化和鑒定,并對其酶學性質進行了初步研究,為該酶進一步的研究以及大環糊精的生產提供了理論依據。
1 材料和方法
1.1 材料
1.1.1 菌種 重組E.coliDH-5α-TA,帶有p6× HTAaGT Ampr質粒(質粒為韓國首爾國立大學Kwan-Hwa Park教授提供)。
1.1.2 培養基LB斜面培養基(g/L):胰蛋白胨10,酵母膏5,NaCl 10,瓊脂20,氨芐青霉素0.1, p H7.5;LB種子培養基(g/L):胰蛋白胨10,酵母膏5,NaCl10,氨芐0.1,p H 7.5;LB發酵基本培養基(g/L):胰蛋白胨10,酵母膏5,NaCl 10,氨芐0.1,p H 7.5。
1.1.3 主要試劑 低相對分子質量標準蛋白:14 400~97 400,分析純,重慶升博科技有限公司產品; SDS-PAGE試劑:分析純,北京經科宏達生物技術有限公司產品;麥芽糖標樣、三(羥甲基)氨基甲烷:分析純,國藥集團化學試劑有限公司產品。
1.1.4 主要設備 SW-CJ-1F雙人單面超凈工作臺:蘇州安泰空氣技術有限公司產品;LS-B50L自動立式電熱壓力蒸汽滅菌鍋:上海華線醫用核子儀器有限公司產品;3K30冷凍離心機:SIGMA公司產品;J YD-650超聲波細胞粉碎機:上海三信儀器有限公司產品;純泰PURIBEST Ni-NTA親和層析柱:生物醫藥純化技術公共服務平臺;SDS-PAGE電泳儀:伯樂公司產品;HPLC:LC-20AT(包括二元泵,手動進樣器,柱溫箱,示差折光檢測器),日本島津公司產品。
1.2 方法
1.2.1 菌體的收集
1)種子培養 將穩定表達的基因工程菌種從平板上挑取單菌落接種至裝有80 mL種子培養基的500 mL三角瓶中培養,設置搖床轉速為200 r/ min,37℃培養12h。
2)搖瓶發酵培養 將培養好的種子培養基按照2%(體積比)的接種量,接種至裝有80 mL發酵培養基的500 mL三角瓶中,設置轉速200 r/min, 37℃發酵培養10 h。
3)收集菌體 將發酵完成的菌液于10 000 r/ min離心10 min收集菌體,用50 mmol/L Tris-HCl (p H 7.5)洗滌細胞兩次,徹底去除殘余培養基,上清液即為粗酶液。
1.2.2 粗酶液的制備 將濕菌體充分均質,冰浴超聲。程序為超聲波功率400 W,處理2 s,間隔5 s,循環100次。然后將其混合液于4℃,10 000 r/ min離心10 min,上清液即為粗酶液[17]。
