真核細胞翻譯起始因子5A(eIF5A)研究進展
2010-08-09 06:46:42韓英鵬李文濱
東北農業大學學報 2010年8期
關鍵詞:植物
雙 寶,韓英鵬,李 明,李文濱*
(1.東北農業大學生命科學學院,哈爾濱 150030;2.東北農業大學農學院,哈爾濱 150030)
人們對細胞凋亡的研究過程中發現除了一些轉錄因子以外,在植物的木質部的發生和對病原體的過敏反應以及衰老等過程中,細胞的死亡或其他的生理變化還被一些調控翻譯的因子所調節,其中最值得關注的就是真核細胞翻譯起始因子5A(eIF5A)[1]。
真核細胞翻譯起始因子5A(eIF5A)原名是eIF-4D,最早是從兔子網狀紅血球的核糖體上分離純化出的一個分子質量較小(16~18 ku)[2]的真核生物和古生菌當中最保守蛋白之一[3]。它對細胞的持續性增殖起著關鍵性作用。盡管eIF5A在蛋白合成和其他細胞途徑中扮演的確切角色還不清楚,但普遍認同的假說認為,eIF5A是輔助核糖體具有mRNA特異性的翻譯因子[4],能夠刺激核糖體發揮功能[2]。除此之外,還有關于運輸HIV-mRNA和mRNA的穩定性等功能的報道[5]。近年來發現,eIF5A還在細胞衰老和死亡以及一些植物環境脅迫應答方面有一定的作用,從而使人們更加關注eIF5A功能的研究,本文就這方面的研究進展做一綜述。
1 真核細胞翻譯起始因子5A(eIF5A)的激活
未經翻譯后修飾的eIF5A是無活性的,eIF5A是唯一一個被認知的含有翻譯后修飾形成的氨基酸衍生物羥腐氨酸[Hypusine,Nε-(4-氨基丁基-2-羥丁基)]賴氨酸的蛋白質[6]。而羥腐氨酸的合成在基因翻譯后水平上分兩步進行。第一步,脫氧羥腐氨酸合成酶(DHS,EC 2.5.1.46)在以NAD+為輔助因子的情況下,催化多氨基亞精胺的氨基丁基部分轉化成一個ε-氨基酸族的單一保守的賴氨酸殘基(存在于人類eIF5A中的Lys50),從而使其變成脫氧羥腐氨酸;第二步,脫氧羥腐氨酸殘基中加上的4-氨基丁基端鏈上的二號碳在被依賴Fe(II)的脫氧羥腐氨酸羥化酶(EC 1.14.99.29)羥基化,完成羥腐氨酸的合成及eIF5A的激活,如圖1所示[7]。
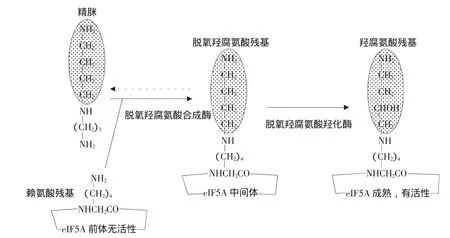
圖1 eIF5A中羥腐氨酸生物合成的途徑Fig.1 Scheme of hypusine biosynthesis in eIF5A
2 真核細胞翻譯起始因子5A(eIF5A)的功能
2.1 起始蛋白質合成與核質轉運
eIF5A最初在一個多核糖體結合的片段上被分離出來,是作為翻譯因子的身份出現的[8]。它可能涉及第一個肽鍵的生成,酵母菌eIF5A在蛋白質合成中起關鍵的起始作用,它將與Met-tRNAi結合的eIF-2帶離48S核糖體,組裝60S核糖體,形成80S核糖體復合體并帶有methionyl-tRNAi[9],隨即起始第一個肽鍵形成[10]。但是近期的體內試驗數據已清楚表明,eIF5A不是大多數蛋白的起始因子。在沒有eIF5A活性的酵母中,并不是所有蛋白質都不能合成,而是蛋白質合成僅下降了30%。這說明翻譯的起始和延長可以在沒有eIF5A或其濃度很低的情況下進行。但是eIF5A缺失的酵母最終是致死的[4-5],一個明顯的缺陷就是它的mRNA受損降解,尤其是那些生活周期短的mRNA[5]。有活性的eIF5A表現出促進特殊mRNA翻譯的作用,DHS被滅活的酵母細胞不能分裂,而只是增大[11]。更多的證據表明,eIF5A通過促進特殊mRNA的核輸出來促進蛋白質的合成[12]。也有人提出eIF5A作用涉及mRNA的轉移[10]。植物eIF5A基因亞型的發現說明了不同的亞型促進了特殊生理學功能所需要的mRNA的翻譯。這些特殊生理學功能中包括光合作用[7]。
