松嫩平原賴草種群的實際營養繁殖力及芽流模型
2024-12-31 00:00:00韓大勇李海燕張維楊允菲
草地學報 2024年7期
摘要:為了評估無性系實際營養繁殖力對植物種群增長的影響,本文基于同質園建立的賴草(Leymus secalinus)實驗同生群,以單個無性系營養繁殖指標衡量實際營養繁殖力,分析了兩個年份賴草單個無性系構件生產和生長的階段性和連續性增長規律,并構建芽流模型。結果表明,賴草無性系在當年和翌年根莖總長度、根莖數、芽數等營養繁殖指標均具有較大可塑性,變異系數均大于74%。在年份間,實際營養繁殖力的增長大于營養繁殖力的增長。在根莖生長序列中,根莖總長度、根莖數、分株數、活芽總數、活構件總數的階段性和連續性增長過程均符合指數模式。在芽流中,兩個年份賴草無性系營養繁殖失敗率均低于1%,而成功率均較高,當年為40.6%,翌年為58.1%,其中,分株輸出率分別是根莖的4.3倍和5.5倍。無性系實際營養繁殖力的可塑性體現了植物種群生存對策,小無性系采取優先占據更多地上空間的策略。
關鍵詞:無性系生長;構件;實驗同生群;異速生長;休眠芽庫;活動芽庫;繁殖策略;植物生態學
中圖分類號:Q948.1""" 文獻標識碼:A""""" 文章編號:1007-0435(2024)07-1995-10
doi:10.11733/j.issn.1007-0435.2024.07.001
引用格式:
韓大勇, 李海燕, 張" 維,等.松嫩平原賴草種群的實際營養繁殖力及芽流模型[J].草地學報,2024,32(7):1995-2004
HAN Da-yong, LI Hai-yan, ZHANG Wei,et al.Actual Vegetative Propagation Ability and Bud Flow Model of Leymus secalinus Population in Songnen Plain,China[J].Acta Agrestia Sinica,2024,32(7):1995-2004
收稿日期:2023-11-27;修回日期:2024-02-19
基金項目:國家自然科學基金項目 (32171682,31472134)資助
作者簡介:
韓大勇(1978-),男,漢族,吉林榆樹人,博士,教授,主要從事植物種群動態與調節研究,E-mail:411430667@qq.com;*通信作者Author for correspondence,E-mail:yangyf@mail.nenu.edu.cn
Actual Vegetative Propagation Ability and Bud Flow Model of Leymus secalinus
Population in Songnen Plain,China
HAN Da-yong1, LI Hai-yan2, ZHANG Wei1, YANG Yun-fei1,2*
(1.School of Biology Science and technology, Yili Normal University, Yining, Xinjiang 835000, China;2.Key Laboratory of
Vegetation Ecology, Ministry of Education, Northeast Normal University, Changchun, Jilin Province 130024, China)
Abstract:To evaluate the impact of actual vegetative propagation ability (AVPA) of clones on population growth,an experimental cohort of Leymus secalinus was established under common-garden conditions. AVPA was estimated by using the vegetative reproductive indexes of a single clone. This phased and continuous growth patterns of module production and growth based on two-year experimental observations of the cohort of L. secalinus were analyzed. A bud flow model was also constructed based on the findings of the experiment. The vegetative reproductive indexes such as total length of rhizomes,number of rhizomes,and number of buds in the experimental cohort clone of L. secalinus in the same year and the following year all had significant plasticity,with coefficients of variation greater than 74%. The increase in AVPA between years was greater than that of vegetative propagation ability in the experimental cohort clones of L. secalinus. In the rhizome growth sequence,the total length of rhizomes,number of rhizomes,number of ramets,total number of active buds,and total number of live modules all follow a classic exponential growth pattern during the phased and continuous growth processes. In the bud flow,the failure rate of vegetative propagation of the experimental cohort in the two years was less than 1%. The success probability of vegetative reproductive was relatively high,with 40.6% in the first year of transplantation and 58.1% in the second year. Among them,the output rates of ramets were 4.3 times and 5.5 times that of the rhizome,respectively. The actual vegetative fecundity plasticity variations of AVPA within the experimental cohort clones of L. secalinus reflects survival strategies of population,while small clones prioritize occupying more above ground space.