1.2.3 酶活分析 碘比色法:準確吸取250μL質量分數0.2%直鏈淀粉(DMSO),置入10 mL比色管中,加50μL質量分數1%麥芽糖(Tris-HCl,p H 7.5),600μL 50 mmol/L Tris-HCl(p H 7.5),搖勻。于75℃水浴預熱10 min,準確加入0.1 mL酶液,搖勻,立即計時,于75℃水浴準確保溫酶解反應10 min,立即取出100μL反應液加入到900μL稀碘液,搖勻即可終止反應。以稀碘液為空白,620 nm波長下測吸光度[18]。
酶活定義:在測定條件下,10 min內吸光度每下降0.1為一個酶活單位。
1.2.4 目標酶的分離純化
1)酶的初步純化 將粗酶液置于65℃處理20 min,然后4℃、10 000 r/min離心10 min,取上清液[10],這樣以使其中大部分的不耐熱的酶變性而沉淀,為進一步的分離純化提供了方便。
2)Ni-NTA柱親和層析 所表達的基因工程重組蛋白帶有一個組氨酸標簽,可用Ni-NTA樹脂柱進行親和層析純化。將粗酶液經微孔膜過濾后注入平衡好的Ni-NTA親和層析柱,上樣完畢后靜置2 h,以便His-tag標簽與填料中Ni2+充分結合。先用4 mL洗滌緩沖液[50 mmol/L Tris-HCl(p H 7.5),300 mmol/L NaCl,20 mmol/L咪唑]洗滌過柱以洗去未結合的雜蛋白,再用洗脫緩沖液[50 mmol/L Tris-HCl(p H 7.5),300 mmol/L NaCl, 250 mmol/L咪唑]來洗脫下與親和柱結合的目標蛋白,此時收集目的酶液。
3)透析脫鹽處理 將半透膜制成袋狀,將粗酶液置入袋內,將此透析袋浸入去離子水中,樣品液中的大相對分子質量的生物大分子被截留在袋內,而鹽和小分子物質不斷擴散透析到袋外,用磁力攪拌每隔4 h更換一次去離子水,直到袋內外兩邊的濃度達到平衡為止。
1.2.5 目標酶的鑒定
1)SDS-PAGE鑒定酶的純度和測定酶的分子量 SDS-PAGE凝膠電泳可以鑒定酶的純度并測定其分子量。采用不連續垂直平板電泳系統,對純化后的目標酶進行純度鑒定[18,21]。分離膠質量濃度15 g/dL,濃縮膠質量濃度5 g/dL,電泳采用p H 7.5的Tris-HCL緩沖體系,考馬斯亮藍R-250染色。根據標準蛋白Marker與樣品的SDS-PAGE圖比較來判斷目標酶的純度及其相對分子質量大小。
2)HPLC分析 酶的轉糖基活性表現在能否將一種底物(糖)轉移到另一種底物(糖)分子上。作者用HPLC法在平衡條件下測定底物(葡萄糖、麥芽糖、麥芽四糖混合物)及其酶解產物中各個成分的變化情況,為該酶的作用情況提供參考并為以后酶的應用工作提供了依據和方法。分別向葡萄糖、麥芽糖、麥芽低聚糖中添加等量的酶,75℃保溫20 min,100℃加熱滅活10 min,最后用0.45μm微濾膜過濾后上機測定。色譜條件:采用糖柱,柱溫85℃,流動相為超純水,流速4 mL/min,進樣量10μL,等梯度洗脫。