有人推測eIF5A可能是細胞核與細胞質之間的轉運蛋白。雖然eIF5A本身沒有核定位信號(NLS),但其C端包含一段富含亮氨酸的結構域,與核運出信號(NES)極為相似[13];電轉化GFPeIF5A瞬時轉染培養細胞后,通過免疫熒光與免疫膠體金電子顯微鏡觀察發現起初eIF5A分布于全細胞,之后逐漸集中于細胞質,突變的eIF5A則分布于整個細胞[14-15],因而認為eIF5A具有核質穿梭功能。也有研究不支持上述觀點,認為eIF5A并不穿梭于細胞核與細胞質之間,可能與ER相互作用影響蛋白質合成[16];在eIF5A細胞亞定位研究中發現,eIF5A并不參與HIV Rev蛋白的核運出[17]。所以eIF5A是否具有核質穿梭能力,目前還沒有定論。
2.2 促進細胞增殖
eIF5A刺激乙硫磷化嘌呤霉素的合成反應的活性是需要羥腐氨酸(Hypusine)的,而乙硫磷化嘌呤霉素的合成反應是測驗第一肽腱形成的模式分析方法[18]。修飾過的eIF5A對哺乳動物[19]、酵母[20]、古細菌[21]的持續性細胞繁殖都很重要。不論對酵母Saccharomyces cerevisiae eIF5A基因(TIF51A和TIF51B)的破壞還是用沉默的eIF5A基因(第50個賴氨酸突變成精氨酸)替換都會導致生長停滯。同樣,單一DHS基因的失活或用編碼失活酶蛋白的基因替換時也會使酵母停止生長,所以eIF5A是細胞增殖所必需的[11]。
DHS有效的抑制劑和脫氧羥腐氨酸羥化酶抑制劑表現出強烈的抑制增殖的作用,并且使包括多種人體癌細胞系在內的哺乳動物細胞停留在細胞周期的某一時期[22]。作為對抗癌藥物干擾素α(IFNα)的反應,羥腐氨酸合成的減弱看上去是伴隨著對細胞調亡的誘導而產生的。此外,DHS的抑制劑,N1-脒基-1,7-二氨基戊烷在抑制細胞生長和引起調亡中表現出同IFNα的協同性[1]。與此同時,這個發現暗示著eIF5A與細胞的增殖和表達失常有著密切的關系,可能從而就導致細胞的癌變。
DHS突變的酵母中,細胞停止生長,細胞體積增大,所以eIF5A被DHS修飾為有活性的形式對細胞增殖是必需的[11]。在對哺乳動物細胞的研究中,羧腐氨酸的合成速率或者合成時間(即eIF5A活化過程)與細胞的分裂進程緊密相關,所以它可能在細胞分裂增殖過程中有著重要作用[8,11]。當eIF5A在酵母細胞中缺失時,G1期捕獲細胞數目增加,哺乳動物細胞中羥腐氨酸累積,使得細胞增殖被抑制[23]。在細胞周期中肌動蛋白的極性化是G1/S轉化期必需的,而被DHS活化的eIF5A參與建立肌動蛋白的極性,就是說eIF5A是細胞周期中G1期到S期轉化的必需的因子之一[24]。
在玉米的卵細胞周期中,eIF5A表達方式也與體細胞周期中的不同:在G0期的卵細胞中高水平轉錄,而得到的懸浮細胞中轉錄水平較低。在體細胞周期中,eIF5A在G1期被強烈誘導,在S、G2和M期持續減少,直到下一個周期的G1期重新誘導[25]。
2.3 調控細胞衰老、細胞死亡及抵抗環境脅迫
