山羊GnRHR基因部分序列多態性及其與產羔數的關聯性分析
2010-01-30 01:32:52張育軍胡敏蘭張子軍丁建平任春環楊玉敏阮崇美
中國草食動物科學 2010年4期
張育軍,胡敏蘭 ,張子軍,丁建平,任春環,楊玉敏,阮崇美
(安徽農業大學動物科技學院,合肥 230036)
下丘腦分泌的促性腺激素釋放激素(gonadotropin releasing hormone,GnRH)在調節哺乳動物生殖系統正常發育及其機能維持方面發揮著重要作用。GnRH與垂體促性腺細胞表面的GnRHR結合后,刺激垂體前葉合成和釋放促卵泡素(follicle-stimulating hormone,FSH)和促黃體素(luteinizing hormone,LH),從而促進性腺的生長、成熟和調控哺乳動物的繁殖力。GnRHR是位于垂體促性腺細胞表面的一種G蛋白耦聯受體,轉導GnRH信號,并調節LH和FSH的合成和分泌[1]。因此,GnRHR基因是動物繁殖性狀的一個重要候選基因。Antelli等報道,位于人GnRHR的第1個胞外環上的T104I和Y108C的突變可影響配體與受體的結合力,從而導致人第二性征的發育障礙[2]。Dunn等報道,母雞群體中GnRHR限制酶等位基因的頻率分別是0.54(Bpu1102I+)和0.46(Bpu1102I-),GnRHR基因對雙黃蛋數目有顯著加性效應(P<0.05)[3]。劉忠慧等在小尾寒羊和陶賽特羊群體GnRHR基因cDNA檢測到第230處發生G→C的突變,導致第77個氨基酸由甘氨酸變為半胱氨酸(Gly→Cys),這一突變可能與小尾寒羊和陶賽特羊的繁殖力有一定的正相關[4]。儲明星等發現濟寧青山羊GnRHR基因具有AA、AB和BB三種基因型,并且BB型濟寧青山羊平均產羔數比AB和AA型分別多0.69只(P<0.05)和0.82只(P<0.05)[5]。而GnRHR基因多態性分析在黃淮山羊方面還未見相關報道。
黃淮山羊是我國優良的地方山羊品種之一,產羔率達227%~239%[6];波爾山羊則是世界著名的肉用山羊品種,產羔率180%~193%。本研究以黃淮山羊和波爾山羊為研究素材,以GnRHR基因為候選基因,采用單鏈構象多態(single strand conformation polymorphism,SSCP)方法對動物繁殖起重要調控作用的Gn-RHR基因進行單核苷酸多態性(single nucleotide polymorphism,SNP)檢測,以比較GnRHR基因在黃淮山羊和波爾山羊品種中的多態性,旨在尋找與產羔數相關的遺傳標記,為山羊高繁殖力的標記輔助選擇提供科學依據。
1 材料和方法
1.1 材料
1.1.1 材料 具有產羔記錄的黃淮山羊母羊120只(安徽省阜陽市穎東區種羊場)和波爾山羊母羊34只(安徽省肥東縣包公鎮種羊場)。每只采血樣10 mL,用ACD抗凝,-20℃凍存。用酚氯仿抽提法提取基因組DNA,溶于 TE,4℃保存。