1.2.6 酶學性質研究
1)不同溫度對其酶活的影響為了確定酶的最適反應溫度,將酶與底物在p H7.5及不同溫度40~100℃條件下反應,反應10 min后分別測其酶活,以最高的酶活力為100%,其余條件下的酶活與其比值換算成相對酶活。
2)不同p H對其酶活的影響由于酶與緩沖液p H有關,所以在分別將緩沖液的p H值調為4.0、5.0、6.0、7.0、8.0、9.0、10.0、11.0時,當p H介于4.0~6.0使用50 mmol/L醋酸鈉緩沖液作為緩沖體系;當p H介于6.0~7.0使用磷酸鈉緩沖液;當p H介于7.0~8.0使用Tris-HCl(p H 7.5)緩沖液作為緩沖體系;當p H介于8.0~10.0使用氨基乙酸-氫氧化鈉緩沖液作為緩沖體系;當p H介于10.0~11.0使用碳酸氫鈉-氫氧化鈉緩沖液作為緩沖體系,在75℃下反應10 min確定各自的酶活,以最高的酶活力為100%,其余條件下的酶活與其比值換算成相對酶活[17]。
3)溫度對酶的穩定性影響在相對穩定的p H 7.5下,使酶在某個溫度下保溫不同時間測定其酶活,以最高的酶活力為100%,其余條件下的酶活與其比值換算成相對酶活。
4)p H值對酶的穩定性影響將酶液分別加入到p H 6.5、7.0、7.5、8.0、8.5、9.0的緩沖體系中室溫放置2 h,然后調回最適pH值,在常規條件反應體系中(75℃)測定殘余酶活,以最高的酶活力為100%,其余條件下的酶活與其比值換算成相對酶活。
2 結果與分析
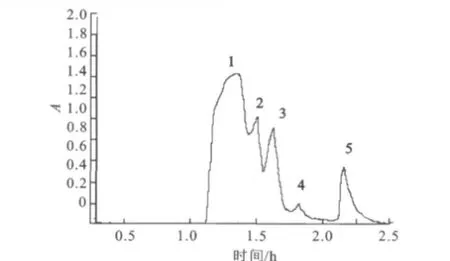
圖1 目標酶的Ni-NTA親和層析柱洗脫曲線Fig.1 Elution curves of purified enzyme on Ni-NTA column
2.1 酶的分離純化
2.1.1 酶的初步純化 由于目標酶耐熱,所以經過65℃保溫20 min處理,可以去除一部分不耐熱的雜蛋白。
2.1.2 Ni-NTA親和層析純化目標蛋白 由圖1中可以看出,粗酶液中蛋白種類較雜,組分較多。經過Ni-NTA親和層析柱分離得到明顯的5個活力峰,即1、2、3、4、5,由1、2、3、4組成的巨大峰為雜蛋白的峰,而5為與Ni-NTA柱耦合后被洗脫液洗洗脫下的目標酶峰。
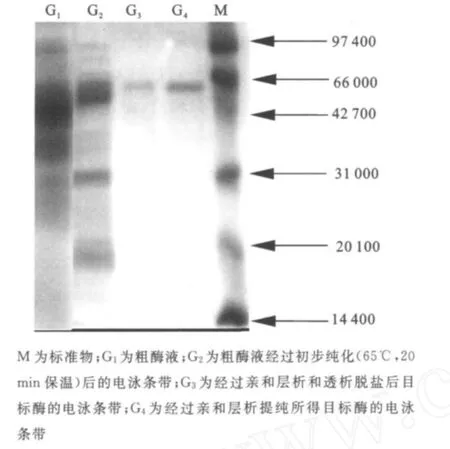
圖2 分離純化的目標酶電泳圖Fig.2 SDS-PAGEanalysis of the purified enzyme
