菊苣TCP基因家族鑒定與分析
2024-12-31 00:00:00杜洋張智韋陸爽劉雪李大勇徐朝陽王晨晨張彬許立新
草地學報 2024年8期


摘要:TCP轉錄因子參與調控植物多種生理過程,包括植物葉片發育、花發育、分枝的形成等方面。菊苣(Cichorium intybus L.)是一種優良牧草,但目前還未有關于其TCP基因家族研究的報道。本研究基于菊苣全基因組數據,利用擬南芥(Arabidopsis thaliana)、生菜(Lactuca sativa L.)TCP基因家族的氨基酸序列,對菊苣TCP基因家族成員進行鑒定篩選,最終得到39個菊苣TCP基因家族成員。接著利用生物信息學方法對這些成員進行分析發現,39個成員不均勻分布在9條染色體上;氨基酸數目為113~498,分子量為12 425.84~56 266.57Da;所有成員分為3個亞家族;均含有保守的basic-helix-loop-helix(bHLH)結構域;順式作用元件分析發現,菊苣TCP家族成員具有多種激素、非生物脅迫和生長發育響應元件;對CIN類TCP基因在菊苣各組織和高溫脅迫下進行表達分析發現,這些成員可能參與菊苣的葉片發育調控且對高溫脅迫響應。本研究可以為改良菊苣牧草品質提供理論依據。
關鍵詞:TCP基因家族;菊苣;表達分析;家族分析
中圖分類號:Q949.783.5""" 文獻標識碼:A"""" 文章編號:1007-0435(2024)08-2428-12
Chicory TCP Gene Family Identification and Analysis
DU Yang1,2#, ZHANG Zhi-wei3#, LU Shuang1, LIU Xue2, LI Da-yong2,
XU Zhao-yang2, WANG Chen-chen2, ZHANG Bin2*,XU Li-xin1*
(1. College of Prataculture and Grassland, Beijing Forestry University, Beijing 100083, China; 2. Institute of Vegetables,
Beijing Academy of Agriculture and Forestry Sciences, Beijing 100083, China;3. CCTEG Ecological Environment Technology
Co., Ltd, Beijing 100020, China)
Abstract:The TCP gene family is involved in the regulation of a variety of physiological processes in plants,including plant leaf development,flower development,and branch formation. Chicory(Cichorium intybus L.) is an excellent forage,but no studies of its TCP gene family have been reported. Based on the whole genome data of chicory,the amino acid sequences of the Arabidopsis thaliana and Lactuca sativa L. TCP gene families were used to identify and screen the members of the chicory TCP gene family,and finally 39 chicory TCP gene family members were obtained. Then,bioinformatics methods were used to analyze