1.1.2 主要試劑 Taq DNA聚合酶、dNTPs、pGEM2T Easy載體、DNA片段回收純化試劑盒、質粒提取純化試劑盒等均購自上海生工生物工程技術有限公司,N、N2亞甲基雙丙烯酰胺、丙烯酰胺、TEMED、過硫酸胺等購自Sigma公司。
1.2 方法
1.2.1 引物設計和PCR擴增 根據GenBank綿羊GnRHR基因的全編碼序列(L42937,L43841),用Primer Premier 5.0軟件在外顯子1、外顯子2處設計3對引物。第1和第2對引物(P1,P2)用于擴增exon 1片段,而P3引物對用于擴增exon 2片段。引物序列、擴增片段大小見表1。
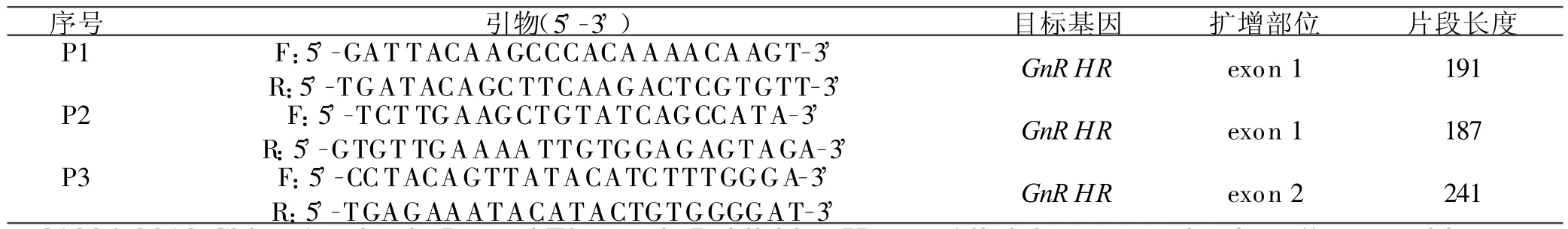
表1 PCR擴增的引物序列
PCR反應總體系為25 μ L,其中基因組DNA 50 ng,10×緩沖 液 2.5 μ L,dNTPs 的 終濃 度 200 μ mol/L,TaqDNA聚合酶1 u。引物的濃度及其PCR擴增條件見表2。產物用1.5%瓊脂糖凝膠電泳檢測。

表2 3對引物PCR擴增的條件
1.2.2 SSCP分析 配制30 mL 12%的聚丙烯酰胺凝膠,混勻后迅速灌膠;當膠灌至離玻璃板上沿0.1 cm時,停止灌膠,插入梳子,室溫聚合;凝膠聚合后,向電泳槽中加1×TBE,用注射器沖洗加樣孔;預電泳10 min,同時準備點樣;取2.0 μ L PCR產物置于 PCR管中,加7 μ L上樣緩沖液,煮沸變性 10 min后迅速置于冰上,冰浴10 min,用微量移液器點樣;150 V電壓,電泳12~14 h。電泳結束后,進行硝酸銀染色。用Alpha Imager TM 2200 and 1220 Documentation and A-nalysis Systems(Alpha Innotech Corporation,San Leandro,CA,USA)拍照和分析。
1.2.3 PCR產物測序 用柱式凝膠回收試劑盒回收純化PCR產物。將回收后的DNA片段與pGEM2T Easy載體連接,轉化大腸桿菌(E.coli)DH5α菌株,酶切鑒定后在3730測序儀上測序。
1.2.4 群體遺傳學研究分析 統計基因頻率及基因型頻率,并運用數學模型進行最小二乘方差分析,比較黃淮山羊產羔數在不同GnRHR基因型之間的差異:yijkl=μ+HYSi+Pj+Gk+eijkl,其中:yijkl為產羔數的記錄值;μ為群體均值;HY Si為第i個場年季的固定效應;Pj為第j個胎次的固定效應;Gk為第k種標記基因型的固定效應;eijk l為隨機殘差效應,且滿足e~N(0,σ2)。用SAS(6.12版本)軟件包的GLM(general linear model)過程完成。
產羔數是指2~4胎繁殖母羊一胎的產羔數量。分別來自2個種羊場2008年后半年的產羔記錄。
2 結果
2.1 PCR擴增 3對引物P1、P2、P3擴增獲得了理想的PCR產物(圖1),片段長度與預期大小一致,并且沒有非特異性擴增條帶,2個品種羊的擴增片段大小也一致,可以直接進行SSCP分析。