2.1.3 SDS-PA GE分析 分離純化的目標酶經處理后,用SDS-PAGE電泳來對其進行純度與相對分子質量檢測,結果如圖2所示。經過初步純化(65℃,20 min保溫)、親和層析和透析脫鹽,目標酶在SDS-PAGE凝膠中被鑒定為電泳純,相對分子質量57 000。 2.1.4 HPLC分析酶促反應混合物 采用HPLC法對底物及其酶解產物進行分析,結果如圖3~6所示,圖中,A1為葡萄糖,B1為麥芽糖,C1為麥芽低聚糖,D為純酶,A2為經過酶處理的葡萄糖,B2為經過酶處理的麥芽糖,C2為經過酶處理的麥芽低聚糖。圖3為葡萄糖標樣、麥芽糖標樣、麥芽四糖混合物標樣的HPLC圖,由此可知對應組分的保留時間。為其保留時間對應的組分分析。根據保留時間定性,可知葡萄糖為底物反應前后(圖4)無變化;
以麥芽糖為底物反應后(圖5)生成了麥芽七糖、麥芽六糖、麥芽五糖、麥芽四糖、麥芽三糖及葡萄糖;
以麥芽四糖混合物為底物反應后(圖6)生成了麥芽七糖、麥芽六糖、麥芽五糖、麥芽四糖、麥芽三糖、麥芽糖及葡萄糖。
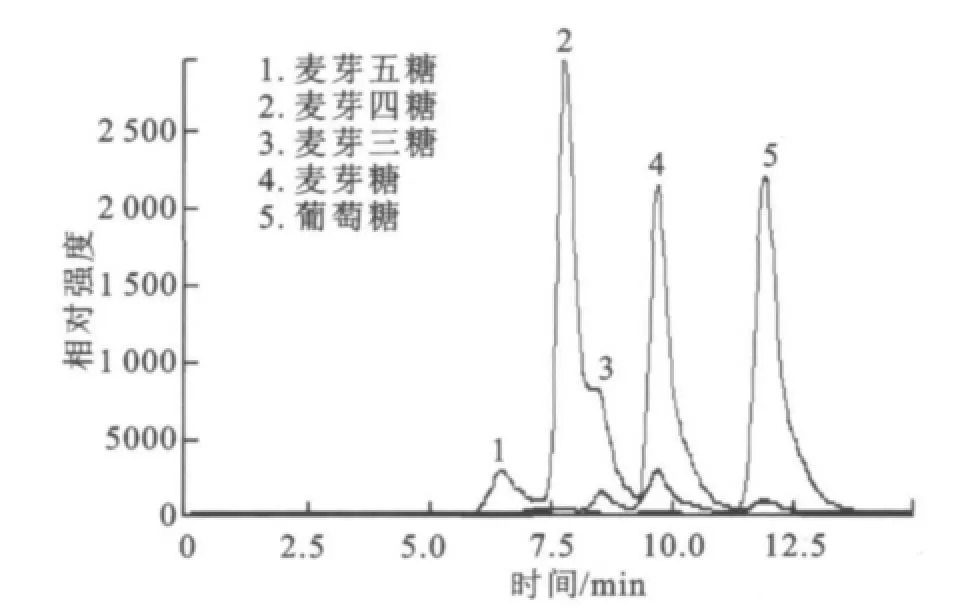
圖3 底物標樣的HPLC圖Fig.3 Standard samples of the substraates
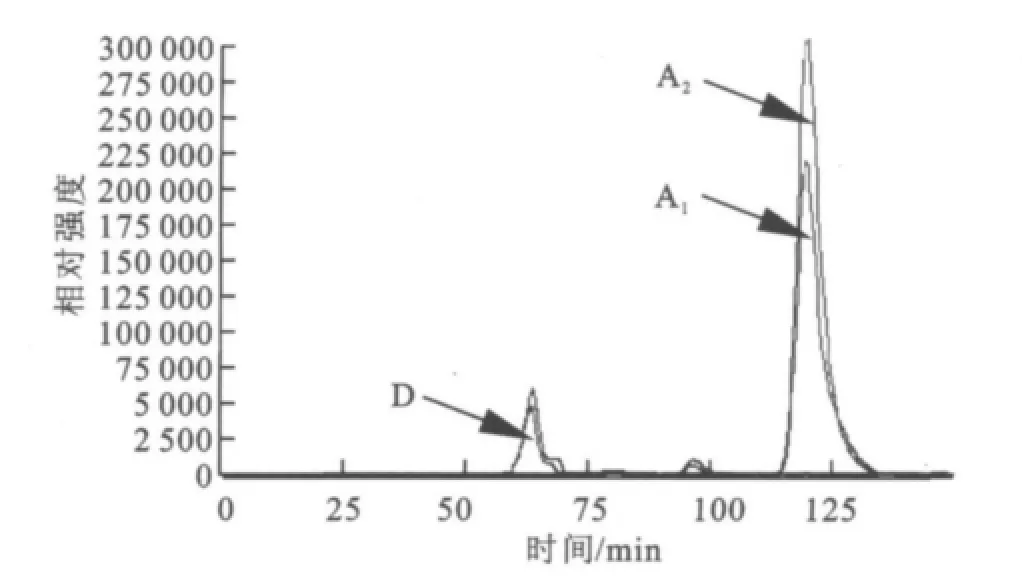
圖4 HPLC分析酶促反應混合物(葡萄糖+純酶)Fig.4 HPLC analysis of the reaction mixture(Glucose +Purified Enzyme)
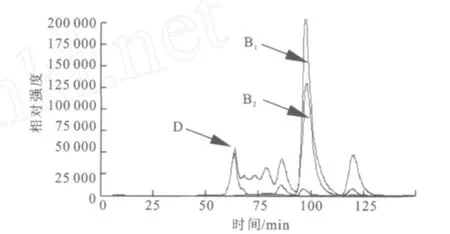
圖5 HPLC分析酶促反應混合物(麥芽糖+純酶)Fig.5 HPLC analysis of the reaction mixture(Maltose +Purified Enzyme)
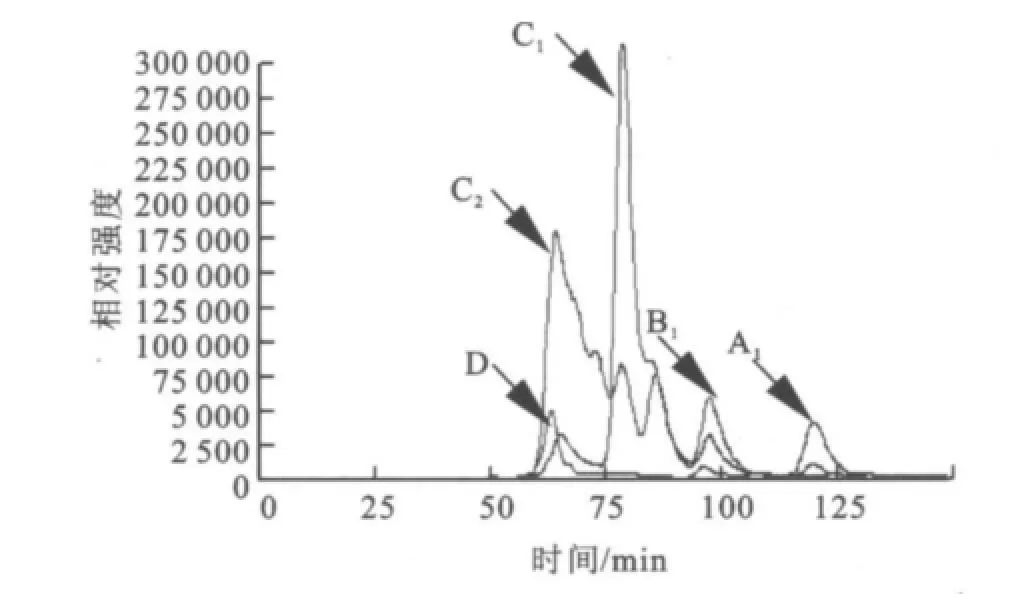
圖6 HPLC分析酶促反應混合物(麥芽低聚糖+純酶)Fig.6 HPLCanalysis of the reaction mixture(Maltotriose&Maltoterraose+Purified Enzyme)
由此可以看出,該酶對葡萄糖沒有轉糖基活性,最小作用底物是麥芽糖。4-α-糖基轉移酶能夠作用麥芽糖及麥芽四糖混合物生成麥芽七糖、麥芽六糖、麥芽五糖、麥芽四糖、麥芽三糖、麥芽糖及葡萄糖等,這表明該酶具有良好的轉糖基活性。
2.2 酶學性質的研究
2.2.1 最適溫度的研究 溫度高于最適溫度時,引起的酶變性而降低反應速度;在溫度低于最適溫度時,酶的反應速度隨著溫度升高而增加,所以為了確定酶的最適反應溫度,研究了一定時間內溫度對酶活性的影響。如圖7所示,該酶隨著溫度的增加酶活呈上升趨勢,約到75℃酶活達到最高,溫度超過75℃后酶活開始下降,得知其最適溫度為75℃,且在50~85℃范圍內酶活都可以達到80%以上,可見該酶為耐熱酶,至100℃時仍剩下20%左右的酶活。溫度對該酶的影響結果與Jin-Hee Park[15]、王水興[21]等人研究基本一致。