Key words:Clone growth;Module;Experimental cohort;Allometric growth;Dormant bud bank;Active bud bank;Reproductive strategy;Plant ecology
繁殖是天然種群更新和持續發展的基礎[1-2]。植物的繁殖有兩種方式,一種是有性繁殖,一種是營養繁殖。在兩種繁殖方式中,種子是通過有性繁殖補充更新的潛在種群,芽是營養繁殖補充更新的潛在種群[3]。植物均為構件生物,廣義的構件是指植物學概念的枝條、葉片、莖和根等器官組分,狹義的構件是對無性系植物而言的具有獨立生活能力的全部營養繁殖體組分[4-6]。許多草本植物,特別是天然草地的優勢種,通常都是無性系植物,這些植物在適宜的生境中主要依靠營養繁殖持續更新和占據空間,經常形成單一優勢種群落[7]。以往對無性系植物的營養繁殖力多有報道[8-12],但尚未見有關實際營養繁殖力的報道。
溫帶草地四季分明,群落組成的全部現實種群每年都要更替,基本都是從芽庫中得到補充更新。由于天然草地種類組成和營養繁殖類型均多樣,芽的死亡時間難以判斷,所以,長期以來多有芽庫的研究報道[13],而極少有關于芽流的研究報道[3]。種群生態學特征是個體相應特征的統計量,種群生態學理論中許多概念的解釋就是一種方法和結果的文字描述,如年齡譜、生殖分配等[14]。種群生態學理論就是在不斷提出新概念同時創建新方法,從而促進學科發展。由于植物個體在生長過程中經常發生被昆蟲局部采食,或大風折斷個別枝條,或冰雹打掉部分葉片等,以及土壤微環境的差異,即使是同生群也會在數量性狀上表現出生態可塑性。意味著在種群增長過程中既有遭遇到不利影響的個體,也會有生長和繁殖潛力得到充分釋放的個體,造成實際營養繁殖力在個體間發生較大變異。目前,尚未建立定量化界定與分析無性系植物實際營養繁殖力的方法。
賴草(Leymus secalinus)為長根莖型禾草,是典型的無性系植物,在我國北方草地廣泛分布,經常在砂質土壤生境形成單優群落。賴草具有耐寒、耐旱、耐瘠薄、營養繁殖力強等生物學與生態學特性,是有關無性系植物生態學研究的理想材料。本研究是在同質園初始移植單生根莖芽苗,建立賴草同生群實驗小區,連續兩年,每年于生長季末期挖掘整個無性系取樣測定。營養繁殖是無性系植物的重要特征,以往研究大多通過計算各無性系營養繁殖指標的平均值來衡量種群的營養繁殖力[8,11-12],并作為衡量種群增長的標準。