在植物中,迄今為止人們已經從擬南芥[26]、紫花苜蓿[27]、煙草[7]、玉米[25]、番茄[28]和水稻[29]中分離了eIF5A基因。已有文獻表明eIF5A的表達存在間歇和短暫的差異,抑制eIF5A的活性將會導致多重作用。轉錄分析表明,煙草中的兩個的eIF5A基因NeIF5A1和NeIF5A2的表達存在差異。NeIF5A1更傾向于在光合作用的組織中表達,而NeIF5A2則是在所有檢測的植物組織中組成型表達[7]。兩種水稻的eIF5A基因,OseIF5A1和OseIF5A2,在水稻的葉子和花序中都表達,而且在衰老的葉子中,這兩種基因都有較高的表達量。另外,在水稻葉子的發育期間OseIF5A1和OseIF5A2都有間歇的調控[29]。近期,人們在番茄組織中發現了eIF5A的表達與細胞的程序死亡存在相關性,所以認為羥腐氨酸修飾的eIF5A可能促進細胞程序死亡所需要的mRNA的翻譯[28]。在衰老的番茄花、子葉、果實以及因環境脅迫而發生細胞程序性死亡的葉片中,DHS和eIF5A及它們的同源蛋白的轉錄水平明顯增加,由此推斷eIF5A可能參與植物自然衰老及環境誘導的細胞程序性死亡[30];在擬南芥上的試驗結果表明,DHS和eIF5A在衰老的葉片中有兩個表達高峰,可能與葉片細胞死亡過程中生物大分子分解及營養物質重新分配有關;通過組成型表達一個反義DHS基因來降低擬南芥中內源DHS蛋白和相應活化的eIF5A的水平,這對擬南芥的生長和發育產生了強烈的影響,例如,延緩了葉子的衰老,增加了抗旱性,增強了種子的產量[26]。此外在成熟的鳳梨果實中檢測到eIF5A的大量表達[31]。Thompson等發現擬南芥中存在3個表達特征不同的eIF5A編碼基因[32],Western雜交分析表明,eIF5A1只在衰老組織中表達,eIF5A2在受機械損傷的組織中高水平表達,而eIF5A3在細胞分裂很活躍的吸脹的種子中高水平表達。過量表達eIF5A3的擬南芥中,花瓣從正常的4瓣變成5瓣,葉子變為并生葉,種子變大,為原來的2倍,這說明eIF5A3可能在發育早期就可促進細胞分裂。這項研究暗示植物中不同eIF5A異構體分別與細胞分裂或細胞死亡有關,即在擬南芥中有兩種eIF5A異構體與細胞死亡有關,而另一種eIF5A與細胞分裂有關,認為eIF5A與植物的生長和發育相關,eIF5A調控植物細胞的衰老。
最近的一項研究表明,擬南芥中三種eIF5A基因之一的擬南芥eIF5A2顯示出能夠調控由有毒力的Pst DC3000侵染所引起的細胞程序死亡。AteIF52組成型過量表達的轉基因植株確實表現出了與早熟型細胞死亡相一致的表型。這些結果說明在植株中,擬南芥eIF5A2是引起細胞程序死亡的信號傳導途徑的一個關鍵因素[33]。
3 結語
eIF5A基因發現至今,一直受到廣泛關注,包括動、植物,人體內的eIF5A不斷被克隆和分析。最受關注的熱點研究方向主要集中在人體eIF5A在癌癥細胞發育中的作用以及相關領域的研究和植物eIF5A基因在植物生長發育中的重要作用以及相關應用方向的初步研究。
就eIF5A的功能而言,以蛋白質合成起始功能為基礎,通過促進一些特異mRNA的轉移及相關特異基因的表達來促進細胞的增殖或者細胞的衰老、死亡,或者產生環境脅迫的應答和抵抗能力。
在很多植物種的研究過程中發現,eIF5A基因往往存在2~4個亞型不等,而初步研究證明其每種亞型在生物體內的功能都有所不同。所以,對每個物種體內的eIF5A基因所有異構體進行克隆和功能的進一步研究,并通過促進某些亞型的表達或者抑制某些亞型的表達,有可能獲得產量提高或者對環境脅迫的抗性強化的品種,實現eIF5A基因在生產實際中的有效應用。
[1]Caraglia M,Marra M,Giuberti G,et al.The eukaryotic initiation factor 5A is involved in the regulation of proliferation and apoptosis induced by Interferon-alpha and EGF in human cancer cells[J].Biochem(Tokyo),2003,133(6)∶757-765.