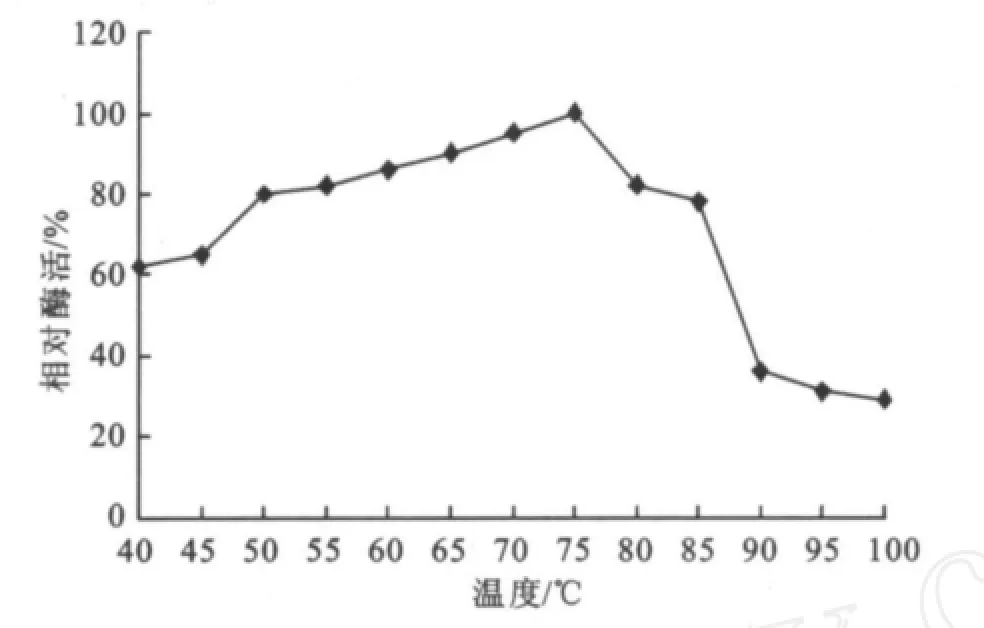
圖7 溫度對酶活力的影響Fig.7 E ffects of temperature on the specific enzymeactivity
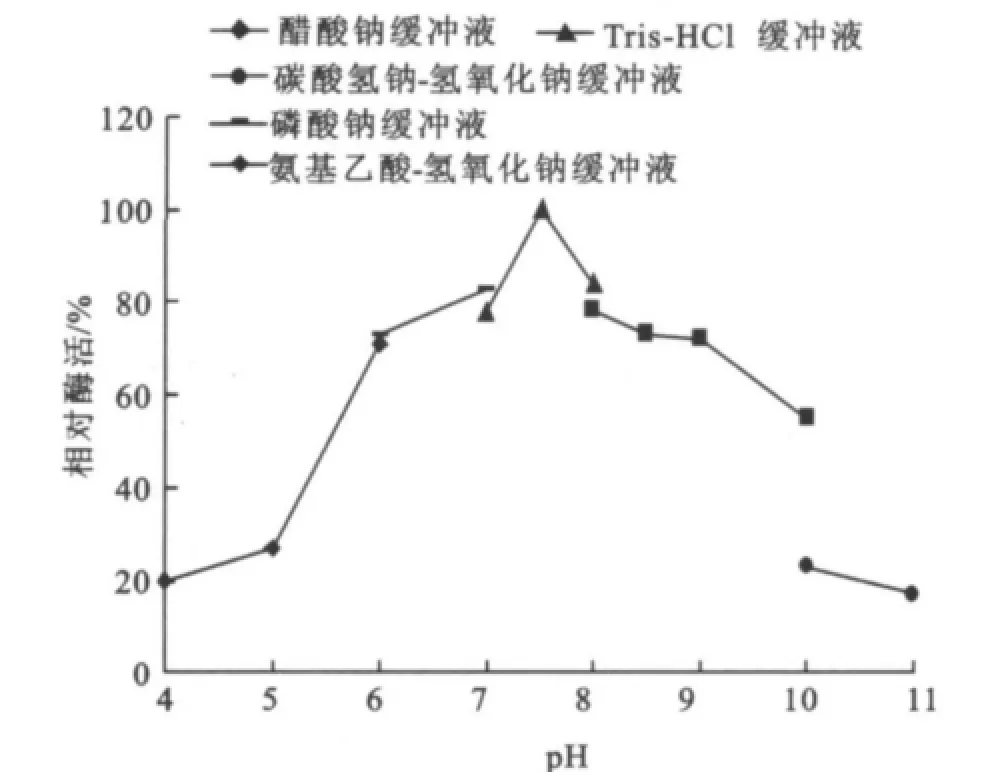
圖8 pH對酶活力的影響Fig.8 Effects of pHon the specific enzyme activity
2.2.2 最適p H值的研究 在溫度為75℃,反應10 min的條件下,測定反應p H值分別為4.0、5.0、6.0、7.0、7.5、8.0、8.5、9.0、10.0、11.0時的酶活,結果見圖8。該酶隨著p H的增加,酶活力呈上升趨勢,在p H值為7.5時酶反應的轉化率最高,然后隨著p H上升酶活開始下降,且在p H 6.0~9.0相對酶活達到80%以上。因此確定其反應最適p H為7.5,在p H 6.0~9.0相對穩定。該結果與菌種的生長特性密切相關。理的酶活為100%,由此確定酶的熱穩定性。由圖9可知,溫度75℃下處理,酶活基本可以保持100%;經80℃處理80 min內都可以基本維持90%左右;當經85℃處理隨著時間的延長酶活逐漸緩慢下降但60 min內都可以維持80%,在保溫80 min殘余酶活力大約為78%;而經90℃處理酶活急劇下降最后在大約57 min后就沒有酶活,而這些充分體現了它在75~85℃范圍內的熱穩定性,與Stefan Jane ek[14]和Jin-Hee Park[15]的研究結果基本一致。
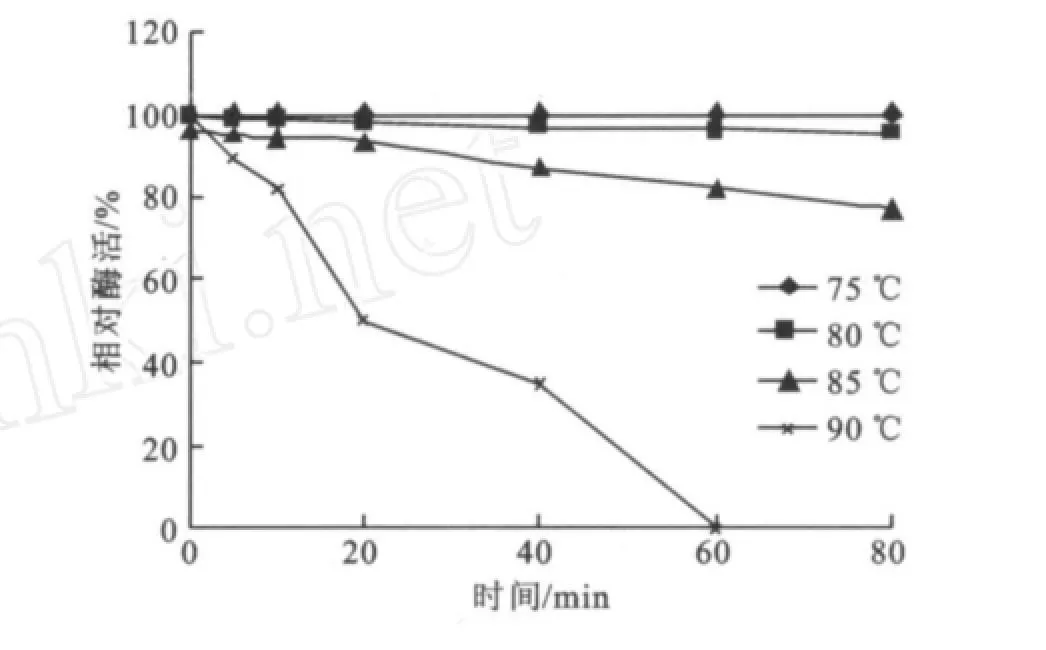
圖9 酶的熱穩定性Fig.9 Effect of temperature on the enzyme stability
2.2.4 酶的p H穩定性研究 如圖10酶的p H穩定性,可見穩定范圍較寬,在p H 6.0~8.5范圍內穩定,殘余酶活力都可以維持在80%左右。
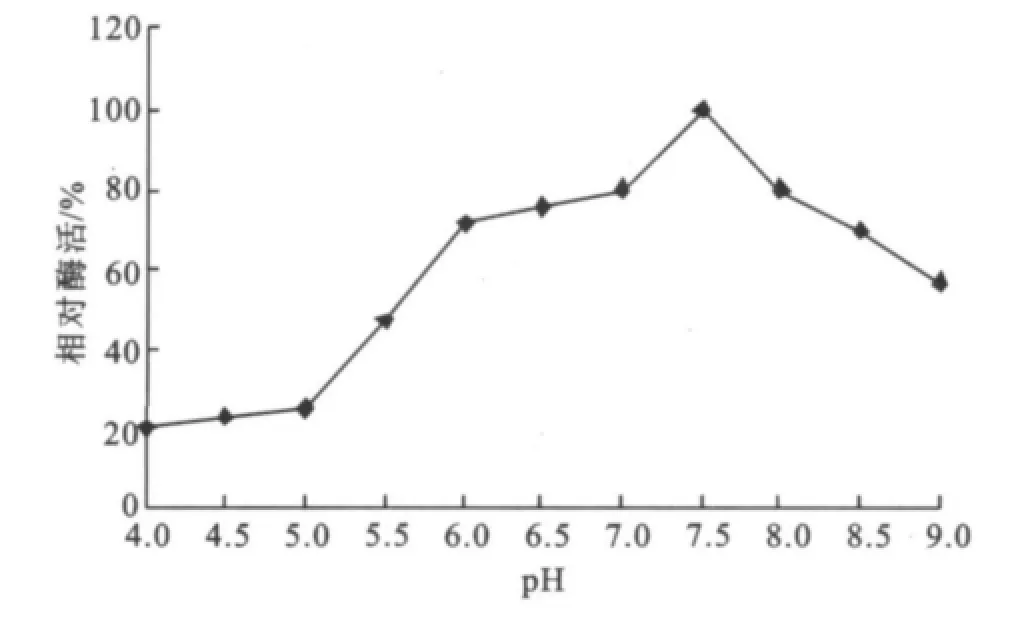
圖10 酶的pH穩定性Fig.10 Effect of pHon the enzyme stability
2.2.3 酶的熱穩定性研究 在相對穩定的p H值下,分別測定酶的熱穩定性。將酶液與50 mmol/L Tris-HCl(p H 7.5)在75~90℃保溫,分別在5、10、20、40、60、80 min取樣測定酶活,以未經保溫處
3 結 語
