干擾對亞高山草甸和凍脹丘植物、土壤和微生物的影響
2025-02-05 00:00:00陳煒昊張國瑞許馭丹楊韋軻李佳成李帥嚴師節趙祥
草地學報 2025年1期

摘要:蘆芽山作為華北地區亞高山草甸和凍脹丘的典型分布區,在多重干擾下面臨著嚴峻的退化壓力。本研究以蘆芽山凍脹丘和草甸為研究對象,探究兩者在植物、土壤以及微生物方面的差異及其對干擾的響應。結果表明:完整凍脹丘和健康草甸在植物、土壤和微生物方面無顯著差異,受損后表現出相似的變化規律,即植物地上生物量和物種豐富度顯著降低(Plt;0.05),土壤含水率、容重和pH值發生顯著變化(Plt;0.05),細菌群落結構和多樣性發生改變,真菌僅群落結構受影響。但凍脹丘憑借土壤含水率較高的特性,能一定程度減緩干擾對植物功能群和生物量的影響。土壤理化性質對植物群落特征和土壤細菌群落特征具有顯著影響(Plt;0.05),對土壤真菌群落沒有顯著影響。綜上所述,應及時采取措施改善亞高山草甸和凍脹丘的植物群落和土壤結構,實現其生態恢復和適應性管理。
關鍵詞:亞高山草甸;凍脹丘;功能群;地上生物量;土壤理化性質;土壤微生物
中圖分類號:X171.1 " " " "文獻標識碼:A " " " "文章編號:1007-0435(2025)01-0107-12
Effects of Disturbance on Plants, Soils, and Microorganisms in Subalpine Meadows and Frost Mounds
CHEN Wei-hao1, ZHANG Guo-rui1, XU Yu-dan1*, YANG Wei-ke1, LI Jia-cheng1, LI Shuai2,
YAN Shi-jie3, ZHAO Xiang1*
(1.College of Grassland Science, Shanxi Agricultural University, Taigu, Shanxi Province 030801, China; 2.College of Resource and Environment, Shanxi Agricultural University, Taigu, Shanxi Province 030801, China; 3.Gaoping County Animal Husbandry and Veterinary Service Center, Gaoping, Shanxi Province 048400, China)
Abstract:As a typical distribution area of subalpine meadows and frost mounds in North China, Luya Mountain is being subjected to significant degradation pressure due to the combined effects of multiple disturbances. The objective of this study was to investigate the differences in plant, soil, and microbial communities and their responses to the disturbance in the frost mounds and meadows of Luya Mount. The results demonstrated that there were no significant differences between intact frost mounds and healthy meadows in terms of plants, soils and microorganisms. Furthermore, they exhibited similar patterns of change after damage, with a notable reduction in the aboveground biomass and species richness of plants (Plt;0.05), significant changes in soil water content, bulk density and pH (Plt;0.05). Additionally, bacterial community structure and diversity were altered, and only the community structure of fungi was affected. However, the higher soil water content of frost mounds can, to some extent, mitigate the effects of disturbance on plant functional groups and biomass. Soil physicochemical properties had a significant effect (Plt;0.05) on plant community characteristics and soil bacterial community characteristics. Conversely, no significant impact was observed on the soil fungal communities. In conclusion, it is imperative that prompt measures be taken to enhance the plant communities and soil structure of subalpine meadows and frost mounds, thereby facilitating their ecological restoration and adaptive management.
Key words:Subalpine meadow;Frost mound;Functional groups;Aboveground biomass;Soil physical and chemical properties;Soil microorganism
蘆芽山亞高山草甸作為華北地區典型的山地草甸景觀,在牧草生產、生物多樣性維持、水源涵養、水土保持、休閑游憩等方面發揮著重要的生產、生態和生活功能,是山西省北部重要的生態安全屏障。同時蘆芽山作為國家4A級景區,每年近百萬人次的游客量對當地生態環境構成了嚴重的威脅。此外,亞高山草甸生態系統自身有一定的“惰性”,具有能量流動和物質循環過程緩慢、對環境變化敏感、自身調節能力弱等特點,一旦遭受破壞難以恢復[1]。近年來,在牧業生產、旅游發展和自身脆弱性的多重干擾下,蘆芽山亞高山草甸發生了大面積退化。平衡當地草甸的生態保護和可持續利用,是當前亟待解決的重要課題。