[2]Kemper W M,Berry K W,Merrick W C.Purification and properties of rabbit reticulocyte protein synthesis initation factors M2Balpha and M2Bbeta[J].Biol Chem,1976,251∶5551-5557.
[3]Gordon E D,Mora R,Meredith S C,et al.Eukaryotic initiation factor 4D,the hypusine-containing protein,is conserved among eukaryotes[J].Biol Chem,1987,262∶16585-16589.
[4]Kang H A,Hershey J W.Effect of initiation factor 5A depletion on protein synthesis and of Saccharomyces cerevisiae[J].Biol Chem,1994,269∶3934-3940.
[5]Zu K D,Jacobson A.A single amino acid substitution in yeast eIF-5A results in mRNA stabilization[J].EMBO,1998,17∶2914-2925.
[6]Caraglia M,Marra M,Giuberti G,et al.The role of eukaryotic initiation factor 5A in the control of cell proliferation and apoptosis[J].Amino Acids,2001,20∶91-104.
[7]Chamot D,Kuhlemeier C.Differential expression of genes encoding the hypusine-containing translation initiation factor,eIF-5A,in tobacco[J].Nucl Acids Res,1992,20∶665-669.
[8]Park M H.The post-translational synthesis of a polyamine-derived amino acid,hypusine,in the eukaryotic translation initiation factor 5A(eIF5A)[J].Journal of Biochemistry(Tokyo),2006,139(2)∶161-169.
[9]Robert E R.Regulation of eukaryotic protein synthesis by initition factors[J].Biol Chem,1993,15∶3017-3020.
[10]Hershey J W.Translational control in mammalian cells[J].Annu Rev Biochem,1991,60∶717-755.
[11]Park M H,Joe Y A,Kang K R.Deoxyhypusine synthase activity is essential for cell viability in the yeast Saccharomyces cerevisiae[J].Biol Chem,1998,273∶1677-1683.
[12]Bevec D,Hauber J.Eukaryotic initiation factor 5A activity and HIV-1 Rev function[J].Biol Signals,1997,6(3)∶124-133.
[13]PollardV W,Malim M H.The HIV-1 Rev protein[J].Annu Rev Microbiol,1998,52∶491-532.
[14]Rosorius O,Reichart B,Kratzer F,et al.Nuclear pore localization and nucleocytoplasmic transport of eIF-5A∶evidence for direct interaction with the export receptor CRM1[J].J Cell Sci,1999,112(Pt14)∶2369-2380.
[15]何昆,周濤,靳寶鋒,等.eIF5A的亞細胞定位研究[J].軍事醫學科學院院刊,2003,27∶330-333.
[16]Shi X P,Yin K C,Waxman L.Effects of inhibitors of RNA and protein synthesis on the subcellular distribution of the eukaryotic translation initiation factor,eIF-5A,and the HIV-1 Rev protein[J].Biol Signals,1997,6(3)∶143-149.
[17]Valentini S R,Casolari J M,Oliveira C C,et al.Genetic interactions of yeast eukaryotic translation initiation factor 5A(eIF5A)reveal connections to poly(A)-binding protein and protein kinase C signaling[J].Genetics,2002,160(2)∶393-405.
