KLF4通過上調Mfn2表達減輕棕櫚酸致L6骨骼肌細胞的胰島素抵抗
2014-09-13 06:25:32趙志英
中國老年學雜志 2014年21期
關鍵詞:胰島素
張 哲 趙志英 王 超
(河北省人民醫院 河北省老年醫學重點實驗室,河北 石家莊 050051)
長期胰島素抵抗與糖尿病、肥胖及高血壓等心血管疾病的發病密切相關,而骨骼肌是胰島素刺激下機體攝取并利用葡萄糖的主要靶器官〔1〕。線粒體融合蛋白(Mfn)2是表達在線粒體外膜上的一個具有多重生物學活性的細胞內分子,具有促進線粒體融合并維持線粒體形態和功能的作用。近年來研究發現,在多種胰島素抵抗的動物及細胞模型中均存在Mfn2表達減低,充分證實了線粒體損傷和功能異常是胰島素抵抗發生的重要病理機制〔2,3〕。Krüppel樣因子(KLF)4是一種鋅指蛋白轉錄因子,在細胞增殖、分化、血管形成、腫瘤發生發展中發揮廣泛的調控作用〔4〕。有研究發現KLF4可直接與Mfn2基因的啟動子序列結合促進Mfn2的表達,二者共同參與血管平滑肌細胞的分化〔5〕。然而,KLF4在骨骼肌胰島素抵抗中的作用目前尚未見報道。本實驗旨在觀察KLF4/Mfn2對胰島素抵抗的影響。
1 材料與方法
1.1材料 L6骨骼肌細胞株購自中國協和醫科大學細胞中心。α-MEM、胎牛血清(FBS)、牛血清白蛋白(BSA)、胰蛋白酶購自美國GIBCO公司。青霉素、鏈霉素、棕櫚酸、葡萄糖氧化酶試劑盒購自上海生工生物工程技術服務有限公司。兔抗KLF4、Mfn2、胰島素受體(IR)、GLUT4、GAPDH多克隆抗體購自美國Sigma公司。轉染試劑Lipofectamin 2000購自Invitrogen 公司。辣根過氧化物酶標記的羊抗鼠IgG抗體、BCA蛋白濃度測定試劑盒、高靈敏度化學發光檢測試劑盒購自北京康為世紀生物科技有限公司。Trizol、RT-PCR試劑盒購自Promega 公司。PCR引物及靶向Mfn2 mRNA的siRNA由上海生工生物工程技術服務有限公司合成。
1.2實驗方法
1.2.1細胞培養 在5% CO2、37℃飽和濕度條件下,將復蘇后的大鼠L6骨骼肌細胞接種在含10% FBS、100 U/ml青霉素、100 μg/ml鏈霉素的α-MEM培養基中培養。2 d后更換含2% FBS的α-MEM培養基進行誘導培養,每天換液一次,直至細胞長出肌管。
1.2.2胰島素抵抗模型的建立及鑒定
1.2.2.1細胞培養 采用棕櫚酸體外誘導培養法建立L6骨骼肌細胞的胰島素抵抗模型。將處于對數生長期的細胞消化后轉入24孔板中,調整細胞密度為1×106個/孔。待細胞形成成熟肌細胞后,隨機分為對照組和誘導組。對照組每孔加入不含棕櫚酸的α-MEM培養基,誘導組每孔加入含0.3 mmol/L棕櫚酸的α-MEM培養基,培養24 h待用。
1.2.2.2葡萄糖消耗實驗 將對照組和誘導組細胞用預冷0.01 mmol/L磷酸鹽緩沖液(PBS)洗滌2次,先加入含100 ng/ml胰島素的正常培養基孵育30 min,再加入5.6 mmol/L葡萄糖孵育24 h。采用葡萄糖氧化酶法測定細胞上清液中葡萄糖濃度,操作嚴格按照試劑盒說明書進行。
1.2.3重組腺病毒轉染
1.2.3.1重組腺病毒的構建 采用基因重組技術,將大鼠全長 KLF4 cDNA序列及靶向Mfn2 mRNA的siRNA克隆到腺病毒表達載體pAd/CMV/V5-DEST中。酶切和測序鑒定后,將空載體病毒及重組腺病毒利用Lipofectamin 2000脂質體轉染A293細胞,包裝和擴增后得到重組腺病毒質粒Ad-KLF4和Ad-Mfn2-siRNA。轉染24 h后,反復凍融裂解細胞并收集上清液,用0.3 g/L G418篩選陽性克隆并培養14 d。利用Real-time PCR和Western印跡檢測KLF4和Mfn2表達情況。
1.2.3.2細胞分組 將誘導成功的L6骨骼肌細胞隨機分為空載體對照組(Ad)和轉染組(Ad-KLF4和/或Ad-Mfn2-siRNA),繼續培養48 h進行后續試驗。