these members and found that 39 members were unevenly distributed on 9 chromosomes. The number of amino acids is 113~498,and the molecular weight is 12 425.84~56 266.57 Da. All members contain conserved TCP domains,namely basic-helix-loop-helix(bHLH) domains,and are divided into 3 subfamilies. The analysis of cis-acting elements showed that members of the chicory TCP family had a variety of hormones,abiotic stress and growth and development response elements. The expression analysis of CIN class TCP genes in chicory tissues and under high temperature stress showed that these members may be involved in the regulation of leaf development and response to high temperature stress in chicory. This study can provide a theoretical basis for improving the quality of chicory forage.
Key words:TCP gene family;Cichorium intybus L.;Expression analysis;Family analysis
菊苣(Cichorium intybus L.),又稱歐洲菊苣,起源于地中海地區,是菊科(Asteraceae)菊苣屬(Cichorium)多年生草本植物,在20世紀70年代末期,引入我國開始種植。作為飼用牧草,菊苣具有礦物質含量高、營養價值高、適口性好、易消化、生產率高和刈割后再生能力強等優良特性[1-2]。其營養價值類似于苜蓿,含有較高的粗蛋白和能量,礦物質含量甚至高于紫花苜蓿[3]。
TCP(Teosinte branched1/Cycloidea/Proliferating cell factor)基因家族被廣泛報道于很多物種,是一類非常重要的基因家族。家族成員均含有一個55~60堿基的TCP保守結構域,即basic-helix-loop-helix(bHLH)結構域[4]。依據這種結構域的差別,可以將TCP家族分為兩個亞家族Class Ⅰ和Class Ⅱ,Class Ⅱ進一步分為CIN和CYC/TB1兩個亞家族。Class Ⅰ亞族基因的調控作用主要表現在促進植物生長發育,但由于Class Ⅰ家族基因的冗余性導致其單突變體表型較弱甚至無表型,因此對于Class Ⅰ類基因家族的研究較少;Class Ⅱ亞族基因調控作用大多表現在抑制植物生長和細胞增殖[5]。Class Ⅱ亞族中的CIN類基因子集的AtTCP2,AtTCP3,AtTCP4,AtTCP10,AtTCP24均受microRNA319靶向抑制[6]。有研究分析了多毛番茄(S. habrochaites L.)和栽培番茄(S. lycopersicum L.)miR319家族在低溫和高溫脅迫下的表達特征,證實miR319a,b,c,d參與調控番茄耐高溫脅迫[7]。蘿卜(Raphanus Sativus L.)[8]和醋栗番茄(Solanum pimpinellifolium L.)[9]中也有類似的報道。