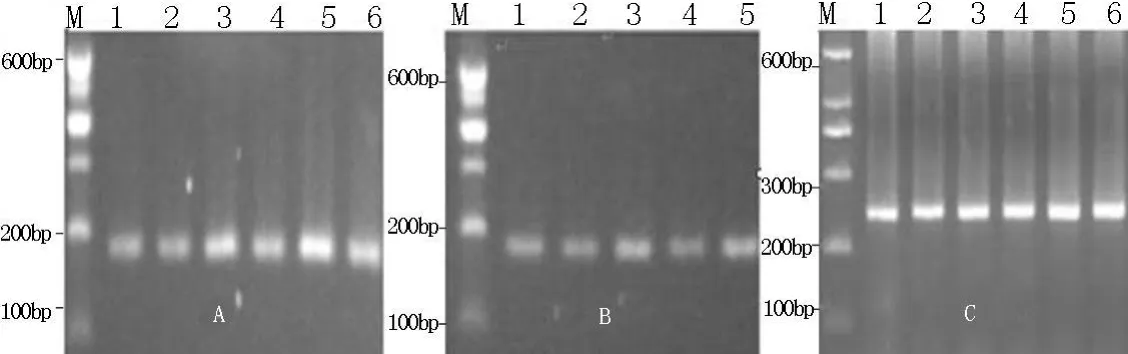
圖1 GnRHR基因引物擴增產物
2.2 SSCP檢測 對P1、P2、P3引物擴增的黃淮山羊和波爾山羊2個品種的GnRHR基因PCR產物分別進行SSCP分析。結果發現:在檢測山羊中,引物P1、P3擴增片段均不存在多態性(結果未呈示)。引物P2擴增片段有3種基因型(圖2),分別命名為AA、AB和BB,存在多態性。
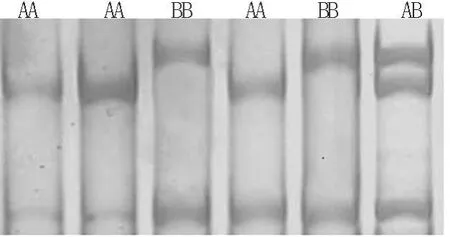
圖2 山羊GnRHR基因P2擴增片段的SSCP結果
2.3 克隆測序 經PCR-SSCP分析,所設計的引物中僅在引物P2擴增的片段中檢測到多態性,擴增出3種基因型,選擇黃淮山羊純合的AA型和BB型進行測序分析。結果顯示,擴增片段長度為188 bp;AA型與BB型序列相比較,在58 bp處發生A→C堿基突變(圖3),編碼的氨基酸由賴氨酸變為谷氨酰胺。
2.4 基因群體遺傳學分析
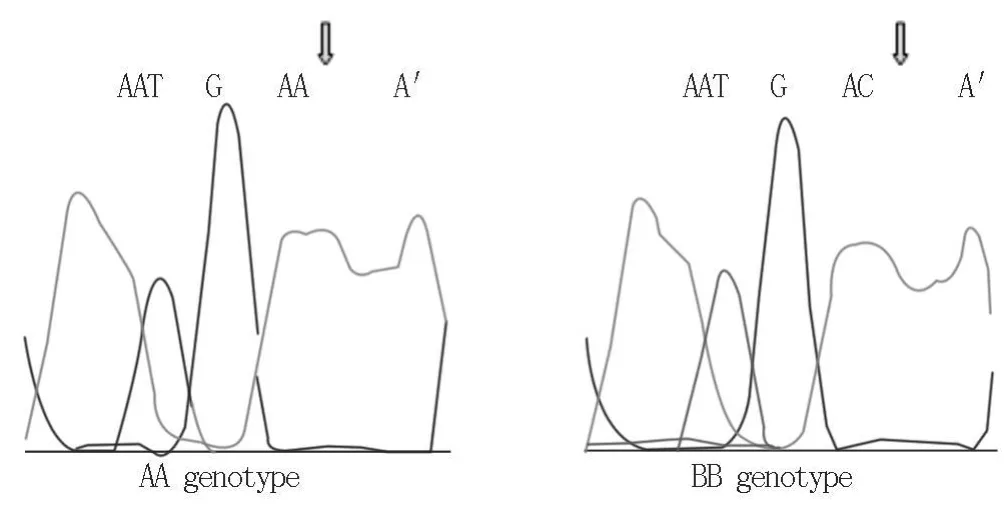
圖3 P2引物擴增片段序列及測序圖比較
2.4.1 基因頻率和基因型頻率 對引物P2擴增的黃淮、波爾山羊GnR HR基因外顯子1的基因頻率和基因型頻率進行了統計分析,其基因多態性分布情況見表3。
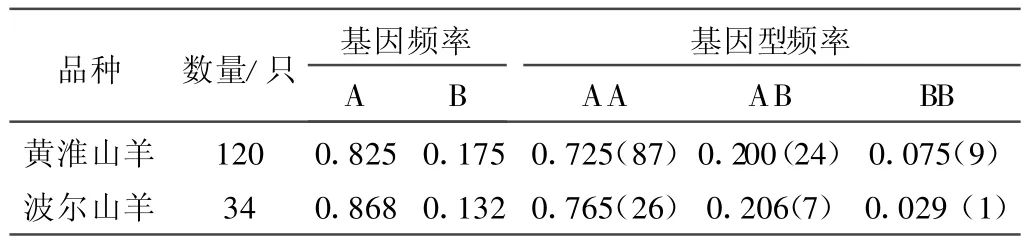
表3 GnRHR基因在黃淮山羊和波爾山羊品種中的多態性分布
2.4.2 GnRHR基因與黃淮山羊產羔數的關聯分析 黃淮山羊群體中GnRHR基因P2擴增產物不同基因型的產羔數的最小二乘均值及標準誤見表4。由表可見:BB型黃淮山羊平均產羔數比AA型多0.66只(P<0.05);AB基因型黃淮山羊產羔數比BB基因型的少0.39只,但差異不顯著(P>0.05),比AA基因型的多0.27只,差異也不顯著(P>0.05),見表4。
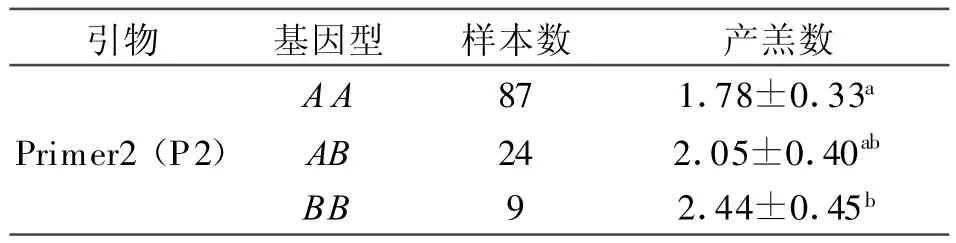
表4 不同GnRHR基因型黃淮山羊產羔數的最小二乘均值及標準誤差
3 討論
GnRHR基因是分子標記的研究熱點之一,國內外學者對GnRHR基因的多態性進行了研究。De Roux等[7]發現,人GnRHR cDNA存在317G→A和785A→G兩個突變,分別導致氨基酸改變(Gln106Arg和Arg262Gln)。吳旭等[8]在對文昌雞群體GnRHR內含子1的多態性檢測中發現,內含子的多態性對文昌雞300、400日齡產蛋數有顯著的影響,以顯性方式作用。Jiang等[9]在梅山×歐洲大白豬的F2代群體中發現GnRHR基因3'端UTR(非翻譯區)1 721位有C/G替代;1 721位等位基因G在梅山豬中的頻率為0.94,在歐洲大白豬中的頻率為0.42。