1.2.4Real-time PCR 根據GenBank中大鼠KLF4、Mfn2、IR、GLUT4 mRNA序列,采用DNAMAN軟件設計引物,引物序列見表1。Trizol提取各組細胞總RNA,用紫外分光光度計鑒定RNA的純度并定量。參考北京全式金生物技術有限公司 EasyScript First-strand cDNA synthesis SuperMix 試劑盒說明書進行反轉錄合成cDNA第1鏈。PCR 反應按照Syber Green I GoTaq?qPCR Master Mix試劑盒說明書進行操作。擴增完畢,進入ABI 7300型熒光定量PCR儀附帶的SDS v1.3軟件結果分析界面,設基線為3~15個循環,得到各樣本、各基因擴增的Ct值。設對照組樣品為標準1,目的基因表達水平的相對定量值RQ=2-△△Ct,將RQ值用于統計分析,設GAPDH為內參照基因。

表1 KLF4、Mfn2、IR、GLUT4、GAPDH引物序列及擴增產物長度
1.2.5Western印跡 各組細胞培養48 h后提取細胞總蛋白,BCA法測定蛋白濃度。蛋白樣品按4∶1體積比加入5×上樣緩沖液內混勻,100℃煮沸5 min變性,置于-20℃冰箱備用。將100 μg總蛋白進行10% SDS-PAGE,電轉移至PVDF膜上,10% 脫脂奶粉封閉2 h。用特異性一抗(1∶2 000稀釋) 4 ℃孵育過夜,TBST洗膜后加入辣根過氧化物酶標記的羊抗鼠IgG(1∶5 000稀釋)室溫孵育1 h,洗膜后ECL化學發光法顯色、定影。用UVP軟件分析,檢測蛋白條帶IOD值。
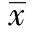
2 結 果
2.1兩組葡萄糖代謝水平的比較 誘導組細胞上清液中葡萄糖濃度〔(13.09±2.52)mmol/L〕顯著增加,與對照組〔(3.74±0.12)mmol/L〕比較差異有統計學意義(P<0.05),提示棕櫚酸誘導L6肌細胞胰島素抵抗模型建立成功。
2.2對照組和誘導組KLF4、Mfn2、IR、GLUT4表達的比較 以GAPDH為內參照,Real-time PCR和Westernw印跡結果顯示,誘導組細胞KLF4、Mfn2、IR、GLUT4 mRNA和蛋白表達水平均有顯著下降,分別相對下降了(60.52±7.43)%和(57.84±6.73)%、(54.28±3.62)%和(52.16±4.52)%、(46.63±6.21)%和(44.04±5.45)%、(44.01±6.08)%和(42.43±5.22)%,且葡萄糖代謝能力顯著降低,與對照組比較差異有統計學意義(P<0.05)。見圖1。
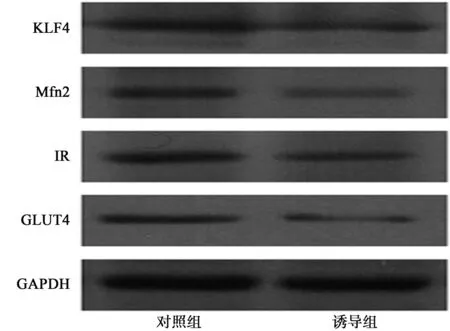
圖1 棕櫚酸刺激對KLF4、Mfn2、IR、GLU4表達的影響
2.3Ad-KLF4轉染L6骨骼肌細胞對Mfn2、IR、GLUT4 表達和葡萄糖代謝的影響 結果顯示,轉染L6骨骼肌細胞48 h后,重組腺病毒Ad-KLF4組 KLF4 mRNA和蛋白表達水平均有顯著升高,分別相對增加了(240.61±28.33)%和(222.52±20.46)%與空載體對照組比較差異有統計學意義(P<0.05),提示Ad-KLF4能夠轉染并高效表達于L6骨骼肌細胞。轉染Ad-KLF4后Mfn2、IR、GLUT4表達水平明顯上調,mRNA和蛋白表達水平分別相對增加了(183.16±35.02)%和(177.62±30.24)%、(151.22±29.34)%和(143.21±21.32)%、(159.31±32.12)%和(145.43±25.11)%,細胞對葡萄糖的代謝能力亦明顯改善。見圖2。
2.4Ad-Mfn2-siRNA轉染L6骨骼肌細胞對Mfn2、IR、GLUT4表達和葡萄糖代謝的影響 結果顯示,轉染L6肌細胞48 h后,重組腺病毒Ad-Mfn2-siRNA組Mfn2 mRNA和蛋白表達水平均有顯著降低,分別相對降低了(74.42±6.18)%和(80.35±7.32)%,與空載體對照組比較差異有統計學意義(P<0.05),提示siRNA成功干擾了L6骨骼肌細胞Mfn2的表達。另外,Ad-Mfn2-siRNA不影響Ad-KLF4的高表達,但IR、GLUT4表達水平未見升高,與空載體對照組比較差異無統計學意義(P>0.05)。細胞對葡萄糖的代謝能力亦無改善。