已經在多種物種中鑒定和分析了TCP家族,如煙草[10]、茄子[11]、荔枝[12]、刺槐[13]、紫花苜蓿[14]等。但目前還未見在菊苣中的TCP基因家族報道。TCP家族蛋白廣泛參與植物葉片形態建成[15]、脅迫響應[16]、花發育[17]、激素調節[18]等生理生化活動,因此這些轉錄因子的研究對于提高菊苣的牧草品質、對增強抗逆性有很大的幫助。
在我國,菊苣主要分布在中部和北部地區,而限制菊苣分布的主要因素之一就是溫度。菊苣植株生長的最適溫度一般在17~20℃之間,幼苗期的植株耐高溫能力稍強,生長適溫為20~25℃。有研究發現,高溫會誘導菊苣抽薹開花[19-20],進而影響其營養物質的產量,降低其作為牧草的品質。因此,本文利用生物信息學的方法對菊苣的TCP蛋白家族成員進行了鑒定篩選并分析,同時進行了高溫脅迫下的表達模式分析,以期為研究TCPs對于植物應對高溫的作用機理提供理論依據。
1 材料與方法
1.1 試驗材料及培養
試驗在北京市農林科學研究院蔬菜所(116°42′10.41″ E,39°42′1.35″ N)進行,所用菊苣品種為‘普那’(K017)。挑選飽滿種子播種于濕濾紙上,進行萌發;當芽伸長約2 cm時,轉到蛭石∶營養土為3∶1比例的基質中;5~6天后轉入大棚,自然培養。當菊苣生長至抽薹開花,取其莖、老葉、幼葉、花、萼片和薹上葉;參考相關研究[19],35℃高溫培養,取0 h,1 h,4 h,8 h,16 h和24 h的幼嫩葉片,立即放入液氮中速凍,隨后轉移至-80℃冰箱保存,以備后續實驗使用。
1.2 菊苣TCP基因家族鑒定及基本特征分析
在網站(ftp://ftp.agis.org.cn/Inulin_plants/)下載菊苣全基因組數據;在TAIR(https://www.arabidopsis.org)下載擬南芥(Arabidopsis thaliana (L.) Heynh.)TCP成員基因序列、蛋白編碼序列和全長氨基酸序列;在數據庫Phytozome v13(http://phytozome.jgi.doe.gov/pz/)下載生菜(Lactuca sativa L.) TCP成員基因序列、蛋白編碼序列和全長氨基酸序列。本文使用擬南芥和生菜的TCP基因家族成員的基因序列在菊苣全基因組數據庫進行blast比對,刪掉E值大于1e-16的基因;然后將序列上傳至在線分析軟件Batch CD-Search(https://www.ncbi.nlm.nih.gov/Structure/bwrpsb/bwrpsb.cgi),刪掉不含TCP保守結構域的基因,最終確定了39個菊苣TCP基因家族成員。利用psRNATaget(https://www.zhaolab.org/psRNATarget/)網站對microRNA319靶基因進行預測。使用TBtools[21]軟件獲得各成員在染色體上對應的位置信息,并進行可視化作圖。利用Prot Param(https://web.expasy.org/protparam/)在線工具對39個TCP蛋白序列進行分析,得出其蛋白分子量、理論等電點、氨基酸數量等理化性質數據。通過在線工具WoLF PSORT(https://wolfpsort.hgc.jp)對39個蛋白序列進行亞細胞定位預測。
1.3 蛋白系統進化分析
使用已下載好的擬南芥TCP家族蛋白、生菜TCP家族蛋白與篩選好的39個菊苣TCP基因家族蛋白序列在MEGA7.0軟件中進行系統發育樹的構建。構建時,選擇ClustalW進行多序列比對,設置參數:鄰接法(Neighbor-Joining),Partial Deletion,P-distance,Bootstrap為1000次。
1.4 基因結構,Domain,Motif分析
使用菊苣基因組GFF文件和篩選后的菊苣TCP家族的核苷酸序列,利用TBtools軟件進行基因結構的分析。將菊苣39個TCP家族成員蛋白序列上傳至在線分析軟件Batch CD-Search進行Domain分析;使用在線網站MEME(https://meme-suite.org/meme/tools/meme)進行Motif分析,motif數目設置為10。使用TBtools軟件對菊苣TCP家族基因結構、Domain,Motif分析進行可視化作圖。