考慮到在營養繁殖數量上的可塑性問題,意味著種群中不同無性系的營養繁殖力有的受阻,有的則得到充分發揮,并最終影響種群增長率。為此,提出并界定無性系實際營養繁殖力有助于更好的反映不同大小無性系的實際貢獻和影響,能夠更真實地了解種群的增長過程。本研究重新設定根莖總長度作為無性系大小的衡量指標,對賴草無性系大小和構件組分數量分別進行了兩個年份階段性和連續性的生長規律分析,從構件數量特征與無性系大小關系的規律和方程參數解析種群的階段性調節和發展策略,在種群水平上,考慮到所有現實構件都是由營養繁殖的芽形成,所以根據構件組成計算芽的周轉,構建了芽流模型。本研究開拓了植物營養繁殖和芽流研究的新模式,其研究方法和理論分析也可為相關研究提供參考,具有重要的理論意義。
1" 材料與方法
1.1" 研究地點的氣候與土壤概況
本項研究是在東北松嫩平原南部東北師范大學吉林松嫩草地生態系統國家野外科學觀測研究站的栽培實驗地進行。地理位置為44′38′N,123′41′E附近。行政區劃隸屬于吉林省長嶺縣種馬場。屬于溫帶半干旱半濕潤區,具有典型的季風氣候特征,年平均溫度4.6℃ ~6.4℃,1月份的平均溫度為-20.6℃,7月份的平均溫度 28.2℃,無霜期120~140 d;年平均降水量 400~500 mm,主要集中在6—8月份。年蒸發量 1 200~1 400 mm,是降水量的3倍[7]。實驗地原本是農田。土壤類型為砂壤土。表層20 cm 厚的土壤是均質的,全氮為1.01 g·kg-1,有機碳為6.23 g·kg-1,全磷為0.74 g·kg-1,pH 值為8.37,電導率為70.85 μs·cm-1[15-16]。
1.2" 研究材料與實驗小區的設計
在天然草地自然生長的賴草種群中分株由不同齡級組成[17]。據觀察,賴草的營養株不拔節,其生長點位于地表以下,可以安全越冬和多次返青,保持分蘗節的多年生。分蘗節也經常生長出分株而呈叢生狀。因此,采用移栽分株時,必須考慮研究材料的初始條件,包括分株形態、大小和生活力等保持相對一致。選定由上一個生長季末期形成根莖節單生幼小芽苗為實驗材料,每個根莖節芽苗兩端分別帶有2 cm小段根莖。于5月初建成20 m×10 m的實驗區,移栽的行距和株距為2 m×2 m,在實驗小區內建立50個賴草無性系同生群。定期清除雜草,但未做施肥、澆水和防蟲害等田間管理。
1.3" 取樣與測定方法