圖2 過表達KLF4對Mfn2、IR、GLUT4表達和葡萄糖代謝水平的影響
3 討 論
胰島素抵抗Mfn2通過調節線粒體膜電位和氧化磷酸化系統調控線粒體代謝,而Mfn2表達減低和線粒體代謝紊亂是胰島素抵抗發生的重要機制之一。本課題組前期研究證實,體外造成胰島素抵抗的肝細胞和骨骼肌細胞模型均存在Mfn2低表達,與國外報道一致〔6,7〕。
KLF4是一種具有結合位點特異性的鋅指結構轉錄因子,羧基端含有3個連續且高度保守的鋅指結構(Cχ2-4CX3FX5Lχ2HX3H),此為該家族的典型結構特征。 KLF4通過與p53、細胞周期蛋白D1等下游基因啟動子區的結合元件結合調控目的基因轉錄,在細胞增殖、分化和凋亡過程中發揮廣泛的調控作用。迄今研究發現,KLF4與腫瘤發生、皮膚屏障形成、睪丸發育等生理、病理過程密切相關〔8〕。
2010年,Rui等〔5〕采用報告基因分析發現Mfn2啟動子區存在KLF4的結合位點(CACCC/GTGGG),KLF4可激活Mfn2基因轉錄,二者在調控大鼠血管平滑肌細胞增殖的信號網絡中發揮重要作用。既然血管平滑肌細胞Mfn2可被KLF4誘導表達,我們推測在骨骼肌細胞中也有相似作用。本文結果顯示胰島素抵抗的L6骨骼肌細胞Mfn2、IR、GLUT4表達和葡萄糖代謝明顯下降,KLF4過表達可上調Mfn2、IR、GLUT4的表達并改善骨骼肌細胞的胰島素抵抗。相反,采用siRNA技術造成Mfn2基因沉默后,KLF4對上述基因表達的上調作用和骨骼肌細胞胰島素抵抗的改善作用并未出現,提示 KLF4參與了骨骼肌胰島素抵抗的發生發展,其機制可能與調控Mfn2表達有關。
KLF4在細胞內的信號調節機制尚未完全闡明。多項研究表明,KLF4和p300相互作用造成KLF4乙酰化后通過增強腸堿性磷酸酶、p21、Mfn2啟動子活性促進這些基因的轉錄和表達〔5,9〕。在骨骼肌細胞,KLF4是否通過上述機制調控Mfn2基因表達還有待進一步研究。而Mfn2促進骨骼肌細胞葡萄糖轉運的分子機制可能與GLUT4的轉位和激活有關。本實驗結果顯示,上調KLF4表達后,Mfn2和GLUT4的表達均明顯增加,二者呈協同作用,與上述觀點一致。GLUT4是肌肉和脂肪細胞膜上參與葡萄糖轉運的主要蛋白分子,其表達水平和活性與胰島素抵抗的形成有重要聯系。有研究發現,GLUT4對葡萄糖的轉運過程可被胰島素依賴的和非胰島素依賴的機制調控〔10〕。胰島素通過刺激Akt和MAPK信號途徑激活GLUT4,促進細胞對葡萄糖的攝取。而AMPK通路主要介導非胰島素依賴的糖轉運。本課題組最近研究發現,Mfn2過表達通過AMPK途徑上調GLUT4的表達,這可能是Mfn2改善骨骼肌胰島素抵抗的機制之一〔2〕。
4 參考文獻
1Lipina C,Macrae K,Suhm T,etal.Mitochondrial substrate availability and its role in lipid-induced insulin resistance and proinflammatory signalling in skeletal muscle 〔J〕.Diabetes,2013;62(10):3426-36.
2Kong D,Song G,Wang C,etal.Overexpression of mitofusin 2 improves translocation of glucose transporter 4 in skeletal muscle of high-fat diet-fed rats through AMP-activated protein kinase signaling 〔J〕.Mol Med Rep,2013;8(1):205-10.