1.5 蛋白質三維結構預測
將確定好的菊苣TCP家族成員蛋白序列依次上傳到蛋白質三維結構在線預測網站Swiss-Model(https://swissmodel.expasy.org),得到各成員的蛋白質三維預測結構。
1.6 順式作用元件分析
在網站下載菊苣全基因組序列與GFF文件,使用TBtools軟件,提取每個成員基因序列上游2 000 bp區域的序列。將其上傳至在線預測網站PlantCare(http://bioinformatics.psb.ugent.be/webtools/plantcare/html/)進行預測。使用TBtools軟件對預測的結果進行可視化作圖。
1.7 熒光定量PCR及數據分析
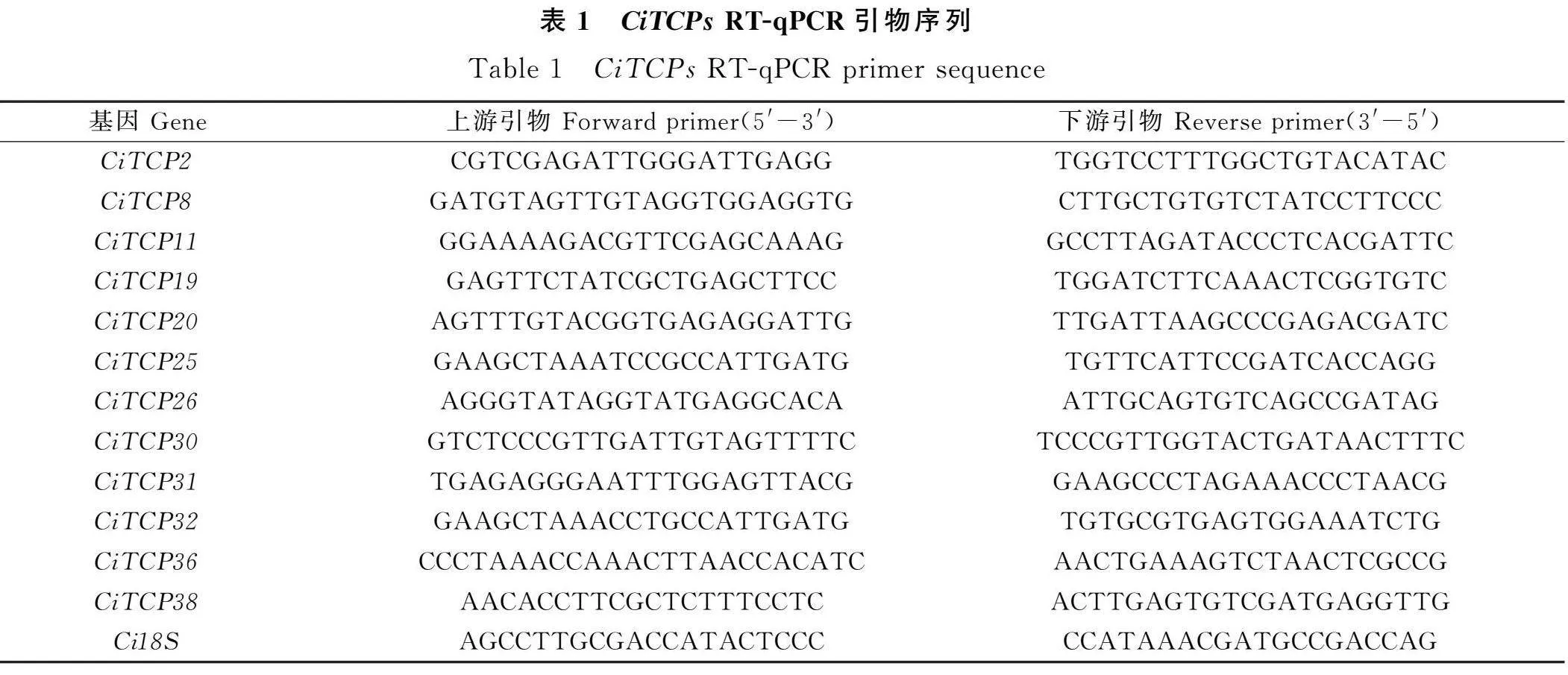
選擇18S作為內參基因,根據各成員CDS序列在在線網站設計引物(表1)。采集菊苣的莖、幼葉、老葉、花、萼片和薹上葉進行RNA提取。RNA提取和實時熒光定量PCR預混液均采用南京諾唯贊公司試劑盒進行,試驗方法以說明書為準,RT-qPCR實驗以羅氏(Roche)LC480熒光定量PCR儀進行基因表達檢測。選用2-ΔΔCt方法[22]進行數據分析,將每一個樣本技術重復3次,用“平均值±標準差”表示基因的相對表達量。
2 結果與分析
2.1 菊苣TCP基因家族鑒定及基本特征分析
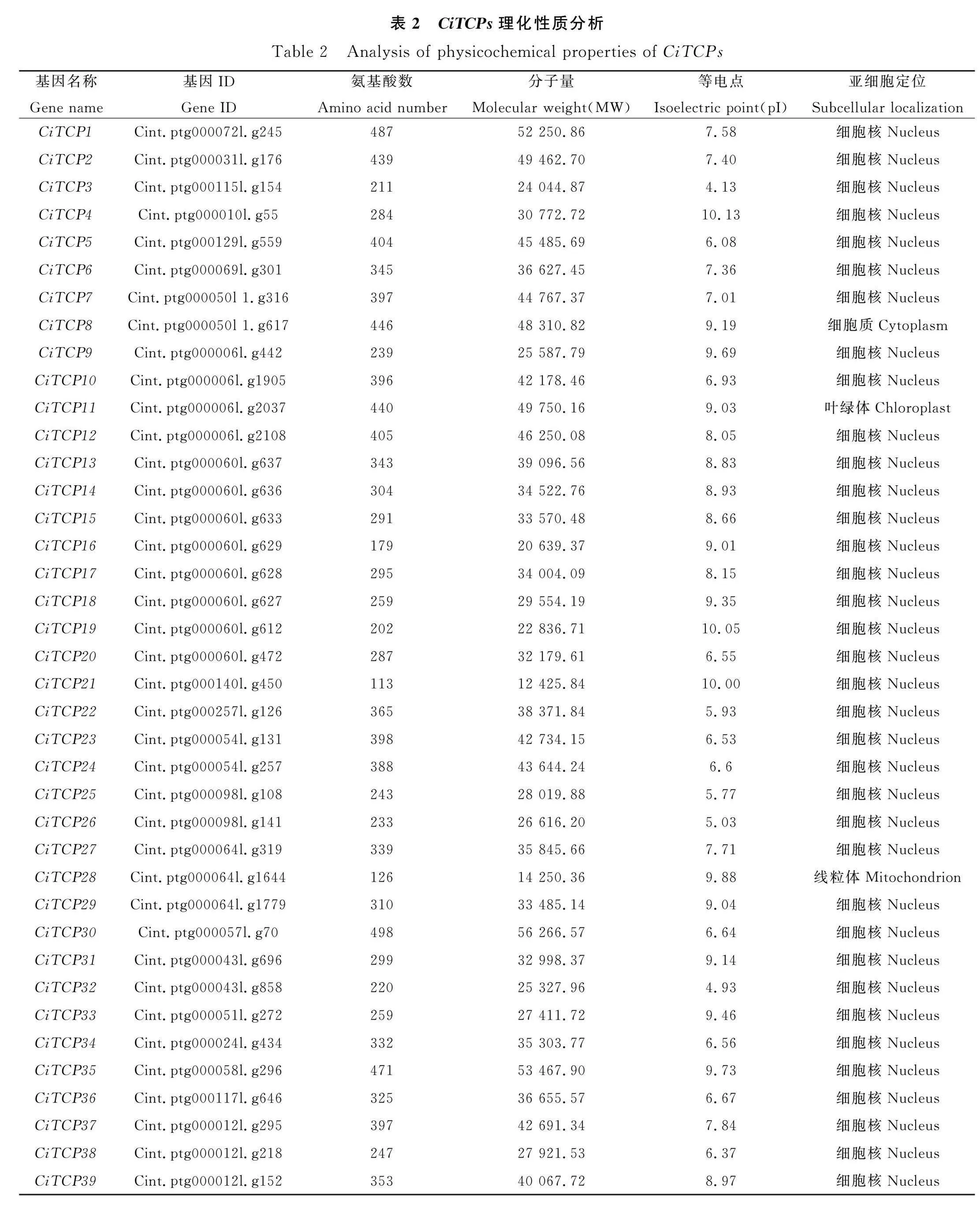
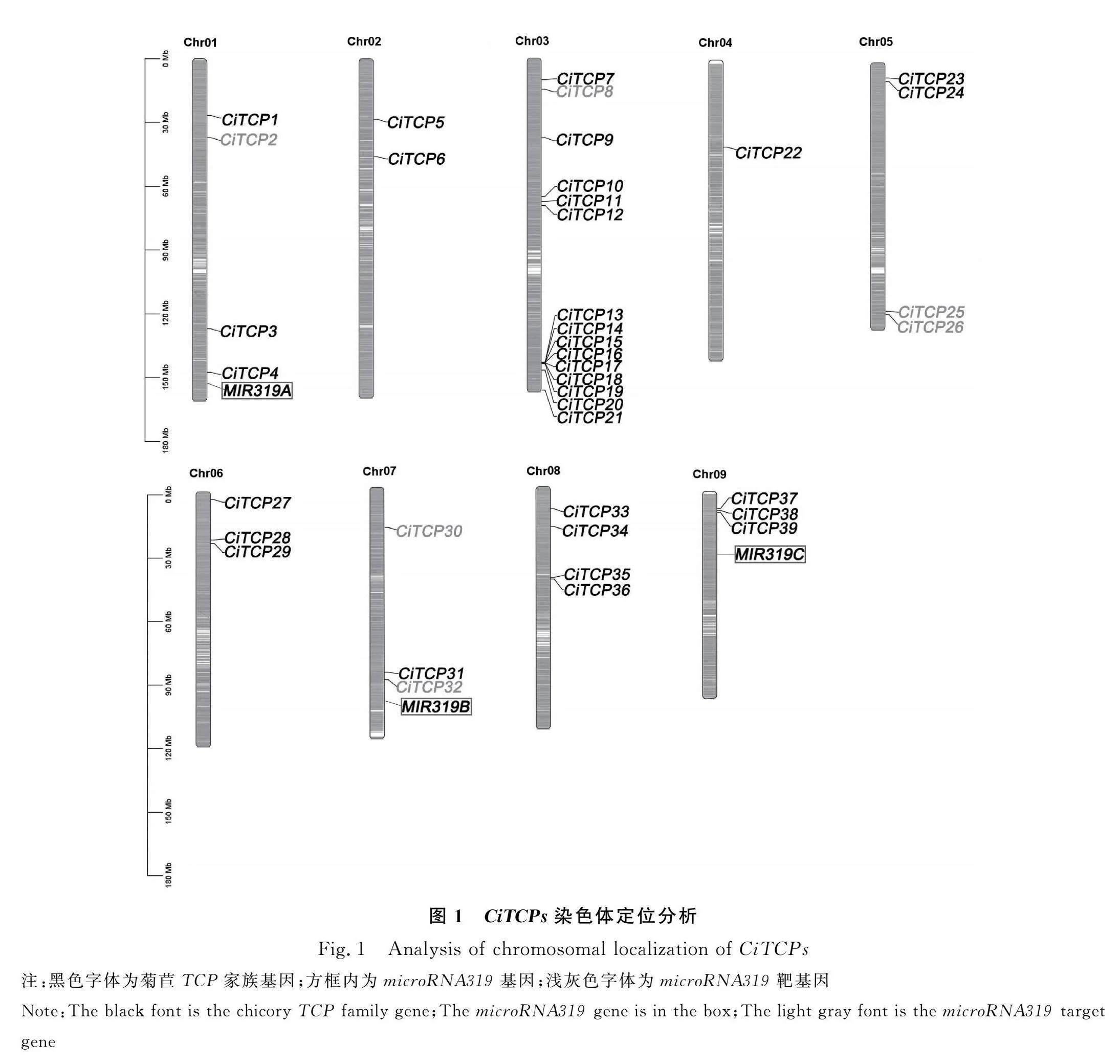
本研究以擬南芥、生菜TCP蛋白序列為檢索序列,基于菊苣全基因組數據,共鑒定出39個菊苣TCP基因(表2)。同時,使用生菜microRNA319基因序列在菊苣基因組中blast出了3個基因。對菊苣TCP基因家族成員和blast出的菊苣microRNA319基因的染色體定位信息,進行可視化作圖。39個TCP家族成員在菊苣9條染色體中呈不均勻分布(圖1)。其中,3號染色體擁有最多的TCP家族成員,有15個家族成員;4號染色體分布最少,僅有1個家族成員;其余染色體分布2~4個家族成員;microRNA319分布在1號、7號和9號染色體上,其靶基因分布在1號、3號、5號和7號染色體上。按照各成員在染色體上分布的位置順序進行命名,將39個基因命名為CiTCP1~CiTCP39;microRNA319基因命名為MIR319A,MIR319B和MIR319C。對39個菊苣TCP基因家族成員的理化性質進行分析,結果顯示:氨基酸數目為113(CiTCP21)~498(CiTCP30),對應分子量為12 425.84~56 266.57Da;所有成員均為親水蛋白,且不穩定系數均大于40;等電點介于4.13~10.13,亞細胞定位預測顯示除CiTCP8位于細胞質、CiTCP11位于葉綠體、CiTCP28位于線粒體外,其余成員均位于細胞核內(表2)。
2.2 菊苣TCP家族基因蛋白質系統進化分析
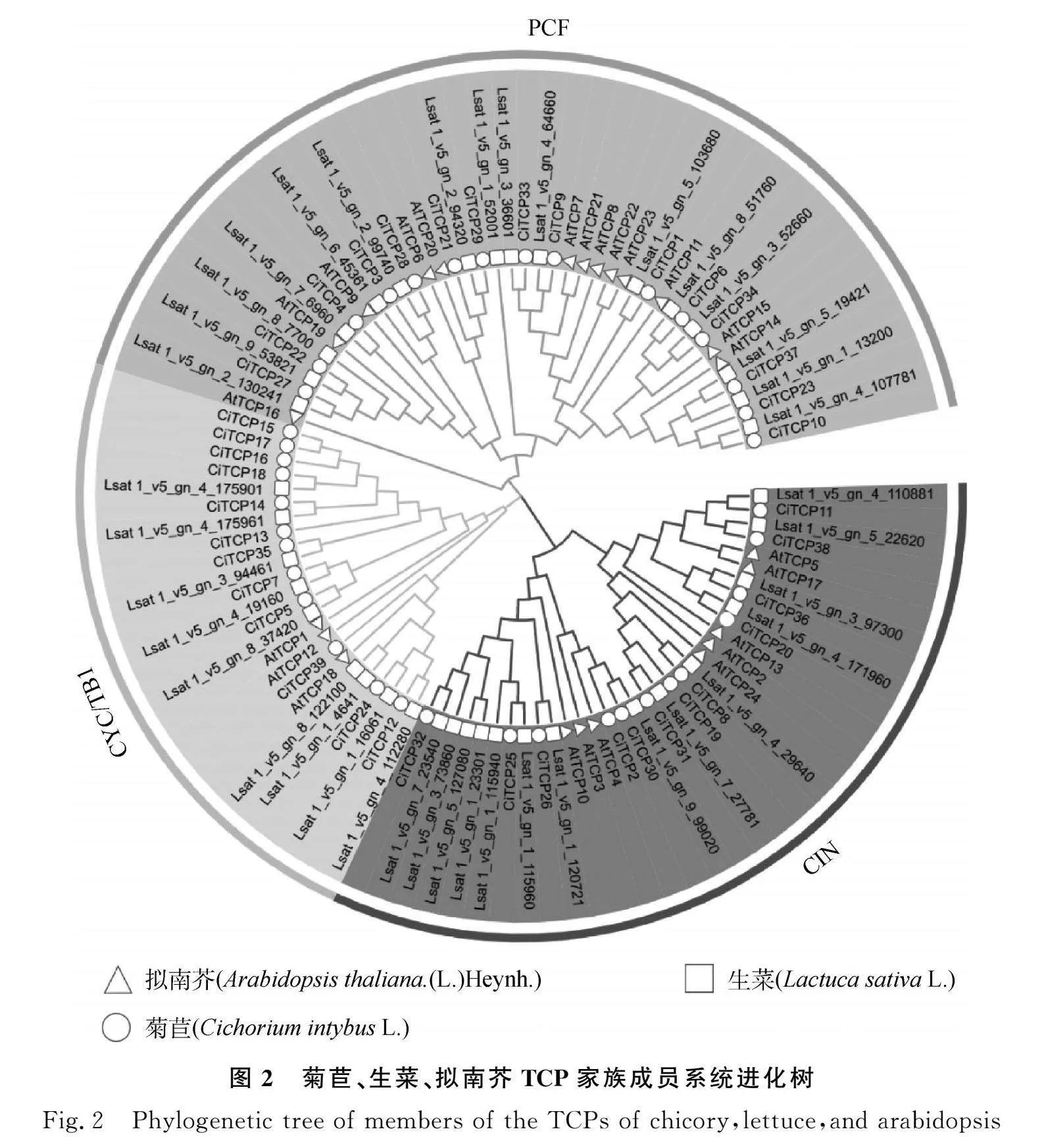
本研究利用MEGA7.0軟件,對擬南芥、菊苣和生菜中被報道的TCP家族進行系統進化分析。參考擬南芥TCP基因家族的分類,將菊苣TCP家族分為Class Ⅰ和Class Ⅱ兩類,其中Class Ⅰ又稱PCF亞族,Class Ⅱ可進一步劃分為兩個亞家族CIN,CYC/TB1。CIN,CYC/TB1亞家族中各有12個菊苣基因,PCF亞家族中則含有15個菊苣基因,是分布最多的亞家族。其中CiTCP13~18不止在染色體上的分布位置聚集,親緣關系也較為接近,推測其在植物生長發育過程中發揮相似的功能。
2.3 菊苣TCP家族Domain、基因結構、Motif分析