凍脹丘(Frost mound)是凍土區常見的冰緣地貌[2-3],其成因是地層中水分發生聚集并凍結膨脹,使得地面抬升,導致地表呈現出錐狀、丘狀、臺狀等各種形態的隆起[4]。凍脹丘由于其獨特的形成過程,對溫度變化十分敏感[5]。在全球氣候變暖的背景下,季節性凍土區的凍結時期延遲,而解凍時期提前[6-7],這將導致凍土層厚度改變、涵養水源能力下降、植被和土壤發生退化等一系列生態問題[8]。同時,草甸退化會影響陸地生態系統的熱量收支平衡和水循環過程,導致“根系-土壤-凍土”系統嚴重失衡,進一步加速凍土和草甸的退化[9-10]。凍脹丘在蘆芽山亞高山草甸有典型分布,但在多重干擾下受損嚴重,因此明晰凍脹丘受損的生態效應成為當前凍脹丘保護和恢復的首要任務。
學者們針對凍脹丘進行了大量研究,結果表明凍脹丘的分布和特性會影響植被的分布范圍、物種組成、數量特征和生物量[11],其主要通過凍融作用改變土壤水熱條件和理化性質來影響植被的生存環境,進而對植物的生理特性和分布特征產生影響[12-13]。同時,土壤凍融過程造成的地形破損及坍塌會使得植物受到機械破壞、生長發育不良甚至死亡[11]。除了植物群落外,土壤的理化性質同樣受到凍脹丘凍融作用的影響[14-15]。凍融作用主要通過土壤冰水相態轉化過程中的體積脹縮破壞團聚體結構[5],打破顆粒間的聯結狀態而影響土壤質地[16-17],最終導致土壤容重降低、孔隙度增加、飽和導水率升高[18]。凍融過程初期,淋溶作用等促進土壤硝化過程和溶解性有機酸的釋放,使得土壤pH值降低、可利用養分含量增加[19-20]。然而頻繁的凍融循環會造成植物根部死亡和有機質的釋放,加快有機質的礦化分解速率,增加了土壤中可溶性有機物和養分的濃度,使得解凍時養分更容易伴隨水土流失而喪失,導致土壤肥力下降[21-23]。此外,不同類群土壤微生物的群落結構和多樣性對凍融作用的響應存在差異[24]。凍融過程中溫度、水分和土壤小分子有機質的變化會對土壤微生物活動造成影響[25],表現為凍融初期土壤中一些耐受性較差的微生物(主要以細菌為主)會隨著溫度降低而活性減弱甚至死亡,影響土壤微生物的物種組成和多樣性,適應環境變化后僅部分微生物可恢復至原始狀態[26-28]。然而,凍融作用對一些耐受性較強的微生物(主要以真菌為主)沒有顯著影響[29]。
綜上所述,前人對凍脹丘的研究多集中于其形成原因、分布范圍、內部結構和凍融變化,而針對凍脹丘破損之后生態效應的研究仍然較少,且缺乏關于草甸和凍脹丘對退化的響應是否一致的認識。同時,我國有關凍脹丘的研究多見于青藏高原等高寒地區,而蘆芽山作為華北地區凍脹丘的典型分布區,相關研究卻鮮見報道[4,30]。鑒于此,本研究以蘆芽山的凍脹丘及其亞高山草甸為研究對象,探究完整凍脹丘和健康草甸在植物、土壤以及微生物方面有哪些差異,以及兩者受損后的響應過程,為蘆芽山凍脹丘和亞高山草甸的生態恢復和管理提供科學依據。
1 材料與方法
1.1 研究區概況
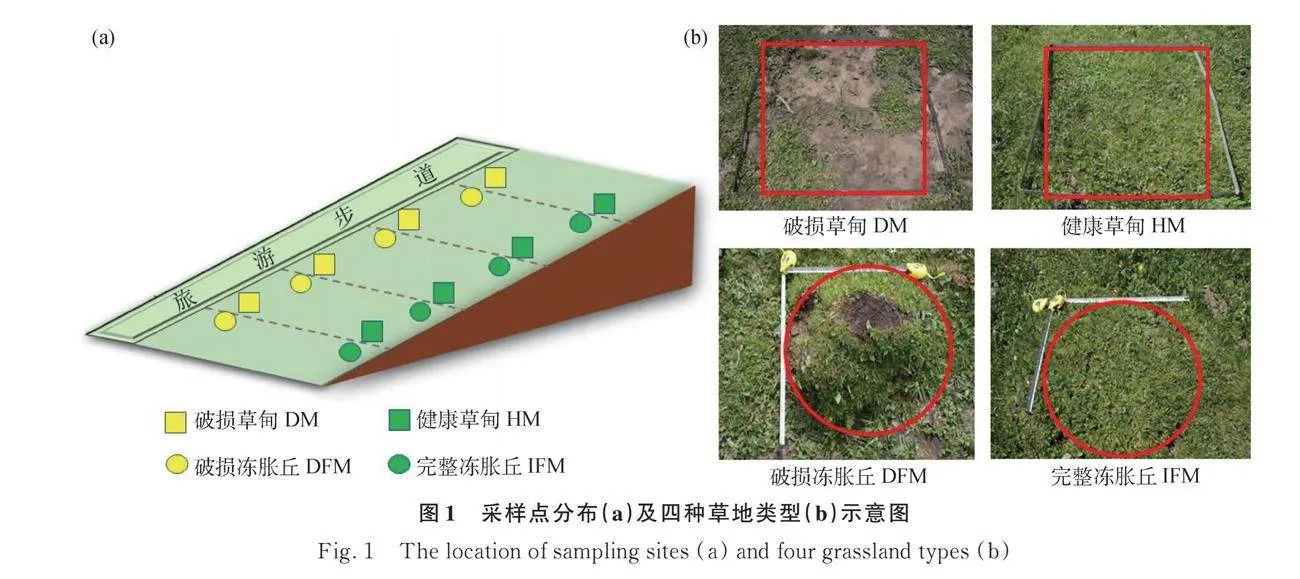
研究區位于山西省忻州市寧武縣蘆芽山馬侖草原,地理位置處于38°36′~39°02′ N,111°46′~112°54′ E,海拔2600~2700 m。研究區屬溫帶大陸性季風氣候,年均氣溫2℃~6℃,年降水量500~600 mm,年均日照2944 h。研究區主要植被類型為亞高山草甸,主要土壤類型有山地褐土和山地草甸土。凍脹丘在該地區有典型分布,其隆起呈橢圓形,直徑一般在30~80 cm,高度一般在15~35 cm。然而在牧業生產、旅游發展和自身脆弱性的多重干擾下,大量凍脹丘受到了不同程度的破壞,表現為植被矮小稀疏、土壤裸露、凍脹丘坍塌等。
1.2 試驗設計與樣品采集
2022年7月在蘆芽山馬侖草原垂直于步道布設4條平行樣帶,每條樣帶分別設置4個處理:破損草甸樣地、健康草甸樣地、破損凍脹丘樣地和完整凍脹丘樣地。每塊樣地隨機設置4個重復樣方,共設置64個樣方(圖1a)。為了保證結果的可比性,本次調查中所有凍脹丘的面積和受損程度接近(圖1b)。
亞高山草甸的植物群落樣方調查面積為1 m×1 m,凍脹丘的植物群落樣方調查面積為0.4 m×0.4 m。植物樣方調查的內容包括記錄樣方內植物物種組成、每個物種的株數、高度和蓋度。完成群落調查后,用剪刀齊地面刈割(草甸為30 cm×30 cm、凍脹丘為20 cm×20 cm)植物地上生物量后帶回實驗室烘干稱重。在每個樣方內用環刀法取0~15 cm的土壤樣品,用于測定土壤容重和含水率。用直徑為3.5 cm的土鉆采集2鉆0~15 cm的土壤樣品,一半土壤樣品風干研磨過篩后用于測定土壤理化性質;另一半土壤冷藏保存,用于土壤細菌和真菌群落的測定。
1.3 土壤理化性質分析方法
土壤容重和含水率采用烘干法測定,土壤pH值采用電極法測定。土壤有機碳采用重鉻酸鉀容量法—外加熱法進行測定,土壤全氮含量用元素分析儀(vario MACRO cube,Elementar,Germany)測定,土壤堿解氮采用堿解擴散法測定。土壤全磷和速效磷的測定分別用硫酸-高氯酸消煮和碳酸氫鈉浸提后,采用鈉鉬銻抗比色法測定。具體分析方法詳見《土壤農業化學分析方法》[31]。
1.4 土壤微生物的測定
采用OMEGA Soil DNA Kit(D5635-02)(Omega Bio-Tek,Norcross,GA,USA)試劑盒抽提樣品DNA,并利用Nanodrop對DNA進行定量。利用16S擴增子技術對細菌V3-V4區引物338F和806R進行擴增,用ITS測序技術對真菌ITS1區引物ITS5和ITS2進行擴增。PCR產物用Vazyme VAHTSTM DNA Clean Beads(Vazyme,Nanjing,China)進行純化,經瓊脂糖凝膠電泳檢測后進行測序,用Quant-iT PicoGreen dsDNA Assay Kit(Invitrogen,Carlsbad,CA,USA)對PCR產物進行定量、混樣并利用Illumina公司的TruSeq Nano DNA LT Library Prep Kit構建文庫。經過對文庫的質檢和定量后在Illumina NovaSeq儀器上利用NovaSeq 6000 SP Reagent Kit(500 cycles)進行高通量測序。最后使用QIIME2 2019.4對微生物組生物學信息進行分析并生成特征性序列ASVs以及豐度數據表格,再采用UNITE數據庫對ASV進行分類并鑒定。
1.5 數據分析與處理
將所有植物按照功能群劃分為禾本科、莎草科和雜類草三類[32]。使用相對蓋度和相對高度計算各功能群的重要值[33]。