本研究在GnRHR基因外顯子1和外顯子2處共設計3對引物,在外顯子1設計的2對引物中有1對引物(P2)擴增片段具有多態,而P1的擴增片段沒有多態。克隆獲得的山羊GnRHR基因外顯子1的片段長度為232bp。GnRHR基因純合的BB基因型黃淮山羊平均產羔數比AA基因型多0.66只,差異顯著(P<0.05),而比雜合AB基因型多0.39只,差異不顯著(P>0.05)。BB型與AA型相比,在外顯子1有1個突變(C→A),而且引起氨基酸改變。由此可以推測GnRHR基因可能是控制黃淮山羊多胎性能的一個主效基因或是與之存在緊密連鎖的一個遺傳標記。
[1]劉忠慧,儲明顯,陳國宏.促性腺激素釋放激素基因與其受體基因的研究進展[J].中國畜牧獸醫,2006,33(3):35-38.
[2]姜德相,董桂紅,王達榮.GnRH對提高母豬受胎率和產仔存活數作用的研究[J].中國畜牧獸醫,2005,32(6):28-29.
[3]Dunn I C,Miao Y W,Mo rris A,et al.A study of association between genetic markers in candidate genes and reproductive traits in one generation of a commercial broiler breeder hen population[J].Heredity,2004,92(2):128-134.
[4]劉忠慧,儲明星.綿羊GnRHR基因部分序列PCR-SSCP分析[J].安徽農業大學學報,2006,33(3):318-321.
[5]儲明星,肖杰文,狄冉,等.促性腺激素釋放激素受體(GnRHR)基因多態性及其與濟寧青山羊高繁殖力關系[J].農業生物技術學報,2009,17(2):218-223.
[6]趙本領,包玉亭,霍福新,等.黃淮山羊品種資源調查報告[J].中國畜禽業,2008(1):642-661.
[7]De Roux N,Young J,Misrahi M,et al.A family with hy pogonadotropic hypogonadism and mutations in the gonadotropin-releasing ho rmone receptor[J].T he New England Journal of Medicine,1997,337(22):1597-1602.
[8]吳旭,王金玉,嚴美嬌,等.GnRHR,IGF-1基因對文昌雞繁殖性能的遺傳效應分析[J].畜牧獸醫學報,2007,38(1):31-35.
[9]Jiang ZH,Gibson J P,A rchibald A L,et al.The porcine gonadotropin-releasing hormone receptor gene(GnRHR):genomic organization,polymorphisms,and association with the number of corpora lutea[J].Genome,2001,44(1):7-12.