4-α-轉糖基酶在重組E.coliDH-5α-TA中成功地被表達、分離并鑒定。據現有研究報道,4-α-轉糖基酶具有產大環糊精的功能[15],該實驗用HPLC法證實了該酶的轉糖基活性,且最小的作用單位是麥芽糖,為該酶的進一步轉糖基活性研究奠定了基礎。重組E.coliDH-5α-TA菌株所產的4-α-轉糖基酶有較好的熱穩定性,在70~85℃范圍內基本保持穩定且最適溫度為75℃,適合高溫生產工藝,為進一步的酶的分離純化提供了方便。酶活最佳p H為7.5,在p H在介于6.0到8.5基本穩定。酶的相對分子質量(約為57 000)和酶學性質與報道的芽孢桿菌屬的其他轉糖基酶性質類似。因此本研究為今后對該菌的生產化應用及大環糊精的研究提供了可能,但該菌種的產酶率還偏低,有待進一步提高。
[1]Haruhisa U,Masahiro Wasahiro W,Hiromasa N,et al.Physicochemical properties of large-ring cyclodextrins(CD18-
CD21)[J].Journal of Inclusion Phenomena and Macroccyclic Chemistry,2002,44:403-405.
[2]金征宇,顧正彪,童群義.碳水化合物化學[M].北京:化學工業出版社,2007:126-184.
[3]Qingsheng Q,Tomohiro E,Wolfgang Z.Effect of the reaction temperature on the transglycosylation reactions catalyzed by the cyclodextrin glucanotransferase fromBacillus maceransfor the synthesis of large-ring cyclodextrins[J].Tetrahedron,2004,60(3),799-806.
[4]王水興,郭勇,許楊.大環糊精及其在食品工業應用前瞻[J].食品與機械,2006,22(2),113-116.
WANG Shui-xing,GUO Yong,XU Yang:Large cyclodextrins and their utilization in food industry[J].Food&Machnery,2006,22(2):113-116.
[5]Lambertsen Larsen K,Duedahl Olesen L,Wolfgang Z,et al.Purification and characterisation of cyclodextrin glycosyltransferase fromPaneibacillu sp.F8[J].Carbohydrate Research,1998,310(3):211-219.
