16S rRNA甲基化酶在氨基糖苷類抗生素耐藥革蘭陰性菌中的分布
2010-06-12 01:50:26周穎杰郭慶蘭徐曉剛葉信予王明貴
中國感染與化療雜志 2010年5期
關鍵詞:耐藥
周穎杰, 余 慧, 郭慶蘭, 徐曉剛, 葉信予, 吳 湜, 郭 燕, 王明貴
氨基糖苷類抗生素在臨床廣泛應用,它通過作用于細菌核糖體,抑制其蛋白質合成,并破壞細胞膜完整性而殺滅細菌[1]。由于這類藥物具殺菌作用,抗菌譜廣、有較長的抗生素后效應,與β內酰胺類等抗菌藥物有協同作用,目前是治療革蘭陽性及陰性菌感染,尤其是耐多藥革蘭陰性菌感染的常用聯合治療藥物之一[2]。
2003年,一種由質粒介導的新耐藥機制—16S rRNA甲基化酶(16S rRNA methylase)被發現,該酶使革蘭陰性桿菌對卡那霉素組和慶大霉素組的多種臨床常用氨基糖苷類抗生素高度耐藥。目前已發現的16S rRNA甲基化酶基因有rmtA[3]、rmtB[4]、rmtC[5]、rmtD[6]、npmA[7]和armA[8]。這些基因位于質粒或轉座子上,易于傳播,是造成細菌對氨基糖苷類抗生素高度耐藥的重要原因。然而,16S rRNA甲基化酶系列基因在不同國家和地區的不同細菌中分布可有很大差異,因此有必要進行調查,明確本地區16S rRNA甲基化酶在氨基糖苷類抗生素耐藥菌中的分布,并通過同源性分析,分析該基因的傳播方式,將有助于臨床抗感染治療和新藥開發[9]。
本研究對我院臨床分離的氨基糖苷類抗生素耐藥革蘭陰性菌進行16S rRNA甲基化酶的檢測。
材料與方法
一、材料
(一)菌株來源 收集我院(三級甲等醫院,床位數1 300張)2007年10月1日—12月31日臨床分離的所有連續非重復的革蘭陰性桿菌。藥敏試驗質控菌大腸埃希菌ATCC25922和銅綠假單胞菌ATCC27853為本研究所保存菌株。16S rRNA甲基化酶基因陽性對照株由 Yoshichika教授贈與[3]。
(二)培養基及主要試劑 Mueller-Hinton(MH)瓊脂培養基、阿米卡星和慶大霉素紙片為英國OXOID公司產品,阿米卡星、卡那霉素、妥布霉素、慶大霉素、奈替米星、新霉素、大觀霉素、環丙沙星、頭孢噻肟、頭孢他啶、頭孢吡肟和美羅培南標準品購自中國藥品生物制品檢定所,PCR引物由上海生工生物工程技術服務有限公司合成。
二、方法
(一)藥敏試驗 紙片擴散法藥敏試驗的操作和結果判讀根據CLSI 2007年版[10]。采用瓊脂對倍稀釋法對紙片擴散法提示耐藥的菌株進行阿米卡星等12種抗菌藥的最低抑菌濃度(MIC)測定,操作和結果判讀根據CLSI 2009年版[11]。
(二)16S rRNA甲基化酶基因檢測 瓊脂稀釋法測定對阿米卡星和慶大霉素耐藥的菌株(阿米卡星MIC≥64 mg/L,慶大霉素MIC≥16 mg/L)進行armA,rmtA,rmtB,rmtC,rmtD和npmA基因檢測。制備反應模板采用煮沸法。armA,rmtA,rmtB,rmtC,rmtD基因檢測所用引物見參考文獻[6,12],npmA基因檢測所用引物為 5′-GGAGGGCTATCTAATGTGGT-3′和 5′-GCCCAAAGAGAAT TAAACTG-3′。PCR產物用1%瓊脂凝膠進行電泳,紫外光檢測儀檢測結果。