[18]Smit-McBride Z,Schnier J,Kaufman R J,et al.Protein synthesis initiation factor 4D∶functional analysis of native and unhypusinated forms of the protein[J].J Biol Chem,1989,264∶18527-18530.
[19]Chen Z P,Yan Y P,Ding Q J,et al.Effects of inhibitors of deoxyhypusine synthase on the differentiation of mouse neuroblastoma and erythroleukemia cells[J].Cancer Lett,1996,105∶233-239.
[20]Magdolen V,Klier H,Lottspeich F.The function of the hypusinecontaining proteins of yeast and other eukaryotes is well conserved[J].Mol Gen Genet,1994,244∶646-652.
[21]Jansson B P M,Malandrin L,Johansson H E.Cell-cycle arrest in archaea by the hypusination inhibitor N1-guanyl-1,7-diaminoheptane[J].Bacteriol,2000,182∶1158-1161.
[22]Clement P M.,Hanauske-Abel H M,Wolff E C,et al.The antifungal drug ciclopirox inhibits deoxyhypusine and proline hydroxylation,endothelial cell growth and angiogenesis in vitro[J].Int J Cancer,2002,100∶491-498.
[23]Wolff E C,Kang K R,Kim Y S,et al.Posttranslational synthesis of hypusine∶evolutionary progression and specificity of the hypusine modification[J].Amino Acids,2007,33∶341-350.
[24]Zanelli C F,Valentini S R.Pkc1 acts through Zds1 and Gic1 to suppress growth and cell polarity defects of a yeast eIF5A mutant[J].Genetics,2005,171(4)∶1571-1581.
[25]Dresselhaus T,Cordts S,L?rz H.A transcript encoding translation initiation factor eIF-5A is stored in unfertilized egg cells of maize[J].Plant Mol Biol,1999,39(5)∶1063-1071.
[26]Wang T W,Lu L,Zhang C G,et al.Pleiotropic effects of suppressing deoxyhypusine synthase expression in Arabidopsis thaliana[J].Plant Mol Biol,2003,52∶1223-1235.
[27]Pay A,Heberle-Bors E,Hirt H.Isolation and sequence determination of the plant homologue of the eukaryotic initiation factor 4D cDNA from alfalfa,Medicago sativa[J].Plant Mol Biol,1991,17∶927-929.
[28]Wang T W,Lu L,Wang D,et al.Isolation and characterization of senescence-induced cDNAs encoding deoxyhypusine synthase and eucaryotic translation initiation factor 5A from tomato[J].Biol Chem,2001,276(20)∶17541-17549.
[29]Chou W C,Huang Y W,Tsay W S,et al.Expression of genes encoding the rice translation initiation factor,eIF5A,is involved in developmental and environmental responses[J].Physiologia Plantarum,2004,121(1)∶50-57.
[30]Xu A,Jao D L,Chen K Y.Identification of mRNA that binds to eukaryotic initiation factor 5A by affinity co-purification and differential display[J].Biochem,2004,384(Pt3)∶585-590.
[31]Moyle R,Fairbairn D J,Ripi J,et al.Developing pineapple fruit has a small transcriptome dominated by metallothionein[J].J Exp Bot,2005,56(409)∶101-112.
[32]Thompson J E,Hopkins M T,Taylor C,et al.Regulation of senescence by eukaryotic translation initiation factor 5A∶implications for plant growth and development[J].Trends Plant Sci,2004,9(4)∶174-179.
[33]Hopkins M T,Lampi Y,Wang T W,et al.Eukaryotic translation initiation factor 5A is involved in pathogen-induced cell death and development of disease symptoms in Arabidopsis[J].Plant Physiol,2008,148(1)∶479-489.
猜你喜歡
少兒科學周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學閱讀指南·低年級版(2021年3期)2021-03-19 06:12:40
小天使·二年級語數英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級語數英綜合(2020年11期)2020-12-16 02:57:22
學苑創造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