3Gan KX,Wang C,Song GY,etal.Mitofusin-2 ameliorates high-fat diet-induced insulin resistance in liver of rats 〔J〕.World J Gastroenterol,2013;19(10):1572-81.
4Lin ZS,Chu HC,Yen YC,etal.Krüppel-like factor 4,a tumor suppressor in hepatocellular carcinoma cells reverts epithelial mesenchymal transition by suppressing slug expression 〔J〕.PLoS One,2012;7(8):e43593.
5Rui ZH,Mei HA,Bin ZH,etal.Krüppel-like factor 4 interacts with p300 to activate mitofusin 2 gene expression induced by all-transretinoic acid in VSMCs 〔J〕.Acta Pharmacol Sin,2010;31:1293-302.
6Goodpaster BH.Mitochondrial deficiency is associated with insulin resistance 〔J〕.Diabetes,2013;62(4):1032-5.
7Sebastián D,Hernández-Alvarez MI,Segalés J,etal.Mitofusin 2(Mfn2) links mitochondrial and endoplasmic reticulum function with insulin signaling and is essential for normal glucose homeostasis 〔J〕.Proc Natl Acad Sci,2012;109(14):5523-8.
8Evans PM,Liu C.Roles of Krüpel-like factor 4 in normal homeostasis,cancer and stem cells 〔J〕.Acta Biochim Biophys Sin,2008;40(7):554-64.
9Evans PM,Zhang W,Chen X,etal.Krüppel-like factor 4 is acetylated by p300 and regulates gene transcription via modulation of histone acetylation 〔J〕.J Biol Chem,2007;282:33994-4002.
10Ong KW,Hsu A,Tan BK.Chlorogenic acid stimulates glucose transport in skeletal muscle via AMPK activation:a contributor to the beneficial effects of coffee on diabetes 〔J〕.PLoS One,2012;7(3):e32718.
猜你喜歡
人人健康(2023年26期)2023-12-07 03:55:46
家庭醫藥(2019年9期)2019-09-23 18:54:32
中國生殖健康(2019年2期)2019-08-23 08:12:10
家庭科學·新健康(2018年8期)2018-10-30 10:23:20
人生與伴侶·共同關注(2018年5期)2018-08-15 10:00:00
科學生活(2016年9期)2016-10-20 13:12:45
中國衛生標準管理(2015年1期)2016-01-14 03:41:27
人人健康(2015年17期)2015-09-09 16:25:20
藥學與臨床研究(2015年4期)2015-06-05 11:35:51
中國醫藥科學(2015年15期)2015-02-27 12:32:27