利用TBtools軟件與在線工具對菊苣TCP家族進行Domain、基因結構和Motif分析。Domain分析發現,39個家族成員均含有TCP保守結構域或TCP superfamily保守結構域,說明39個成員均能發揮TCP結構功能。基因結構分析結果如圖3所示,菊苣TCP家族成員基因結構較簡單,其中15個成員有內含子,所有成員均不含UTR區。Motif分析發現,所有的成員均含有Motif 1結構,說明Motif 1是TCP基因家族成員發揮功能的重要結構。Motif 4僅在PCF亞族中出現,Motif 9僅在CYC/TB1亞族中出現,推測其可能是這兩個亞家族的特有結構。
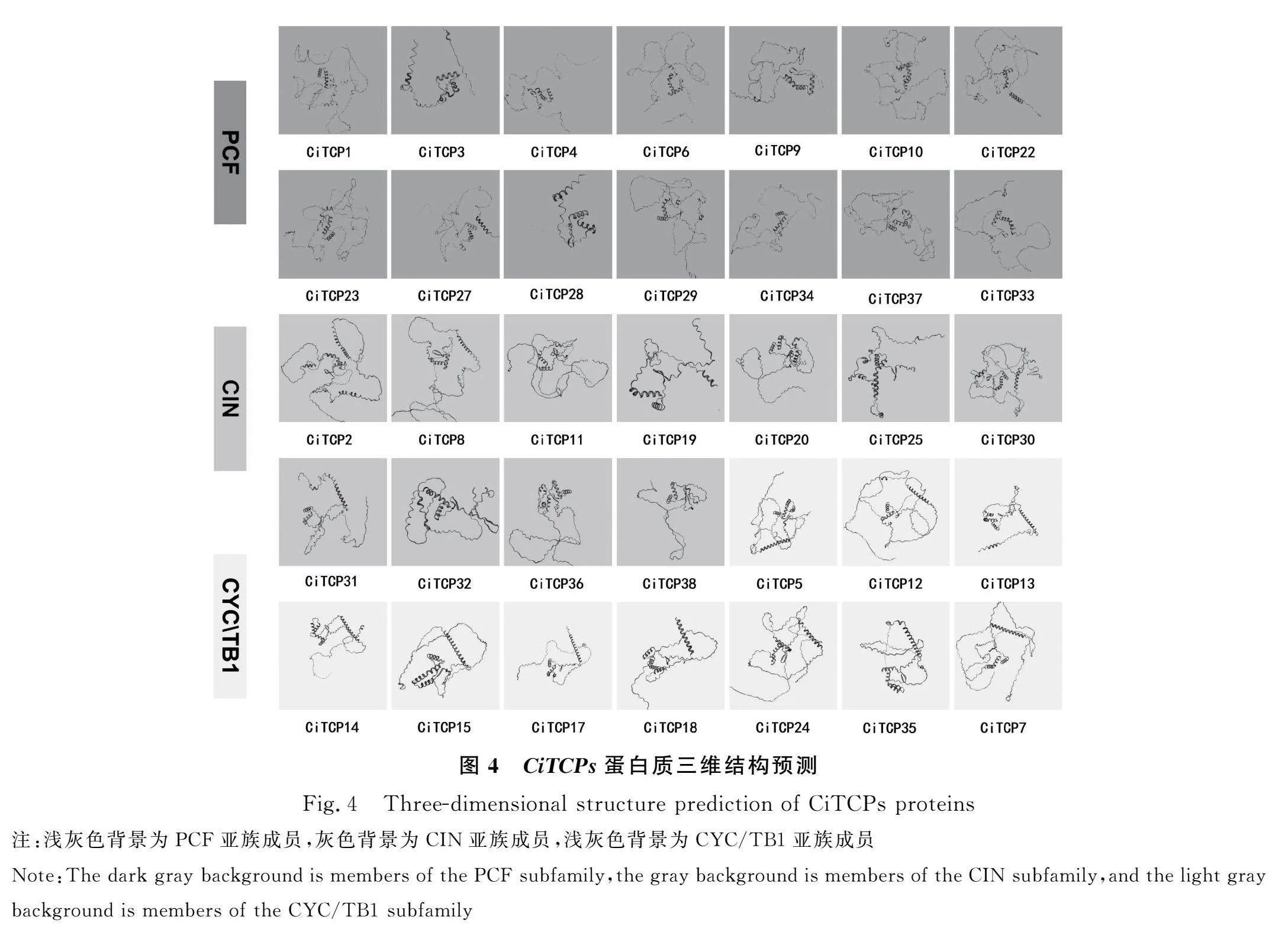
2.4 菊苣TCPs蛋白三維結構預測
上傳各成員蛋白序列至Swiss-Model在線工具,預測各成員的蛋白三維結構模型(圖4)。其中CiTCP16,CiTCP21,CiTCP26,CiTCP39的蛋白結構預測的GMQE值低于0.4,預測結構不可信。根據預測的結果,TCP家族蛋白均含有非典型的bHLH(basic-helix-loop-lelix)結構。其中PCF作為TCP家族Class Ⅰ類中的分支,明顯比另兩個亞家族中的成員結構堿性區短,CIN亞族堿性區比CYC/TB1亞族稍短但不明顯。
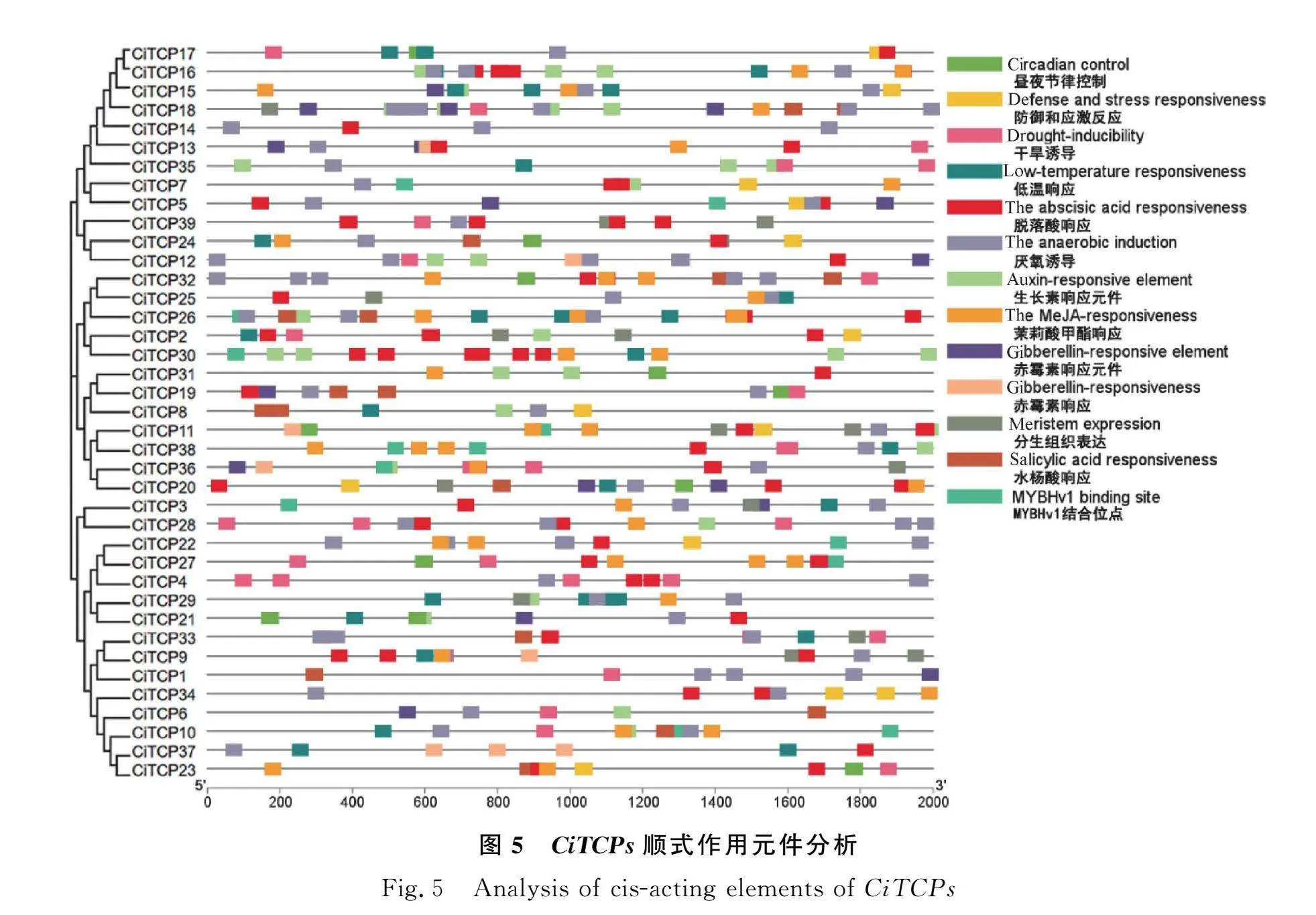
2.5 菊苣TCPs順式作用元件分析
順式作用元件在一定程度上可以反映基因的轉錄調控作用。提取菊苣各成員上游2000 bp序列,上傳到PlantCARE網站進行分析,利用TBtools進行作圖。結果發現,菊苣TCP家族成員上游啟動子區域含有與5種激素有關的作用元件。有脫落酸響應元件(31個成員)、茉莉酸甲酯響應元件(24個成員)、生長素響應元件(19個成員)、赤霉素響應元件(15個成員)、水楊酸響應元件(12個成員)。
在脅迫方面,有厭氧誘導元件(34個成員)、干旱響應元件(21個成員)、低溫響應元件(19個成員)、防御和應激作用元件(12個成員)。
在植物生長發育上,鑒定到的元件有晝夜節律控制元件(11個成員)、分生組織表達元件(12個成員)、MYBHv1結合位點作用元件(11個成員)。
綜上所述,菊苣TCP家族可能在植物激素調控、生長發育和應對環境脅迫方面發揮作用。
2.6 CIN類基因組織表達模式分析
本文對菊苣TCP基因家族中的CIN亞族中的基因進行了表達分析,結果顯示如圖6。此子集基因在各組織中的表達量差異較大,各基因在莖的表達量都極低,甚至不表達,在幼葉、老葉、花和萼片中的表達量大不相同。CiTCP25,CiTCP26,CiTCP31,CiTCP38,CiTCP19在幼葉、老葉、花和萼片中的表達量相差不大;CiTCP2在老葉中的表達量較高;CiTCP8在老葉和萼片中的表達量要稍高于幼葉和花;CiTCP11在各部位的表達量相差不大,在老葉和花中的表達量稍高于幼葉和萼片部位的表達量;CiTCP20在萼片中的表達量稍高于幼葉、老葉和花部位的表達量;CiTCP30在花中的表達量明顯高于幼葉、老葉和萼片部位的表達量;CiTCP32在花中的表達量低于幼葉、老葉和萼片部位的表達量;CiTCP36在花、萼片中的表達量較高。各成員在薹上葉的表達量均高于在其他部位的表達量,可能與各成員參與葉片發育調控有關。
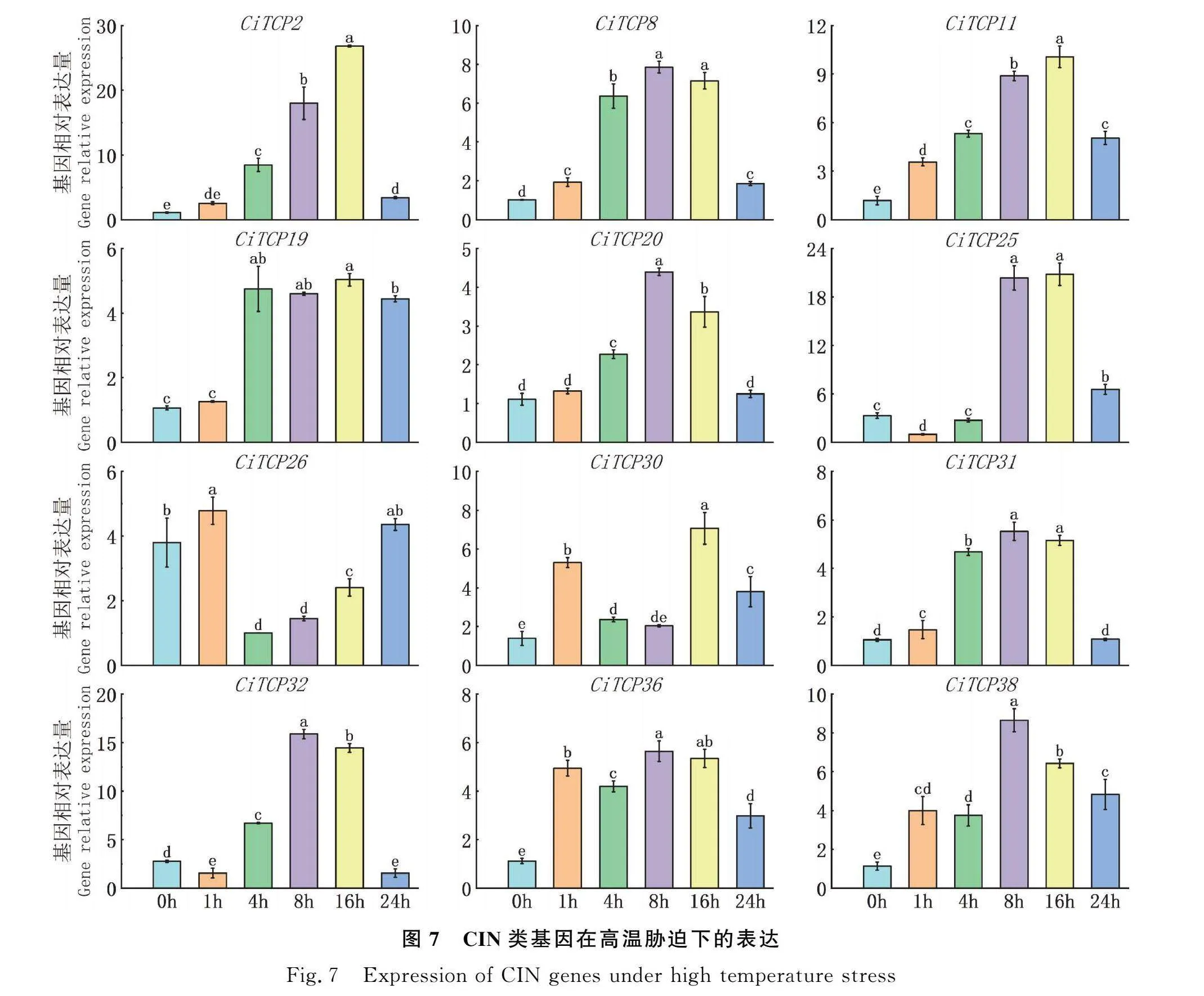
2.7 CIN類基因高溫脅迫下的表達模式分析
耐熱性較差是限制菊苣分布的重要因素,因而本文對菊苣進行了高溫脅迫處理,對CIN亞族中的基因進行了表達分析,結果顯示如圖7。各基因在高溫脅迫下的表達量均有不同程度的升高。除基因CiTCP26,CiTCP30表達量呈現先下降后上升的趨勢外,其余成員的表達量呈現緩慢上升后下降的趨勢。這種變化區別可能是由于各成員對高溫脅迫的不同的響應機制造成的。
3 討論
3.1 菊苣TCP家族基因的結構特點及功能