(三)ERIC(Enterobacterial repetitive intergenic consensus sequence)-PCR分析 ERIC-PCR的引 物 為 5′-AAGTAAGTGACTGGGGTGAGCG-3′[13]。反應條件為94 ℃,5 min;4個循環:94℃1 min,26℃2 min,72℃1 min;40個循環:94℃1 min,40℃1 min,72℃1 min;最后,72℃10 min延伸。反應產物用2%的瓊脂凝膠進行電泳,紫外光檢測儀檢測結果。
(四)統計學 采用SPSS 17.0 for windows進行統計分析。
結 果
一、臨床分離菌株的耐藥情況
共收集741株連續非重復臨床分離的革蘭陰性菌,MIC結果提示其中211株細菌同時對阿米卡星(MIC≥64 mg/L)和慶大霉素(MIC≥16 mg/L)耐藥。
二、甲基化酶基因的檢出
對以上211株耐藥菌進行了PCR檢測。193株(193/211,91.5%)檢出16S rRNA甲基化酶基因,其中armA檢出63.0%(133/211),rmtB檢出28.4%(60/211)。未檢測到rmt A,rmtC,rmtD和npmA基因。
將阿米卡星每個MIC水平的甲基化酶陽性菌株數兩兩做 Fisher′s Exact檢驗,發現256 mg/L與512 mg/L兩組 P<0.01,512 mg/L與 1 024 mg/L兩組,P=0.008,甲基化酶檢出的差異有統計學意義;1 024 mg/L與>1 024 mg/L兩組,P=1,差異無統計學意義(表1)。將慶大霉素每個MIC水平甲基化酶陽性菌株數分別做Fisher′s Exact檢驗,發現32 mg/L與128 mg/L+256 mg/L兩組P=0.028,甲基化酶檢出率的差異有統計學意義,128 mg/L+256 mg/L+512 mg/L與>512 mg/L兩組,P=1,差異無統計學意義(表1)。
104株阿米卡星MIC≥512 mg/L及慶大霉素MIC≥128 mg/L的腸桿菌科細菌中,甲基化酶基因檢出率為100%,armA和rmtB各約占一半(分別為49與55株)(表2);94株阿米卡星MIC≥512 mg/L及慶大霉素MIC≥128 mg/L不發酵糖菌中,甲基化酶基因的檢出率為94.7%,主要為armA(84株)(表2)。
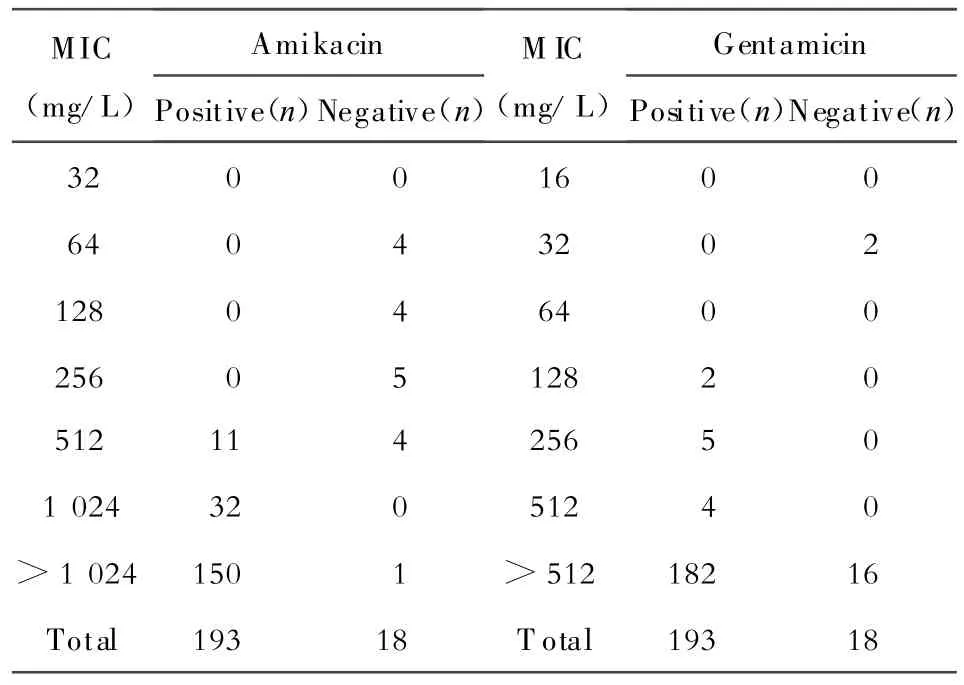
表1 不同阿米卡星和慶大霉素MIC菌株中16S rRNA甲基化酶基因的分布Table 1.The distribution of 16S rRNA methylase genes in strains with different MICs of amikacin and gentamicin
三、藥敏試驗結果
所有含甲基化酶基因的菌株對阿米卡星、卡那霉素、妥布霉素、慶大霉素和奈替米星均呈高度耐藥,但有些菌株對新霉素和大觀霉素敏感(表3)。甲基化酶基因陽性的菌株,阿米卡星的MIC均≥512 mg/L,慶大霉素MIC均≥128 mg/L;甲基化酶陰性的菌株,除5株阿米卡星MIC≥512 mg/L,慶大霉素MIC>512 mg/L,包括1株鮑曼不動桿菌,4株銅綠假單胞菌外,其余13株阿米卡星MIC為64~256 mg/L,慶大霉素MIC 2株為32 mg/L,11株MIC>512 mg/L(表1)。