基于SPSS26.0軟件,運用單因素方差分析(One-way ANOVAs)探究不同草地類型植物地上生物量、物種多樣性和土壤理化性質的差異;通過Venn分析來比較不同樣地細菌和真菌的物種組成差異;通過Circos圖分析不同樣地細菌和真菌的群落組成差異;運用QIIME2軟件計算Chao1指數、Shannon-Wiener指數、Simpson指數和Pielou均勻度指數,并運用單因素方差分析比較不同草地類型間微生物多樣性的差異;運用冗余分析(RDA)探究土壤理化性質對植物群落和土壤微生物群落的影響。基于Origin 2021軟件完成繪圖。
2 結果與分析
2.1 不同草地類型植被地上生物量和多樣性
總體來看,不同草地類型的各植物功能群表現出相似的變化規律,重要值從大到小依次為雜類草、莎草科和禾本科。健康草甸受損后禾本科重要值顯著升高(Plt;0.05),莎草科重要值顯著降低(Plt;0.05),雜類草無顯著變化。完整凍脹丘受損后各功能群無顯著變化。健康草甸和完整凍脹丘之間的重要值無顯著差異。破損草甸的禾本科重要值顯著高于破損凍脹丘(Plt;0.05),莎草科重要值顯著低于破損凍脹丘(Plt;0.05),二者雜類草的重要值無顯著差異(圖2a)。
健康草甸和完整凍脹丘受損后地上生物量均顯著降低(Plt;0.05),健康草甸的地上生物量顯著低于完整凍脹丘(Plt;0.05),破損草甸地上生物量也顯著低于破損凍脹丘(Plt;0.05,圖2b)。
健康草甸受損后Patrick豐富度指數顯著下降(Plt;0.05),Pielou均勻度指數顯著升高(Plt;0.05)。完整凍脹丘受損后Patrick豐富度指數顯著下降(Plt;0.05),而Pielou均勻度指數則無顯著差異(圖2c,2d)。
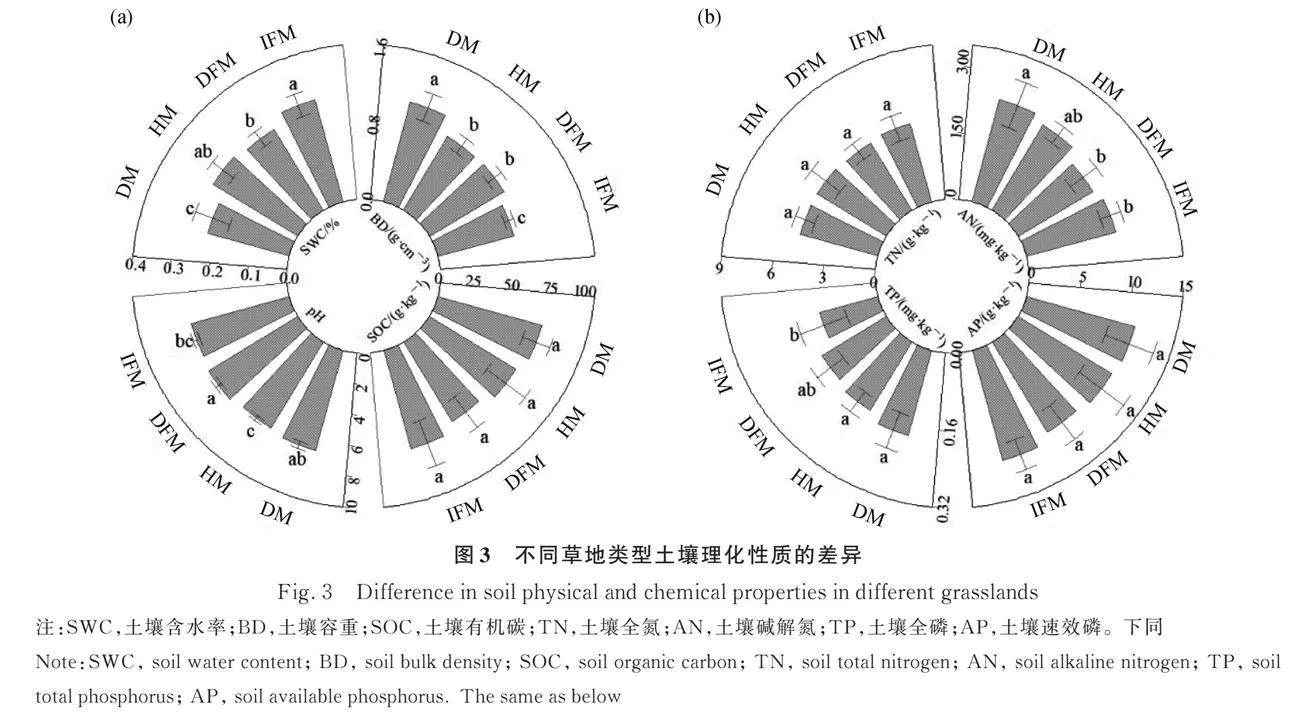
2.2 不同草地類型土壤理化性質
健康草甸和完整凍脹丘受到破壞后均表現為:土壤含水率顯著下降(Plt;0.05),土壤容重和土壤pH值顯著上升(Plt;0.05),而土壤有機碳含量、全氮含量、堿解氮含量、全磷含量和速效磷含量均無顯著差異。健康草甸的土壤容重和全磷含量顯著高于完整凍脹丘(Plt;0.05),其他指標則無顯著差異。破損草甸土壤含水率顯著低于破損凍脹丘(Plt;0.05),土壤容重和堿解氮含量顯著高于破損凍脹丘(Plt;0.05),土壤pH值、有機碳含量、全氮含量、全磷含量和速效磷含量則無顯著差異(圖3a,3b)。
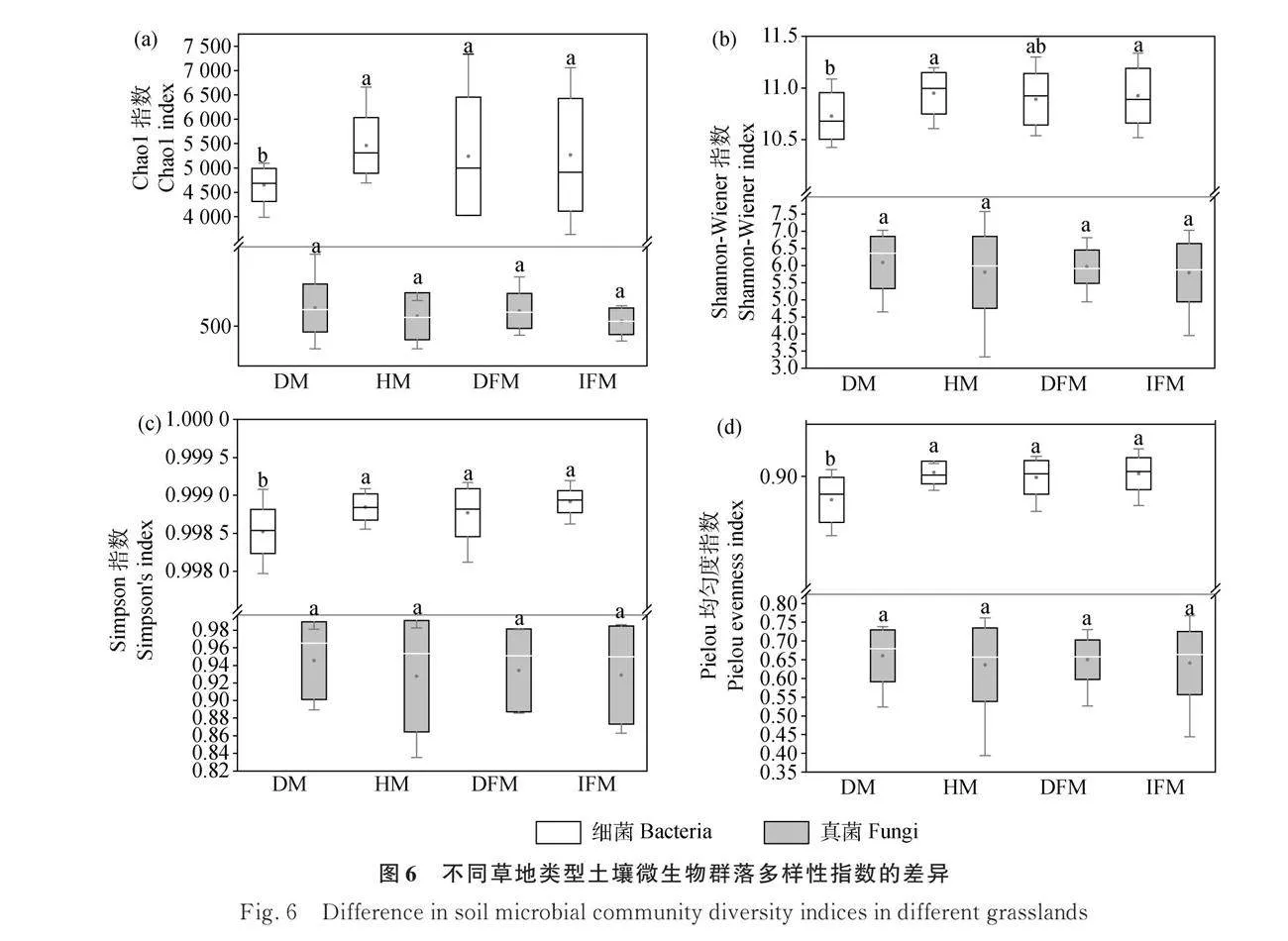
2.3 不同草地類型土壤微生物群落組成和多樣性
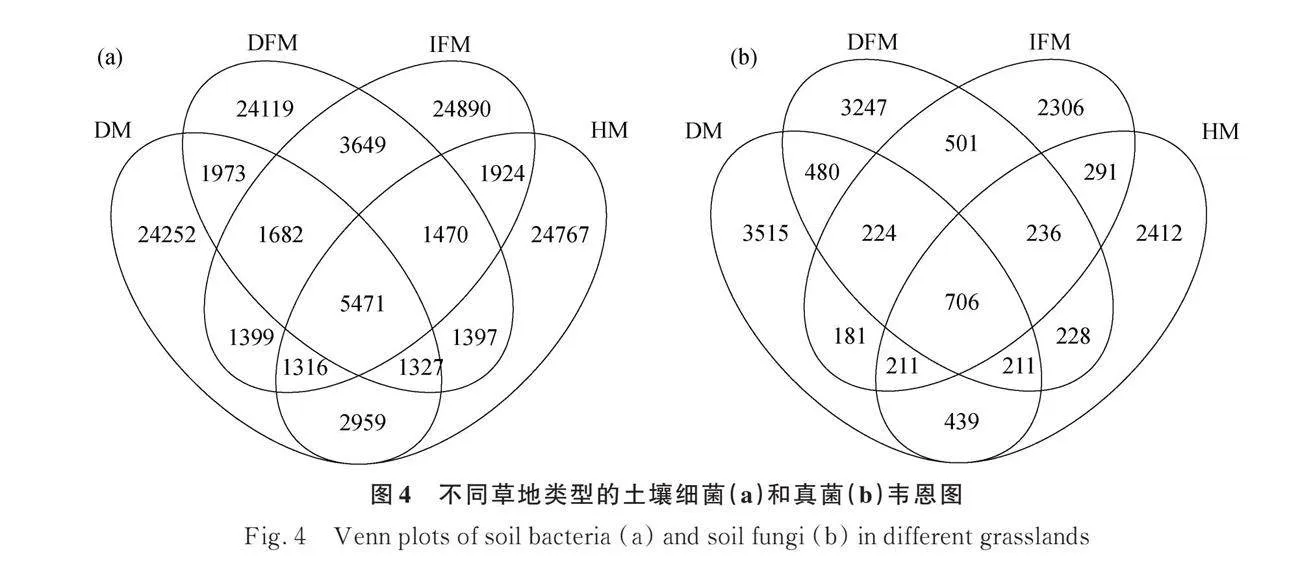
Venn分析結果表明:細菌的ASV數量遠高于真菌的ASV數量。破損草甸、健康草甸、破損凍脹丘、完整凍脹丘的細菌ASV分別為40379,40631,41088,41801。破損草甸、健康草甸、破損凍脹丘、完整凍脹丘的真菌ASV分別為5967,4734,5833,4656。破損草甸、健康草甸、破損凍脹丘、完整凍脹丘特有的細菌分別為24 252,24 767,24 119,24 890個;特有的真菌分別為3515,2412,3247,2306個(圖4a,4b)。
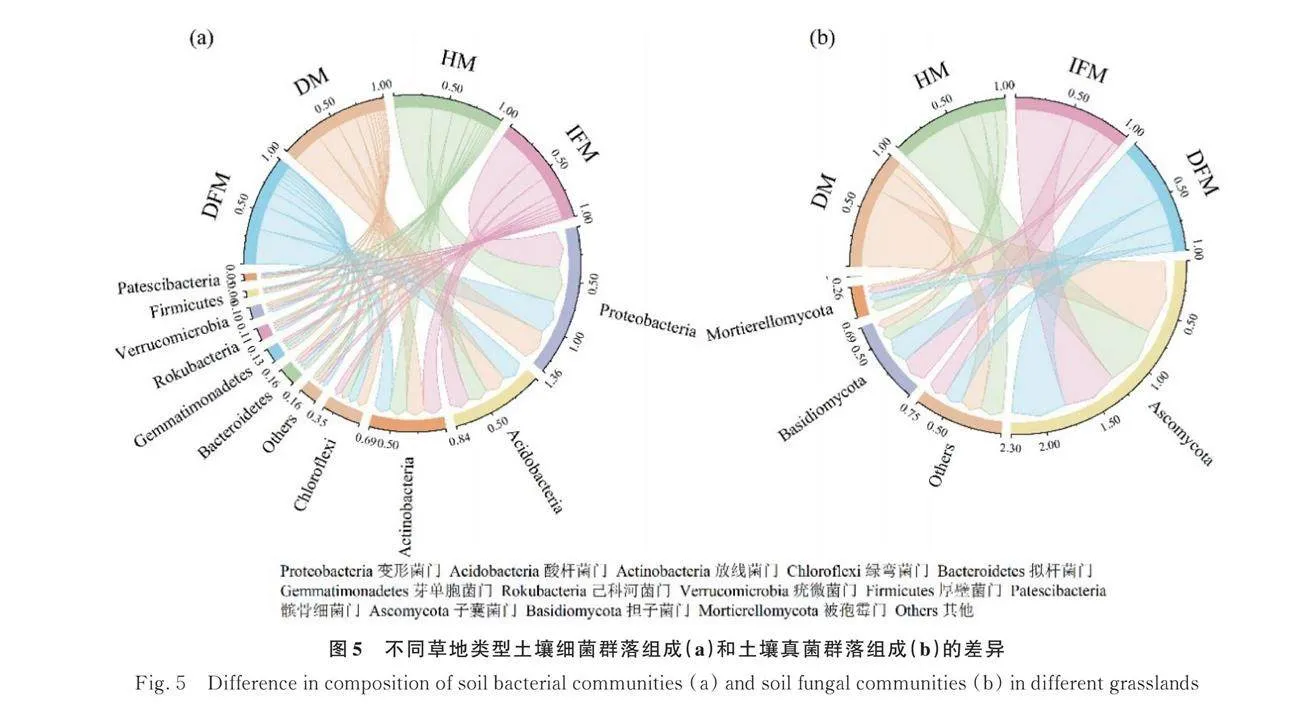
各不同草地類型土壤細菌在門水平上相對豐度大于1%的共有10個類群,其中相對豐度大于5%的優勢類群分別為變形菌門(Proteobacteria)、酸桿菌門(Acidobacteria)、放線菌門(Actinobacteria)和綠彎菌門(Chloroflexi)。當健康草地受損后優勢類群中的變形菌門相對豐度降低,酸桿菌門、放線菌門和綠彎菌門相對豐度升高。完整凍脹丘受到破壞后變形菌門和放線菌門相對豐度降低,酸桿菌門和綠彎菌門相對豐度升高。健康草甸中酸桿菌門和綠彎菌門相對豐度高于完整凍脹丘,變形菌門和放線菌門相對豐度則低于完整凍脹丘。破損草甸中放線菌門和綠彎菌門相對豐度高于破損凍脹丘,變形菌門和酸桿菌門相對豐度則低于破損凍脹丘(圖5a)。
各不同草地類型土壤真菌在門水平上相對豐度大于1%的共有3個類群,其中相對豐度大于5%的優勢類群分別為子囊菌門(Ascomycota)、擔子菌門(Basidiomycota)和被孢霉門(Mortierellomycota)。當健康草甸破損后子囊菌門相對豐度升高,擔子菌門和被孢霉門相對豐度降低。完整凍脹丘受損后被孢霉門相對豐度升高,子囊菌門和擔子菌門相對豐度降低。健康草甸中子囊菌門和被孢霉門相對豐度高于完整凍脹丘,擔子菌門相對豐度則低于完整凍脹丘。破損草甸中子囊菌門相對豐度高于破損凍脹丘,擔子菌門和被孢霉門相對豐度則低于破損凍脹丘(圖5b)。
健康草甸受到破壞后土壤中細菌的Chao1指數、Shannon-Wiener指數、Simpson指數和Pielou均勻度指數均顯著降低(Plt;0.05),真菌的Chao1指數、Shannon-Wiener指數、Simpson指數和Pielou均勻度指數無明顯變化。完整凍脹丘受損后土壤中細菌和真菌的多樣性指數均無顯著變化。健康草甸的細菌多樣性指數和真菌多樣性指數均與完整凍脹丘無顯著差異。破損草甸中細菌的Chao1指數、Simpson指數和Pielou均勻度指數均顯著低于破損凍脹丘(Plt;0.05),真菌的多樣性指數與破損凍脹丘無明顯差異(圖6)。
2.4 土壤理化性質對植物和微生物群落特征的影響
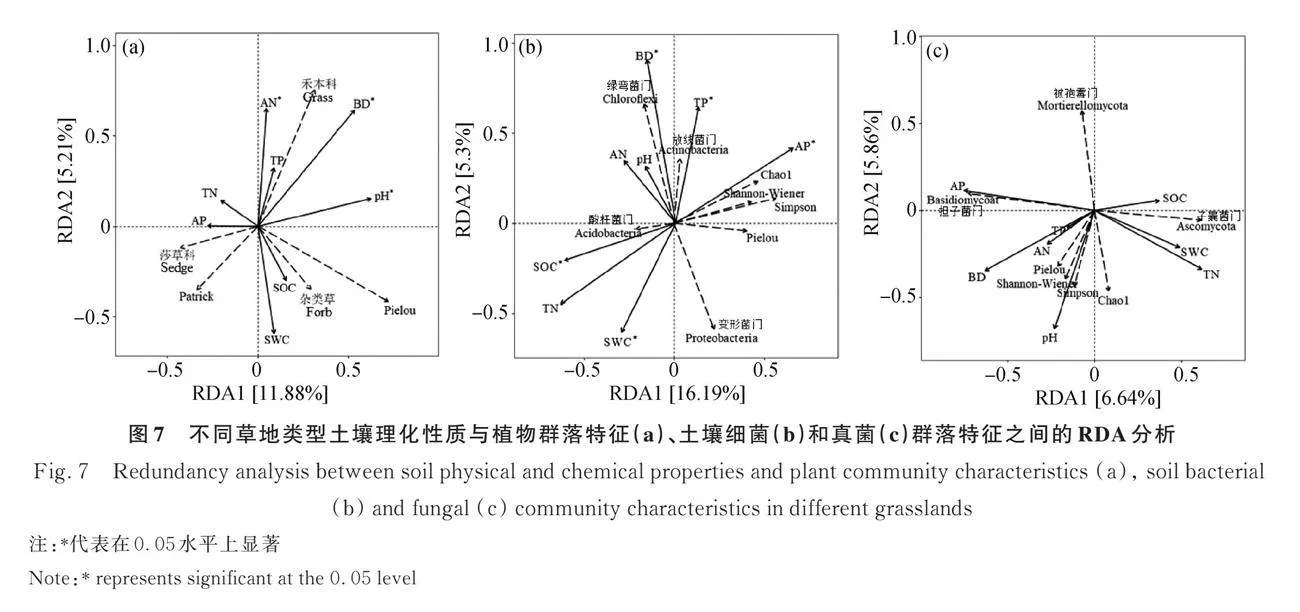
土壤理化性質對植物群落具有顯著影響(Plt;0.05)。禾本科與土壤容重、pH值、全氮、堿解氮和全磷正相關,莎草科與土壤含水率、全氮和速效磷正相關,雜類草與土壤含水率、pH值和有機碳正相關,Patrick豐富度指數與土壤含水率、有機碳、全氮和速效磷正相關,Pielou均勻度指數與土壤含水率、容重、pH值和有機碳正相關。土壤容重、pH值和堿解氮是顯著影響植物群落的主要因子(Plt;0.05,圖7a)。
土壤理化性質對土壤細菌群落具有極顯著的影響(Plt;0.01)。變形菌門與土壤含水率和全氮正相關,酸桿菌門與土壤含水、容重、pH值、有機碳、全氮和堿解氮正相關,放線菌門與土壤容重、pH值、堿解氮、全磷和速效磷正相關,綠彎菌門與土壤容重、pH值、堿解氮、全磷和速效磷正向關,Chao1指數、Shannon-Wiener指數和Simpson指數均與土壤容重、全磷和速效磷正相關,Pielou均勻度指數與土壤全磷和速效磷正相關。土壤含水率、容重、有機碳、全氮、全磷和速效磷是顯著影響細菌群落特征的主要因子(Plt;0.05,圖7b)。