[6]Tomono K,Mugishima A,Suzuki T,et al.Interaction between cycloamylose and various drugs[J].Journal of Inclusion Phenomena and Macrocuclic Chemistry,2002,44:267-270.
[7]Meiying Z,Tomohiro E,Wolfgang Z.Synthesis of large-ring cyclodextrins by cyclodextrin glucanotransferases from Bacterial Isolates[J].Journal of Inclusion Phenomena and Macrocyclic Chemistry,2002,44:387-390.
[8]王璋,許時嬰,湯堅.食品化學[M].北京:中國輕工業出版社,1999,45-83.
[9]Shuang Yan T,Eui Jeon W,Kwan Hwa P,et al.Contribution of W229 to the transglycosylation activity of 4-α-glucanotransferase from Pyrococcus furiosus[J].Biochimica et Biophysica Acta,2006,1764:1633-1638.
[10]Shoichi K:Cyclodetrin producing enzyme(CGTase)[M].Process in biotechnology,1996,12:23-41.
[11]Van der Veen B A,Uitdehaag J C M,Dijkstra B W,et al.Engineering of cyclodetrin glycosyltransferase reaction and product specificity[J].Bicochim Biophys Acta,2000,1543(2):336-360.
[12]Terada Y,Yanase M,Takaha T,et al.Cyclodextrins are not the major cyclic alpha-1,4-glucans produced by the initial action of cyclodextrin glucanotransferase on amylase[J].Biological Chemistry,1997(272):15729-15733.