甲基化酶基因陽性菌株與陰性菌株相比,對頭孢菌素類、氟喹諾酮類及碳青霉烯類抗菌藥物耐藥水平更高(表3)。

表2 對阿米卡星及慶大霉素耐藥革蘭陰性菌中16S rRNA甲基化酶基因的分布(n)Table 2.The prevalence of 16S rRNA methylase genes in Gram-negative strains resistant to amikacin and gentamicin(n)

表3 16S rRNA甲基化酶陽性和陰性菌株對不同抗菌藥物的MIC(mg/L)比較Table 3.MICs(mg/L)of various antimicrobials against 16S rRNA methylase gene positive or negative isolates
四、ERIC-PCR分析結果
198株16S rRNA甲基化酶基因陽性的菌株中,166株成功地進行了 ERIC-PCR分析,其中62株肺炎克雷伯菌分8個克隆,57株鮑曼不動桿菌分7個克隆,19株銅綠假單胞菌分為6個克隆,10株大腸埃希菌分6個克隆,10株奇異變形桿菌分3個克隆,2株摩根摩根菌分2個克隆,4株弗勞地檸檬酸桿菌和2株產酸克雷伯菌分別為1個克隆。同一個ERIC-PCR克隆的菌株可以有不同的甲基化酶基因。
討 論
氨基糖苷類抗生素作用于原核細胞核糖體30S亞單位中16S rRNA的A位(此為mRNA的譯碼區),使細菌在合成蛋白的過程中發生錯誤翻譯,抑制肽鏈延長,造成錯誤的蛋白質插入細胞膜,導致細胞膜斷裂,使細胞內重要物質外漏,細胞迅速死亡[2]。根據在核糖體的作用部位,氨基糖苷類抗生素可以分為:①與A位點結合的4,6-二脫氧鏈霉胺類,其中包括卡那霉素組(阿米卡星)和慶大霉素組;4,5-二脫氧鏈霉胺類,包括新霉素、巴龍霉素等。②與非A位點結合的鏈霉素和大觀霉素[9]。
隨著氨基糖苷類抗生素在臨床廣泛應用,細菌對其耐藥率也持續上升。細菌對氨基糖苷類抗生素的耐藥機制過去認為主要有3種:①細菌外膜蛋白通透性改變或細胞內膜轉運異常,使藥物在菌體內的蓄積減少。②產生氨基糖苷類鈍化酶包括氨基糖苷類乙酰轉移酶、磷酸轉移酶及核苷轉移酶。③核糖體蛋白或16S rRNA突變,使藥物作用靶位改變,藥物進入細菌后不能有效地與核糖體結合而產生耐藥[14-16]。2003年,日本[3]和法國[8]學者發現16S rRNA甲基化酶,該酶使細菌30S核糖體亞單位中16S rRNA A位點的1個或幾個堿基甲基化,使氨基糖苷類抗生素不能與其作用靶點結合,從而介導革蘭陰性桿菌對多種臨床常用氨基糖苷類抗生素耐藥[9]。
自2003年16S rRNA甲基化酶發現以來,該酶在腸桿菌科細菌、銅綠假單胞菌及鮑曼不動桿菌中均有發現[12,17-23]。很多國家及地區對該酶的流行病學進行了研究[3-8],其檢出率在各地區不同菌種及不同耐藥水平的細菌中,從0.03%[12]到95%[17]不等。其分布也有地區及菌種差異:RmtA至今僅在日本的銅綠假單胞菌及韓國的1株銅綠假單胞菌中發現[12,18];RmtD僅在拉丁美洲發現[19-20];鮑曼不動桿菌中僅有ArmA[21-22]。