本研究在菊苣中鑒定到39個家族成員,與其他植物TCP家族成員[10-14]數量相比偏多,這可能與不同物種之間的基因擴張程度有關[7]。
本研究參考模式植物擬南芥的分類,將菊苣的TCP家族成員分為3個亞類:Class I(PCF),Class II(CIN)和Class II(CYC/TB1),這與前人的研究結果一致。根據蛋白系統進化分析的結果,聚類分析越是相近的基因,說明功能和結構越相似。保守基序的結果顯示,在39個CiTCPs基因中,菊苣TCP基因家族的不同亞族之間的保守基序存在差異,基因家族結構上的統一性和變異性,為菊苣TCP家族的轉錄調控作用提供了多樣性。菊苣TCP基因家族包含了大量的順式作用元件、有5種激素響應元件,脅迫相關響應元件和生長發育相關元件。有研究探討了栽培番茄和多毛番茄microRNA319在溫度脅迫中發揮的功能,高溫脅迫下,早期(1 h)miR319a,b,d表達量提高,抑制TCP3,TCP29和TCP2的表達,microRNA319可以提高植物應對溫度脅迫的能力[7]。對水稻中TCP5基因的研究發現,其對于植物干旱和鹽脅迫有所響應[23]。植物形態結構與植物激素之間的相互作用可以提高植物抗逆性,TCP4可能通過調控主根伸長參與了干旱脅迫響應[24]。這些研究與本文順式作用元件的結果一致,說明TCP基因家族的表達可能受植物生長發育、激素和環境脅迫的調控。
3.2 菊苣CIN類TCP家族基因的表達特點
TCP基因家族已經被很多研究證實參與植物的生長發育。microRNA319靶基因CiTCP2,CiTCP8,CiTCP25,CiTCP26,CiTCP30和CiTC-P32均為CIN亞族基因,因此本文選擇了CIN類基因,對其在不同組織和高溫脅迫下的表達模式進行了分析。結果發現各成員在薹上葉組織中的基因表達量均較高;CiTCP2,CiTCP8,CiTCP11,CiT-CP32,CiTCP36在老葉、幼葉、萼片中的表達量也比其他組織高,說明這些基因可能在葉片發育過程中發揮重要作用。AtTCP5參與調控葉邊緣形態[25],TCP4可以調控子葉毛狀體的發育[26]等研究都說明,這些CIN類的TCP基因可能對葉片的形態構成有重要的調控作用。
王利彬[27]對大豆在高溫脅迫應答中的關鍵轉錄因子進行了篩選,確定TCP對高溫有所響應。在本文中,高溫脅迫下大部分成員表達量均上升,在升至最高點后,呈現下降趨勢。其中,除CiTCP26,CiTCP30外其余基因表達變化趨勢較為相似,變化較為平緩,推測它們應對高溫脅迫的響應機制類似。這可能與TCP基因功能存在高度的冗余有關。CiTCP2,CiTCP8,CiTCP25,CiTCP26,CiTCP30和CiTCP32為miR319的靶基因,miR319對于靶基因的調控為負調控[15],番茄中miR319a,b,d在熱處理下的表達量呈現由高到低的變化,miR319c表達量下調[7],這與本文中這些基因的表達變化一致。
4 結論
本研究確定了菊苣39個TCP家族成員,并對這些成員基因進行分析,初步探明了其可能會在植物中發揮的功能。接著對CIN類的成員在各組織和高溫脅迫下的表達模式進行了探究,結果顯示,TCP家族CIN類成員在不同組織中表達各異,主要參與葉片發育調控;各成員均對高溫脅迫有不同程度的響應。
參考文獻
[1] LANBERT M G,CLARK D A,LITHERLAND A J. Advances in pasture management for animal productivity and health[J]. New Zealand Veterinary Journal,2004,52(6):311-319
[2] KIDANCE A,HOUDIJK J G,ATHANASIADOU S,et al. Effects of maternal protein nutrition and subsequent grazing onchicory(Cichorium intybus) on parasitism and performance of lambs[J]. Journal of Animal Science,2010,88(4):1513-1521