土壤理化性質對土壤真菌群落沒有顯著影響。子囊菌門與土壤含水率、有機碳和全氮正相關,擔子菌門與土壤容重、pH值、堿解氮、全磷和速效磷正相關,被孢霉門與速效磷呈正相關,Chao1指數與土壤含水率、容重、pH值、全氮、堿解氮和全磷正相關,Shannon-Wiener指數、Simpson指數和Pielou均勻度指數均與土壤含水率、容重、pH值、全氮、堿解氮、全磷和速效磷正相關(圖7c)。
3 討論
3.1 亞高山草甸退化對植物地上生物量和多樣性的影響
植物群落對草地退化的響應最為迅速直觀,其物種組成、生物量及物種多樣性等是判斷草地生態系統退化的重要指標[34-35]。研究表明,草甸發生退化后,禾本科和莎草科植物被雜類草代替,植物群落結構趨于單一化,多樣性指數降低[36-38],這與本研究的結果有所差異。本研究發現草甸受損后禾本科重要值顯著升高,莎草科重要值顯著降低,這與Xu等[39]的研究結果一致。受損后禾本科群落地位上升的原因可能是原本較為穩定的草地生態系統在退化過程中土壤含水率降低,莎草科植物為了吸收更多水分加強地下根系生長,地上部分受到削弱,導致地上生物量下降,競爭能力變弱;而禾本科植物多為旱生植物和半旱生植物,對水分的需求較少,土壤低含水量時適應性較強,使得禾本科重要值相對升高[40-42]。
其次,本研究發現破損導致健康草甸和完整凍脹丘的地上生物量顯著下降,這與前人的研究結果一致[43-45]。破損導致植被地上生物量下降的原因主要有兩方面:一方面是過度放牧和旅游踐踏等人為活動會直接破壞草地植被,使其受到機械損傷,造成光合作用減弱,生長受限,最終導致生物量減少[1,46];另一方面是頻繁踐踏導致土壤孔隙度減小、緊實度增加,限制了根部對水分和營養物質的吸收和運輸,從而降低植物生產力[47]。同時,本研究發現無論是健康草甸還是破損草甸,其地上生物量均顯著低于凍脹丘。其原因可能是凍脹丘上植物種類的群落結構與草甸不同,這些植物可能具有更高的生物量,同時凍脹丘內部充足的水分可以加快植物根部對養分的吸收和轉運,促進植物的生長發育和生物量的積累,使得凍脹丘地上生物量顯著高于健康草甸和破損草甸[48]。
此外,本研究發現破損導致健康草甸和完整凍脹丘的Patrick物種豐富度指數下降,這與前人的研究相一致[49]。造成物種豐富度下降的原因可能是破損導致植被的生長發育受到嚴重干擾;同時植被蓋度的下降加劇了雨水沖刷造成的養分流失,惡劣的生境脅迫導致部分植物消失[50]。同時,本研究發現健康草甸和完整凍脹丘受損之后的Patrick物種豐富度指數與地上生物量有相同變化趨勢,這可能與生態位互補效應(Niche complementarity effect)和取樣效應(Sampling effect)有關[51]。生態位互補效應認為物種多樣性的下降使得植物群落對資源的利用不夠充分,進而導致群落整體生物量下降。取樣效應認為群落中競爭力較強的物種可以更有效的利用資源,而物種多樣性的下降使得群落內包含高產物種的可能性降低[52-54]。
周華坤[55]和楊元武[56]等在青藏高原的研究發現高寒草甸的物種均勻度指數隨退化程度的增加,呈單峰式的曲線變化規律,在中度退化水平達到最高,但整體呈下降趨勢。本研究發現健康草甸破損后均勻度指數顯著升高,此時可能正處于輕度或中度退化水平。其原因可能是退化過程中原生植被地上部分遭到明顯破壞,生物量減少,可利用空間增大。同時,退化導致土壤含水量減少,這都會使雜類草等適應性較強的物種數量相對增多,使群落內物種數量分布更加均勻,最終導致均勻度指數上升[38,57-58]。然而凍脹丘因其內部含有大量水分,在一定程度上削弱了禾本科和莎草科的競爭,減緩了雜類草的入侵時間,導致其破損后功能群和Pielou均勻度指數并無明顯變化。
3.2 亞高山草甸退化對土壤理化性質的影響
土壤理化性質可以反映草地退化的進程和趨勢,揭示生態系統的受損程度[59]。本研究發現,健康草甸受損后土壤含水率顯著下降,土壤容重和土壤pH值顯著上升,這與趙帥等[45]對祁連山國家公園退化草甸的研究結果一致。受損過程中游客頻繁的踐踏行為使土壤緊實度和容重增加,植被覆蓋度減少,導致土壤蒸散發速率加快,土壤含水率降低,同時大量鹽分伴隨水分蒸發過程向地表聚集,造成土壤pH值上升[60]。然而本研究中草甸受損前后土壤有機碳、全氮、堿解氮、全磷和速效磷等養分含量均無顯著差異,這與前人研究結果不一致[61-62],這可能是因為土壤物理性質的變化相較于土壤養分變化對退化的響應更為直接快速。草地受損后植物群落最先受到影響,植被對表層土壤的保護作用受到削弱,土壤物理性質逐步惡化,然而由于土壤養分含量的變化表現出一定的滯后性,尚未出現明顯的下降[63-65]。凍脹丘受損后土壤有機碳、全氮、堿解氮、全磷和速效磷等養分含量同樣無明顯差異。因此,應采取圍欄封育、控制游客量、補播等措施促進亞高山草甸和凍脹丘的有效恢復。
前人的研究表明凍融作用會使土壤pH值降低、有機碳含量減少、并影響氮和磷的遷移轉化方向[66-68]。本研究發現除了土壤容重和土壤全磷外,健康草甸和完整凍脹丘的含水率、pH值、有機碳含量、全氮含量、堿解氮和速效磷含量沒有顯著差異,其原因可能是以莎草科為主的優勢種無性繁殖旺盛,地表根系發達,導致草氈層較厚,對土壤水分和熱量具有保護作用,從而削弱了凍融作用對土壤理化性質的影響[9,69-70]。此外,完整凍脹丘容重顯著低于健康草甸,其原因是凍融作用通過土壤冰水相態轉化過程改變了土壤孔隙結構并破壞了土壤團聚體結構,導致孔隙度增加、容重降低[16]。
3.3 亞高山草甸退化對土壤微生物的影響
土壤微生物是維持草地生態系統物質循環的關鍵組分,且對外界環境的變化十分敏感,可以作為草甸退化的重要指示指標[71]。本研究結果顯示不同樣地類型細菌的ASV數量遠高于真菌的ASV數量,細菌數量占優勢,與大多數學者研究一致[72-73]。闞海明等[74]發現北京西北淺山區退化草地土壤的主要細菌菌群為變形菌門、放線菌門、厚壁菌門、酸桿菌門和綠彎菌門;金志薇等[75]研究發現滇西北高寒草地土壤真菌優勢菌群為子囊菌門和擔子菌門,本研究的優勢菌群與之類似。由此可見,不同草地類型土壤細菌、真菌的相對豐度在退化進程中均有變化,但都具有相似的優勢類群。本研究發現健康草甸和完整凍脹丘受損后均表現為細菌優勢類群中的變形菌門相對豐度降低,綠彎菌門相對豐度升高,其原因可能為土壤環境受到破壞,削弱了變形菌門中許多物種氧化有機和無機化合物的能力,而酸桿菌門作為一種寡營養型生物則更適生存[76]。同時,本研究發現健康草甸和完整凍脹丘受損后均表現為真菌優勢類群中的擔子菌門相對豐度降低,而擔子菌門富含木質纖維素酶,能分解木質纖維素,促進營養物質和能量的流動,其豐度的降低說明植物群落受到了破壞[77-78]。