以上研究中,氨基糖苷類抗生素耐藥菌株數大多少于100株。本研究對211株連續非重復臨床分離的氨基糖苷類抗生素耐藥菌株進行了檢測,其中包括腸桿菌科細菌及不發酵糖菌,有助于了解該酶在氨基糖苷類抗生素耐藥革蘭陰性菌中的流行情況。
研究發現,在阿米卡星MIC≥512 mg/L及慶大霉素MIC≥128 mg/L的菌株中,104株腸桿菌科細菌100%檢出甲基化酶,94株不發酵糖菌95%檢出甲基化酶。這些甲基化酶陽性的菌株對卡那霉素、妥布霉素、慶大霉素和奈替米星亦均呈高度耐藥,但有些菌株對新霉素和大觀霉素敏感,這是由于新霉素和大觀霉素的作用機制是與非A位點核糖體結合,因此不受甲基化酶對A位點甲基化的影響。而對在阿米卡星MIC為64~256 mg/L的13株細菌中,未檢出甲基化酶,這些菌株中2株慶大霉素MIC為32 mg/L,11株MIC>512 mg/L。
我國臺灣Yan等[23]發現,35株armA或rmtB陽性的大腸埃希菌和肺炎克雷伯菌,其阿米卡星MIC≥512 mg/L,而46株未檢出該類基因的細菌MIC為16~64 mg/L,與本研究的發現相似。另外,本研究發現甲基化酶陰性菌株中存在慶大霉素的MIC>512 mg/L的菌株,可能因氨基糖苷類鈍化酶引起[2],需要進一步研究證實。所以,采用阿米卡星的MIC值預測是否存在16S rRNA甲基化酶較合適。
本研究有4株銅綠假單胞菌和1株鮑曼不動桿菌對阿米卡星高度耐藥,但未檢出現有的6種甲基化酶基因,需要對該5株細菌可能存在的其他耐藥機制進一步檢測和研究,以明確其高度耐藥的原因。
至今在中國大陸僅檢測到armA和rmtB[22,24-25],本研究也未發現其他4種甲基化酶基因。與其他文獻報道類似,本研究在鮑曼不動桿菌中只檢出armA[21-22]。目前,rmtA僅在日本和韓國的銅綠假單胞菌中發現[12,18],本研究的42株銅綠假單胞菌中也未發現有rmt A。
不少文獻報道甲基化酶基因通常與其他耐藥基因同時存在,如 blaTEM、blaCTX-M、blaSHV、blaOX A[4,7,21,23,26]、和qnr[24],所以,含甲基化酶基因的菌株可能同時對β內酰胺類和氟喹諾酮類抗菌藥物耐藥。本研究的結果也發現,甲基化酶基因陽性的菌株比陰性菌株對頭孢菌素類、碳青霉烯類和喹諾酮類抗菌藥物的耐藥水平更高。
ERIC-PCR的結果表明,耐藥菌株并非單克隆傳播,因此,推測該基因的傳播同時有水平和垂直2種方式。Chen等[24]的研究已經證實這一點。
綜上所述,幾乎所有阿米卡星MIC>512 mg/L的革蘭陰性菌臨床株中均檢出16S rRNA armA或rmtB甲基化酶基因。
[1] Magnet S,Blanchard JS.Molecular insights into aminoglycoside action and resistance[J].Chem Rev,2005,105(2):477-498.