[3] 王若楠,曹升旭,鄭飛,等. 菊苣作為飼用牧草的研究與應用[J]. 飼料工業,2018,39(11):19-22
[4] LIN Y F,CHEN Y,HSIAO Y Y,et al. Genome-wide identification and characterization of TCP genes involved in ovule development of Phalaenopsis equestris[J]. Journal of Experimental Botany,2016,67(17):5051-5066
[5] 安琳君. 白樺BpTCP1及BpTCP2基因的功能研究[D]. 哈爾濱:東北林業大學,2020:1
[6] 邵長生,鄒春晶,鄭慧俊,等. TCP轉錄因子在植物表皮毛發育調控中的功能[J/OL]. https://kns.cnki.net/kcms2/detail/46.1068.S.20230616.1021.006.html,2023-06-16/2023-09-03
[7] 石瀟瀑. 溫度脅迫下番茄microRNA319的表達和功能分析[D]. 南京:南京農業大學,2019:47
[8] WANG R,XU L,ZHU X,et al. Transcriptome-wide characterization of novel and heat-stress-responsive micrornas in radish(Raphanus Sativus L.) using next-generation sequencing[J]. Plant Molecular Biology Reporter,2015,33:867-880
[9] ZHOU R,WANG Q,JIANG F,et al. Identification of miRNAs and their targets in wild tomato at moderately and acutely elevated temperatures by high-throughput sequencing and degradome analysis[J]. Scientific Reports,2016,6:33777
[10]張路陽,韓文龍,徐曉雯,等. 煙草TCP基因家族的鑒定及表達分析[J]. 生物技術通報,2023,39(6):248-258
[11]梅文宇,方燕芬,宮超,等. 茄子TCP轉錄因子的鑒定及脅迫處理下的表達分析[J]. 廣東農業科學,2022,49(12):20-33
[12]萬鵬龍,李建國,趙明磊. 荔枝TCP家族全基因組鑒定及miRNA靶點分析[J]. https://link.cnki.net/urlid/46.1068.S.20230829.1636.004,2023-08-30/2023-09-03
[13]米躍騏,田彥挺,楊浩,等. 刺槐TCP基因家族鑒定及表達分析[J/OL]. https://link.cnki.net/kcms/detail/46.1068.S.20230505.1238.008.html,2023-05-06/-2023-09-03
[14]魏娜,李艷鵬,馬藝桐,等. 全基因組水平紫花苜蓿TCP基因家族的鑒定及其在干旱脅迫下表達模式分析[J]. 草業學報,2022,31(1):118-130
[15]PALATNIK J F,ALLEN E,WU X,et al. Control of leaf morphogenesis by micro RNAs[J]. Nature,2003,425:257-263
[16]MARTIN-TRILLO M,CUBAS P. TCP genes:a family snapshot ten years later[J]. Trends in Plant Science,2010,15(1):31-39