其次,本研究結果發現,草甸破損后土壤中細菌的Chao1指數、Shannon-Wiener指數、Simpson指數和Pielou均勻度指數均顯著降低,這與王英成等[79]對三江源區退化高寒草甸的研究一致。草甸受損后,細菌多樣性顯著下降,這可能是因為草甸破損導致原生植物群落受到破壞,植物豐富度下降,根系分泌物和枯落物的多樣性也隨之降低,相對應的微生物也會減少[80-81]。然而本研究中完整凍脹丘受損后細菌的Chao1指數、Shannon-Wiener指數、Simpson指數和Pielou均勻度指數均沒有顯著變化,這可能與完整凍脹丘容重顯著低于健康草甸、破損凍脹丘容重顯著低于破損草甸有關。研究表明,土壤容重會影響土壤的孔隙度和通氣性,而微生物需要氧氣進行呼吸和代謝活動,高容重會限制某些需氧微生物的生長,因此凍脹丘相對較高的土壤通透性可以減弱破損帶來的影響[82]。本研究結果顯示,破損對草甸和凍脹丘土壤真菌群落的多樣性并無顯著影響,這與楊明新等[83]對三江源區退化高寒草甸的研究一致。這可能是因為細菌在物質循環等方面具有更高的活性,對環境變化敏感,因此更容易受到草地退化的影響,而真菌更傾向于與植物根系形成共生關系,這種共生關系相對穩定,使得真菌群落在退化情況下相對不易改變[84]。
此外,本研究中健康草甸的細菌和真菌多樣性指數與完整凍脹丘無顯著差異,與前人研究表明的強烈凍融作用會降低土壤微生物群落的多樣性和豐富度的結果有所差異[85]。這可能與土壤微生物多樣性受到土壤總碳、有機碳、全氮和總磷等養分的顯著影響有關[86],本研究中健康草甸和完整凍脹丘的大部分土壤養分含量并無顯著差異,使得兩者的微生物多樣性指數無顯著差異。
3.4 亞高山草甸土壤理化性質對植物群落特征和微生物群落特征的影響
土壤理化性質與植物群落特征的RDA分析解釋率較低的原因可能是海拔和坡向等其他環境因子使水熱條件發生改變,導致環境異質性分布,進而影響植物群落特征[87]。而土壤理化性質與土壤細菌群落特征的RDA分析解釋率較低的原因可能與細菌群落特征還受到海拔和植物群落組成等其他因素的影響[88]。
本研究中植物群落特征和細菌群落特征均受到土壤容重的顯著影響,與前人研究一致[89-90]。其原因是土壤容重直接決定了土壤孔隙度的大小,而土壤孔隙度一方面影響植物根部的呼吸作用,從而干擾植物物質的吸收和轉運,最終影響植物群落[47],另一方面土壤孔隙度會改變土壤通氣狀況,從而干擾土壤微生物的呼吸和代謝活動,最終影響微生物群落[91]。
4 結論
本研究發現完整凍脹丘和健康草甸在植物、土壤以及微生物方面沒有明顯差異,其受損后表現出相似的變化規律。對于植物而言,破損導致功能群發生改變、植物群落物種豐富度和生物量均顯著下降;對于土壤理化性質而言,破損導致土壤含水率、容重和pH發生顯著變化,而土壤有機碳、全氮、堿解氮、全磷和速效磷含量均無顯著變化;對于土壤微生物而言,破損導致細菌的群落結構和多樣性發生改變,真菌只有群落結構受到影響。但凍脹丘憑借自身含水率較高的特性,能有效緩解干擾對植物功能群和生物量的影響。冗余分析表明,土壤理化性質對植物群落特征和土壤細菌群落特征具有顯著影響,對土壤真菌群落沒有顯著影響,其中土壤容重對植物群落和細菌群落均有顯著影響。綜上所述,干擾背景下蘆芽山亞高山草甸和凍脹丘的植物群落、土壤物理性質和pH受到較大影響,土壤養分并未顯著下降。為防止其受到進一步破壞,可以對受損草甸和凍脹丘采取圍欄封育、草種補播、控制游客量、加強宣傳教育等措施,以達到改良植被、固持土壤、減緩破壞、逐步恢復的目的。
參考文獻
[1] 章異平,江源,劉全儒,等. 放牧壓力下五臺山高山、亞高山草甸的退化特征[J]. 資源科學,2008,30(10):1555-1563
[2] GROSSE G,JONES B M. Spatial distribution of pingos in northern Asia[J]. The Cryosphere,2011,5(1):13-33
[3] POLLARD W H,FRENCH H M. The groundwater hydraulics of seasonal frost mounds,North Fork Pass,Yukon Territory[J]. Canadian Journal of Earth Sciences,1984,21(10):1073-1081
[4] 吳吉春,盛煜,曹元兵,等. 青藏高原發現大型凍脹丘群[J]. 冰川凍土,2015,37(5):1217-1228
[5] 孫輝,秦紀洪,吳楊.土壤凍融交替生態效應研究進展[J]. 土壤,2008,40(4):505-509
[6] LI X,JIN R,PAN X D,et al. Changes in the near-surface soil freeze–thaw cycle on the Qinghai-Tibetan Plateau[J]. International Journal of Applied Earth Observation and Geoinformation,2012,17:33-42
[7] LI N,CUO L,ZHANG Y. On the freeze-thaw cycles of shallow soil and connections with environmental factors over the Tibetan Plateau[J]. Climate Dynamics,2021,57(11-12):3183-3206
[8] 姚檀棟,秦大河,沈永平,等. 青藏高原冰凍圈變化及其對區域水循環和生態條件的影響[J]. 自然雜志,2013,35(3):179-186
[9] 尚占環,董全民,施建軍,等. 青藏高原“黑土灘”退化草地及其生態恢復近10年研究進展——兼論三江源生態恢復問題[J]. 草地學報,2018,26(1):1-21
[10] 田曉暉,張立鋒,張翔,等. 三江源區退化高寒草甸蒸散特征及凍融變化對其的影響[J]. 生態學報,2020,40(16):5649-5662
[11] 周興民,楊福囤,李秉文,等. 青藏公路南段植被與多年凍土的相互關系[J]. 植物學報,1978(1):13-19,91
[12] 杜子銀,蔡延江,王小丹,等. 土壤凍融作用對植物生理生態影響研究進展[J]. 中國生態農業學報,2014,22(1):1-9
[13] 王洋,劉景雙,王全英.凍融作用對土壤團聚體及有機碳組分的影響[J]. 生態環境學報,2013,22(7):1269-1274
[14] 范繼輝,魯旭陽,王小丹. 藏北高寒草地土壤凍融循環過程及水熱分布特征[J]. 山地學報,2014,32(4):385-392
[15] 趙云朵,胡霞. 基于CT研究凍融對高寒草甸土壤孔隙結構的影響[J]. 水土保持學報,2020,34(3):362-367