[2] 汪復,張嬰元.實用抗感染治療學[M].上海:人民衛生出版社,2004:231-232.
[3] Yokoyama K,Doi Y,Yamane K,et al.Acquisition of 16S rRNA methylase gene in P.aeruginosa[J].Lancet,2003,362(9399):1888-1893.
[4] Doi Y,Yokoyama K,Yamane K,et al.Plasmid-mediated 16S rRNA methylase in S.marcescens conferring high-level resistance to aminoglycosides[J].Antimicrob Agents Chemother,2004,48(2):491-496.
[5] Wachino J,Yamane K,Shibayama K,et al.Novel plasmidmediated 16S rRNA methylase,rmtC,found in a P.mirabilisisolate demonstrating extraordinary high-level resistance against various aminoglycosides[J].Antimicrob Agents Chemother,2006,50(1):178-184.
[6] Doi Y,Oliveria Garcia D,Adams J,et al.Coproduction of novel 16S rRNA methylase RmtD and metallo-β-lactamase SPM-1 in a panresistant P.aeruginosa isolate from Brazil[J].Antimicrob Agents Chemother,2007,51(3):852-856.
[7] Galimand M,Courvalin P,Lambert T.Plasmid-mediated highlevel resistance to aminoglycosides in Enterobacteriaceae due to 16S rRNA methylation[J].Antimicrob Agents Chemother,2003,47(8):2565-2571.
[8] Wachino J,Shibayama K,Kurokawa H,et al.Novel plasmid-mediated 16S rRNA m1A1408 methyltransferase,NpmA,found in a clinically isolated E.coli strain resistant to structurally diverse aminoglycosides[J].Antimicrob Agents Chemother,2007,51(12):4401-4409.
[9] 周穎杰,王明貴.質粒介導氨基糖苷類抗生素新耐藥機制:16S rRNA甲基化酶[J].中國感染與化療雜志,2010,10(2):155-159.
[10] Clinical and Laboratory Standards Institute.Performance Standards for Antimicrobial Susceptibility Testing[S].Seventeenth Info rmational Supplement,2007,M100-S17.
[11] Clinical and Laboratory Standards Institute.Performance Standards for Antimicrobial Susceptibility T esting[S].Nineteenth Info rmational Supplement,2009,M100-S19.
[12] Yamane K,Wachino J,Suzuki S,et al.16S rRNA methylase-producing,gram-negative pathogens,Japan[J].Emerg Infect Dis,2007,13(4):642-646.
[13] Versalovic J,Koeuth T,Lupski JR.Distribution of repetitive DNA sequences in eubacteria and application to fingerprinting of bacterial genomes[J].Nucleic Acids Res,1991,19(24):6823-6831.
[14] Hancock RE.Aminoglycoside uptake and mode of actionwith special reference to streptomycin and gentamicin.I.Antag onists and mutants[J].J Antimicrob Chemother,1981,8(4):249-276.
猜你喜歡
保健醫苑(2022年5期)2022-06-10 07:46:38
現代臨床醫學(2022年3期)2022-06-06 07:59:40
昆明醫科大學學報(2022年1期)2022-02-28 07:43:40
天津醫科大學學報(2021年3期)2021-07-21 09:04:02
科學大眾(2020年12期)2020-08-13 03:22:22
云南醫藥(2019年3期)2019-07-25 07:25:10
現代檢驗醫學雜志(2016年1期)2016-11-12 13:19:40
國外醫藥(抗生素分冊)(2016年6期)2016-07-10 11:34:45
中國衛生標準管理(2015年14期)2016-01-15 02:58:37
中國當代醫藥(2015年17期)2015-03-01 02:03:58