[17]LUCERO L E,UBERTI-MANASSERO N G,ARCE A L,et al. TCP15 modulates cytokinin and auxin responses during gynoecium development in Arabidopsis[J]. The Plant Journal,2015,84(2):267-282
[18]LIN Y F,CHEN Y Y,HSIAO Y Y,et al. Genome-wide identification and characterization of TCP genes involved in ovule development of Phalaenopsis equestris[J]. Journal of Experiment Botany,2016,67(17):5051-5066
[19]MATHIEU A S,CLAIRE.PRILLEUX,JACQUEMIN G,et al. Impact of vernalization and heat on flowering induction,development and fertility in root chicory (Cichorium intybus L var sativum)[J]. Journal of Plant Physiology,2020,254:153272
[20]ANNE-SOPHIE,MATHIEU,BERTRAND,et al. High temperatures limit plant growth but hasten flowering in root chicory (Cichorium intybus) independently of vernalisation[J]. Journal of Plant Physiology,2014,171(2):109-118
[21]CHEN C J,CHEN H,ZHANG Y,et al. TBtools:an integrative toolkit developed for interactive analyses of big biological data[J]. Molecular Plant,2020,13(8):1194-1202
[22]LIVAK K J,SCHMITTGEN T D L. Analysis of relative gene expression data using real-time quantitative PCR and the 2-ΔΔCT method[J]. Methods,2001,25(4):402-408
[23]關紫微,曹希雅,張先文,等. 水稻TCP家族的全基因組鑒定及表達分析[J]. 分子植物育種,2022(10):3145-3156
[24]雷其冬. 擬南芥miR319-TCP4調控植物應答干旱脅迫的分子機制研究[D]. 昆明:昆明理工大學,2021:54
[25]YU H,ZHANG L,WANG W,et al. TCP5 controls leaf margin development by regulating KNOX and BEL-like transcription factors in Arabidopsis[J]. Journal of Experimental Botany,2021,72:1809-1821
[26]蘭婧秋,秦跟基. ClassⅡTCP轉錄因子的主要功能和分子調控機制[J]. 中國科學:生命科學,2021,51(11):1542-1557
[27]王利彬. 大豆苗期干旱和高溫脅迫應答機制研究及關鍵轉錄因子的篩選[D]. 哈爾濱:東北農業大學,2018:112
(責任編輯 閔芝智)