[16] LEHRSCH G A,SOJKA R E,CARTER D L,et al. Freezing effects on aggregate stability affected by texture, mineralogy, and organic matter[J]. Soil Science Society of America Journal,1991,55(5):1401-1406
[17] 杜子銀,王小丹,洪江濤,等. 凍融及牲畜排泄物作用下的高寒草地土壤物理特性和氮磷變化[J]. 山地學報,2022,40(1):29-42
[18] 鄧西民,陳端生,王堅,等. 凍融作用對犁底層土壤物理性狀的影響[J]. 科學通報,1998,43(23):2538-2541
[19] 高敏,李艷霞,張雪蓮,等. 凍融過程對土壤物理化學及生物學性質的影響研究及展望[J]. 農業環境科學學報,2016,35(12):2269-2274
[20] 李貴圓,范昊明. 凍融作用對農田磷素轉化遷移影響研究進展[J]. 中國水土保持科學,2011,9(6):114-120
[21] FREPPAZ M,WILLIAMS B L,EDWARDS A C,et al. Simulating soil freeze/thaw cycles typical of winter alpine conditions:Implications for N and P availability[J]. Applied Soil Ecology,2007,35(1):247-255
[22] GROGAN P,MICHELSEN A,AMBUS P,et al. Freeze–thaw regime effects on carbon and nitrogen dynamics in sub-arctic heath tundra mesocosms[J]. Soil Biology and Biochemistry,2004,36(4):641-654
[23] 王洋,劉景雙,王國平,等. 凍融作用與土壤理化效應的關系研究[J]. 地理與地理信息科學,2007,23(2):91-96
[24] SCHOSTAG M,PRIEMé A,JACQUIOD S,et al. Bacterial and protozoan dynamics upon thawing and freezing of an active layer permafrost soil[J]. The ISME Journal,2019,13(5):1345-1359
[25] NIKRAD M P,KERKHOF L J,H?GGBLOM M M. The subzero microbiome:microbial activity in frozen and thawing soils[J]. FEMS Microbiology Ecology,2016,92(6):fiw081
[26] STRES B,PHILIPPOT L,FAGANELI J,et al. Frequent freeze-thaw cycles yield diminished yet resistant and responsive microbial communities in two temperate soils:a laboratory experiment[J]. FEMS Microbiology Ecology,2010,74(2):323-335
[27] YERGEAU E,KOWALCHUK G A. Responses of Antarctic soil microbial communities and associated functions to temperature and freeze-thaw cycle frequency[J]. Environmental Microbiology,2008,10(9):2223-2235
[28] WALKER V K,PALMER G R,VOORDOUW G. Freeze-thaw tolerance and clues to the winter survival of a soil community[J]. Applied and Environmental Microbiology,2006,72(3):1784-1792
[29] M?NNIST? M K,TIIROLA M,H?GGBLOM M M. Effect of freeze-thaw cycles on bacterial communities of Arctic Tundra Soil[J]. Microbial Ecology,2009,57(3):621-631
[30] 王平. 青藏鐵路多年凍土區凍脹丘發展特征及其對路基穩定性的影響[J]. 鐵道標準設計,2013(5):1-4
[31] 魯如坤.土壤農業化學分析方法[M].北京:中國農業科學技術出版社,2000:40-60
[32] 孫國鈞,張榮,周立. 植物功能多樣性與功能群研究進展[J]. 生態學報,2003,23(7):1430-1435
[33] 馬克平,黃建輝,于順利,等. 北京東靈山地區植物群落多樣性的研究Ⅱ豐富度、均勻度和物種多樣性指數[J]. 生態學報,1995,15(3):268-277
[34] TANG L,DONG S,SHERMAN R,et al. Changes in vegetation composition and plant diversity with rangeland degradation in the alpine region of Qinghai-Tibet Plateau[J]. The Rangeland Journal,2015,37(1):107
[35] 李雪萍,許世洋,李敏權,等. 甘南州不同退化程度高寒草甸植被及土壤特性的演化規律[J]. 生態學報,2022,42(18):7541-7552
[36] PENG F,XUE X,LI C Y,et al. Plant community of alpine steppe shows stronger association with soil properties than alpine meadow alongside degradation[J]. Science of The Total Environment,2020,733:139048
[37] 羅方林,張法偉,王春雨,等. 青藏高原高寒草甸群落特征和代表性植物生存狀態對草地退化的響應[J]. 生態學雜志,2022,41(1):18-24
[38] 李軍豪,楊國靖,王少平. 青藏高原區退化高寒草甸植被和土壤特征[J]. 應用生態學報,2020,31(6):2109-2118