[13]Lambertsen Larsen K.Large cyclodextrin[J].Journal of inclusion phenomena and macrocuclic chemistry,2002,43:1-13. [14]Stefan Jane ek.α-amylase family:Molecular biology and evolution[J].Progress in biophysics and molecular biology,1997, 67(1):67-97.
[15]Jin-Hee P,Kwan-Hwa P.Characterization of 4-α-glucanotransferase from Thermus scotoductus and Its Applications to carbohydrate Engineering[D].Korean:Seoul National University,2004.
[16]Takaha T,Yanase M,Takata H,et al.Potato D-enzyme catalyzes the cyclization of amylase to produce cycloamylose,a novel cyclicglucan[J].Journal of Biological Chemistry,1996,271:2902-2908.
[17]莊海寧,金征宇.酶法合成低聚糖和新糖復合物的研究進展[J].食品與生物技術學報,2008,27(2):1673-1689.
ZHUANG Hai-ning,J IN Zheng-yu.Enzymatic systhesization of oligosaccharides and new compounds research[J].Journal of Food Science and Biotechnology,2008,27(2):1673-1689.
[18]Jin-Hee P,Hyun-Jung K,Yung-Hee K,et al.The action mode of Thermus aquaticus YT-1 4-α-glucanotransferase and its chimeric enzymes introduced with starch-binding domain on amylase and amylopectin[J].Carbohydrate Polymers,2007, 67:164-173.
[19]王水興,李燕萍,許楊.基因工程菌E.coliBL21/pET-Dsba-MaIQ發酵產麥芽糖轉糖基酶的研究[J].食品科學,2007,28 (08):285-289.
WANG Shui-xing,LI Yan-ping,Xu Yang,et al.Research on the glycosyltransferase proceduced by clonedE.coli BL21/ p ET-Dsba-MaIQ[J].Food Chemistry,2007,28(08):285-289.
[20]王水興,郭勇,吳凌偉。來自基因工程菌E.coli BL21/p ET-Dsba-MaIQ麥芽糖基酶的分離純化及酶學性質研究[J].食品科學,2007,28(12):258-262.
WANG Shui-xing,GUO Yong,WU Ling-wei,et al.Purification and Characterization of Amylomaltase from Genetic Engineering StrainE.coliBL21/pET-Dsba-MaIQ[J].Food Chemistry,2007,28(12):258-262.
[21]胡靜,陳育如,魏霞.高產環糊精糖基轉移酶的枯草芽孢桿菌選育、產酶和酶學特性[J].食品與生物技術學報,2008,27 (4):1673-1689.
HU Jing,CHEN Yu-ru,WEI Xia.Screening,Optimized Fermentation and Enzymatic Properties of a Bacillus subtilis Producing Cyclomaltodextrin Glucanotranferase[J].Journal of Food Science and Biotechnology,2008,27(4):1673-1689.
(責任編輯:朱明)
Purification and Properties of 4-α-Glucanotransferase Producing Large-Ring Cyclodextrin
J I Xue-xia1,2, WANGJin-peng1,2, XU Xue-ming1,2, J IN Zheng-yu*1,2
(1.State Key Laboratory of Food Science and Technology,Jiangnan University,Wuxi 214122,China;2.School of Food Science and Technology,Jiangnan University,Wuxi 214122,China)
4-α-glucanotransferase has the function of generating large-ring cyclodextrin.The crude enzymes production by the genetic mutant ofE.coli D H-5α-TA(CGMCC No.3093)was separated and purified by high-temperature processing(65℃,20 min),Ni-NTA affinity column and dialysis.The purified enzyme was demonstrated by SDS-PAGE to be a homogeneous protein and the molecular weight was estimated as 57KDa.It was proved to be with the high level of transglycosylation activity by HPLC which was always used to measure the oligosaccharide and its smallest substrate was maltose.The enzyme exhibited high stability at the temperature range from 70℃to 85℃and the p H range from 6.0 to 8.5.The optimal temperature for the enzyme was 75℃and optimal p H was 7.5,respectively.
4-α-glucanotransferase,large-ringcyclodextrin,separationandpurification, enzymatic property
TS 236.9
:A
1673-1689(2010)03-0336-06
2009-07-15
2007國際科技合作計劃項目(2007DFA31120);江蘇省自然科學基金創新學者攀登項目(BK2008003);食品科學與技術國家重點實驗室2008年度目標導向項目(SKLF-MB-200804);江蘇省科技支撐計劃項目(BE2008317)。
*通信作者:金征宇(1960-),男,江蘇揚州人,教授,工學博士,博士生導師,主要從事碳水化合物研究。Email:jinlab2008@yahoo.com
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
遼金歷史與考古(2019年0期)2020-01-06 07:45:20
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
電子制作(2018年11期)2018-08-04 03:26:04
汽車工程學報(2017年2期)2017-07-05 08:13:02
國際商務財會(2017年8期)2017-06-21 06:14:14
電子制作(2017年23期)2017-02-02 07:17:19