[39] XU H P,ZHANG J,PANG X P,et al. Responses of plant productivity and soil nutrient concentrations to different alpine grassland degradation levels[J]. Environmental Monitoring and Assessment,2019,191(11):678
[40] POORTER H,NIKLAS K J,REICH P B,et al. Biomass allocation to leaves,stems and roots:meta‐analyses of interspecific variation and environmental control[J].New Phytologist,2012,193(1):30-50
[41] SUN D S,WESCHE K,CHEN D D,et al. Grazing depresses soil carbon storage through changing plant biomass and composition in a Tibetan alpine meadow[J]. Plant,Soil and Environment,2011,57(6):271-278
[42] WANG X X,DONG S K,YANG B,et al. The effects of grassland degradation on plant diversity, primary productivity, and soil fertility in the alpine region of Asia’s headwaters[J]. Environmental Monitoring and Assessment,2014,186(10):6903-6917
[43] ZHANG D J,QI Q,TONG S Z,et al. Soil degradation effects on plant diversity and nutrient in tussock meadow wetlands[J]. Journal of Soil Science and Plant Nutrition,2019,19(3):535-544
[44] 孫海群,林冠軍,李希來,等. 三江源地區高寒草甸不同退化草地植被群落結構及生產力分析[J]. 黑龍江畜牧獸醫,2013(19):1-3
[45] 趙帥,楊文權,藺寶珺,等. 祁連山國家公園不同退化高寒草甸植物與土壤特性研究[J]. 草地學報,2023,31(5):1530-1538
[46] 鞏劼,陸林,晉秀龍,等. 黃山風景區旅游干擾對植物群落及其土壤性質的影響[J]. 生態學報,2009,29(5):2239-2251
[47] 杜志勇,叢楠. 植被與土壤特征對青藏高原不同程度退化草地的響應 [J]. 生態學報,2024,44(6):2504-2516
[48] 顧永超,李多才,侯扶江. 高寒草原季節牧場生物量對土壤水分的響應[J]. 草業科學,2019,36(6):1490-1497
[49] 王夢夢,張麗華,當知才讓,等. 高寒濕地退化對植物群落特征與土壤特性的影響[J]. 生態學報,2023,43(19):7910-7923
[50] 劉旻霞,劉成,楊春亮. 甘南高寒草甸退化對植物功能群物種多樣性與物種多度分布的影響[J]. 西北植物學報,2024,44(1):142-153
[51] 楊元合,饒勝,胡會峰,等. 青藏高原高寒草地植物物種豐富度及其與環境因子和生物量的關系[J]. 生物多樣性,2004,12(1):200-205
[52] LOREAU M,HECTOR A. Partitioning selection and complementarity in biodiversity experiments[J]. Nature, 2001,412(6842):72-76
[53] 江小雷,張衛國,嚴林,等. 植物群落物種多樣性對生態系統生產力的影響[J]. 草業學報,2004,13(6):8-13
[54] 王增如,王成,董曉紅,等. 環境因子影響下疏勒河上游高寒草甸物種豐富度與生物量間的關系[J]. 冰川凍土, 2016,38(6):1710-1717
[55] 周華坤,趙新全,周立,等. 青藏高原高寒草甸的植被退化與土壤退化特征研究[J]. 草業學報,2005,14(3):31-40
[56] 楊元武,李希來,周旭輝,等. 高寒草甸植物群落退化與土壤環境特征的關系研究[J]. 草地學報,2016,24(6):1211-1217
[57] 李海英,彭紅春,王啟基. 高寒矮嵩草草甸不同退化演替階段植物群落地上生物量分析[J]. 草業學報,2004,13(5):26-32
[58] 王雪超,劉艷萍,高永,等. 旅游擾動對草原植被及土壤的影響[J]. 草原與草坪,2021,41(6):127-131,138
[59] 邵建翔,劉育紅,馬輝,等. 退化高寒草地淺層土壤理化性質Meta分析[J]. 草地學報,2022,30(6):1370-1378
[60] XIE H H,WU Q G,HU J Y,et al. Changes in soil physical and chemical properties during the process of alpine meadow degradation along the eastern Qinghai-Tibet Plateau[J]. Eurasian Soil Science,2018,51(12):1440-1446
[61] LI H Y,QIU Y Z,YAO T,et al. Nutrients available in the soil regulate the changes of soil microbial community alongside degradation of alpine meadows in the northeast of the Qinghai-Tibet Plateau[J]. Science of The Total Environment,2021,792:148363
[62] SHE Y D,ZHANG Z H,MA L,et al. Vegetation attributes and soil properties of alpine grassland in different degradation stages on the Qinghai-Tibet Plateau,China:a meta-analysis[J]. Arabian Journal of Geosciences,2022,15(2):193
[63] 李紹良,陳有君,關世英,等. 土壤退化與草地退化關系的研究[J]. 干旱區資源與環境,2002,16(1):92-95
[64] 魏強,王芳,陳文業,等. 黃河上游瑪曲不同退化程度高寒草地土壤物理特性研究[J]. 水土保持通報,2010,30(5):16-21
[65] 魏茂宏,林慧龍. 江河源區高寒草甸退化序列土壤粒徑分布及其分形維數[J]. 應用生態學報,2014,25(3):679-686
[66] 柴瑜,劉育紅,魏衛東. 凍融作用對退化紫花針茅草原有機碳含量及組分的影響[J]. 黑龍江畜牧獸醫,2019(13):97-102
[67] 韓露,萬忠梅,孫赫陽. 凍融作用對土壤物理、化學和生物學性質影響的研究進展[J]. 土壤通報,2018,49(3):736-742
[68] 王藝璇,仲秋維,鄭昕雨,等. 凍融循環對土壤性狀特征影響研究進展[J]. 中國土壤與肥料,2022(10):231-240
[69] 梁東營,林麗,李以康,等. 三江源退化高寒草甸草氈表層剝蝕過程及發生機理的初步研究[J]. 草地學報,2010,18(1):31-36
[70] 魏衛東,劉育紅,馬輝,等. 三江源區退化高寒草甸淺層土壤凍融作用特征[J]. 生態與農村環境學報,2019,35(3):352-359
[71] 尚占環,丁玲玲,龍瑞軍,等. 江河源區高寒草地土壤微生物數量特征[J]. 草原與草坪,2006,26(5):3-7
[72] 彭岳林,蔡曉布,薛會英. 退化高寒草原土壤微生物變化特性研究[J]. 西北農業學報,2007,16(4):112-115
[73] 姚拓,馬麗萍,張德罡. 我國草地土壤微生物生態研究進展及淺評[J]. 草業科學,2005,22(11):1-7
[74] 闞海明,龐卓,陳超,等. 北京西北淺山區退化草地植被恢復對土壤微生物群落多樣性的影響[J]. 草地學報,2022,30(6):1350-1358
[75] 金志薇,鐘文輝,吳少松,等. 植被退化對滇西北高寒草地土壤微生物群落的影響[J]. 微生物學報,2018,58(12):2174-2185
[76] 羅正明,劉晉仙,赫磊,等. 基于分子生態學網絡探究亞高山草甸退化對土壤微生物群落的影響[J]. 生態學報,2023,43(18):7435-7447
[77] VAN DER WAL A,VAN VEEN J A,SMANT W,et al. Fungal biomass development in a chronosequence of land abandonment[J]. Soil Biology and Biochemistry,2006,38(1):51-60
[78] 李海云,姚拓,高亞敏,等. 退化高寒草地土壤真菌群落與土壤環境因子間相互關系[J]. 微生物學報,2019,59(4):678-688
[79] 王英成,姚世庭,金鑫,等. 三江源區高寒退化草甸土壤細菌多樣性的對比研究[J]. 生態環境學報,2022,31(4):695-703
[80] HERRERA PAREDES S,LEBEIS S L. Giving back to the community:microbial mechanisms of plant-soil interactions[J]. Functional Ecology,2016,30(7):1043-1052
[81] 盧虎,姚拓,李建宏,等. 高寒地區不同退化草地植被和土壤微生物特性及其相關性研究[J]. 草業學報,2015,24(5):34-43
[82] 王啟蘭,曹廣民,王長庭. 高寒草甸不同植被土壤微生物數量及微生物生物量的特征[J]. 生態學雜志,2007,26(7):1002-1008
[83] 楊明新,陳科宇,李成先,等. 三江源區高寒草原退化對不同生長期土壤真菌群落的影響[J]. 草業科學,2024,41(1):15-25
[84] BARDGETT R D,VAN DER PUTTEN W H. Belowground biodiversity and ecosystem functioning[J]. Nature,2014,515:505-511
[85] LI Y X,WANG L X,TIAN L,et al. Dissolved organic carbon,an indicator of soil bacterial succession in restored wetland under freeze-thaw cycle[J]. Ecological Engineering,2022,177:106569
[86] LI Y M,WANG S P,JIANG L L,et al. Changes of soil microbial community under different degraded gradients of alpine meadow[J]. Agriculture,Ecosystems amp; Environment,2016,222:213-222
[87] 李強,何國興,劉志剛,等. 東祁連山高寒草甸植被特征和生物多樣性對生境的響應[J]. 草地學報,2022,30(1):169-177
[88] 秦浩,李蒙愛,高勁,等. 蘆芽山不同海拔灌叢土壤細菌群落組成和多樣性研究[J]. 生態環境學報,2023,32(3):459-468
[89] 王婷,楊思維,花蕊,等. 高寒草原植物功能群組成對退化程度的響應[J]. 生態學報,2020,40(7):2225-2233
[90] 郭金瑞,宋振偉,朱平,等. 長期不同種植模式對東北黑土微生物群落結構與土壤理化性質的影響[J]. 土壤通報,2016,47(2):353-359
[91] 張玉琪,梁婷,張德罡,等. 祁連山東段退化高寒草甸土壤水分入滲的變化及團聚體對水分入滲的影響[J]. 草地學報,2020,28(2):500-508
(責任編輯 "劉婷婷)
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
中學生數理化·八年級物理人教版(2022年3期)2022-03-16 05:55:08
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
當代陜西(2021年2期)2021-03-29 07:41:24
科技傳播(2019年22期)2020-01-14 03:06:54
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
汽車工程學報(2017年2期)2017-07-05 08:13:02
媽媽寶寶(2017年3期)2017-02-21 01:22:28
中國塑料(2016年3期)2016-06-15 20:30:00