于移植當年生長季末期的10月初,采用整個無性系挖掘的方法,對50個無性系進行間隔取樣,共挖取25個無性系。于第二年10月初再挖取剩下的25個二年生無性系。取樣時將地上分株連同地下根莖一并挖出,注意保持地上部分與根莖的自然聯系。按照無性系分別標記裝袋。對于第二年的樣本,根據形態和色澤先將當年生和二年生的根莖分開,再計數和測量共同數量特征指標。逐個無性系將分株、根莖和芽分別記數,測定根莖的累積長度,記錄每個根莖節芽和分蘗節芽的輸出形式。其中,分蘗節和根莖節向上生長頂端變綠的幼苗均計入分株中,具有明顯生長跡象的芽為活動芽,不具有明顯生長跡象的芽為休眠芽,二者合計視為活芽,相應地,失去生活力而死亡的芽視為死芽。將根莖和分株在80℃下烘干至恒重后,用1/1 000天平稱其生物量。
1.4" 數據處理和各種指標計算方法
用SPSS 17.0 和Origin 2022統計軟件分析數據和制圖,對兩個年份賴草同生群樣本做最大值、最小值、平均值等基本參數統計和變異系數計算,用以反映各數量特征的絕對和相對變化。本研究中,分別將以全部無性系和以單個無性系為統計單位獲得的相應的各營養繁殖器官(根莖或芽)數量指標值作為營養繁殖力和實際營養繁殖力的衡量標準。
在賴草無性系生長的定量分析中,以單個無性系為統計單位,采用根莖累積長度作為衡量無性系大小的尺度,以根莖總長度為基準的升序排列視為一個無性系的生長序列,定量分析無性系根莖數、分株數、芽數和活構件總數等在各年份的階段性及其連續性的生長規律,了解無性系實際營養繁殖力的增長特征。活構件總數為根莖數、分株數和活芽總數之和。分別以當年生和二年生以及兩個年份合并樣本做無性系大小與不同構件數量和生物量的相關性分析和顯著性檢驗,在線性、冪和指數函數中選用確定系數最大的方程作為其相關性規律變化模型。
芽流的計數方法:芽流為芽庫各組分的輸入與輸出比例。為了消除無性系樣本內構件數量的差異,兩個年份分別將25個樣本構件數量合計后再計算各組分的比率。以當年生的全體構件數量計算輸入量和輸入率;其他按各組分實際測定的芽數量計算在芽庫中總芽數所占的比率。芽庫中總芽數為庫存芽數和輸入芽數的總和。各比率的具體計算參見楊允菲等[3]。
2" 結果與分析
2.1" 營養繁殖和物質生產的可塑性
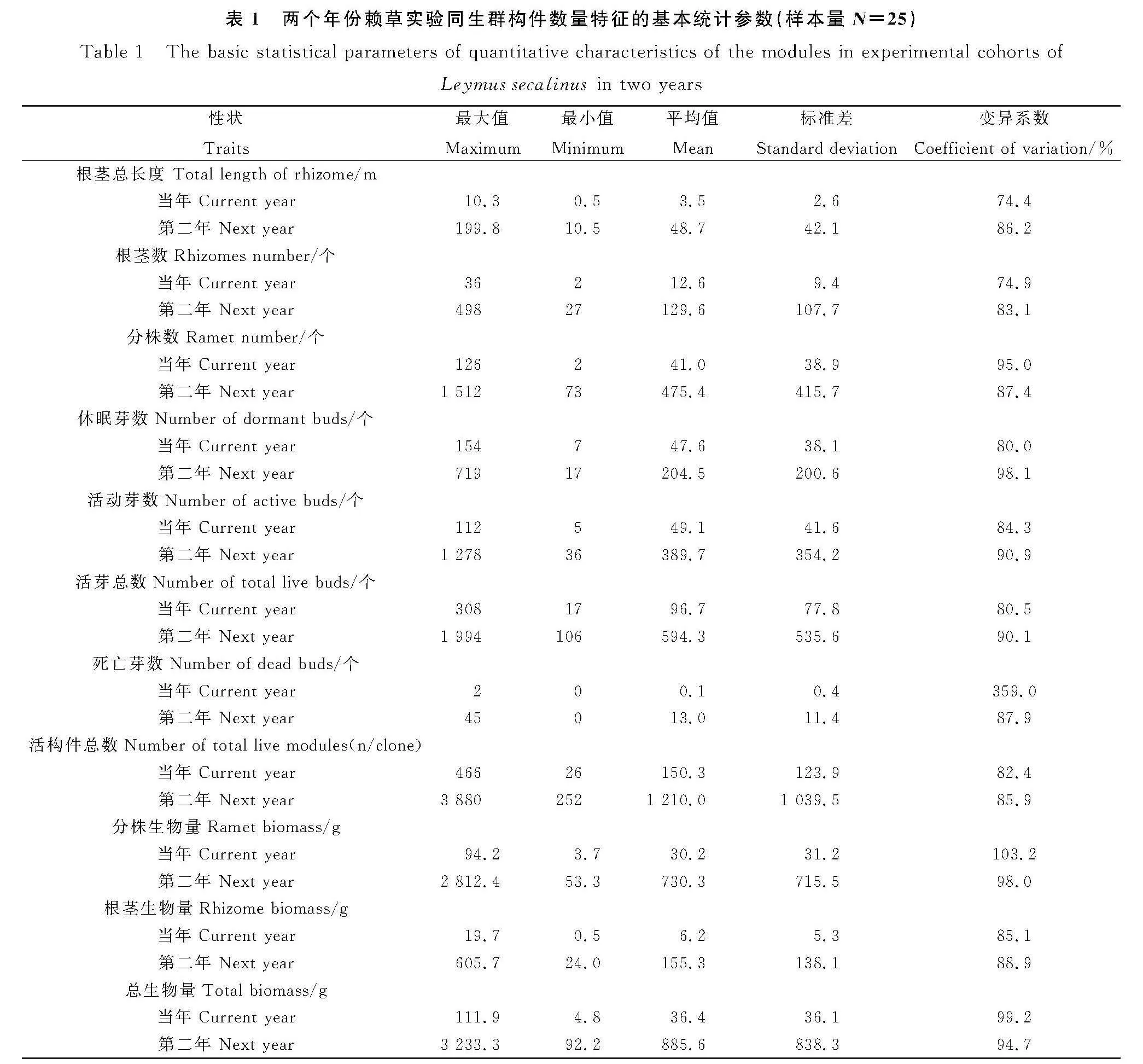
在以無性系為單位的統計樣本參數中,兩個年份賴草構件的11個數量特征,其最大值與最小值的極差普遍較大,變異系數均大于74%(表1)。其中,根莖總長度和根莖數的變異系數均小于分株數和各類芽數的。年份間平均根莖總長度增長了12.9倍,平均根莖數增長了9.3倍,平均根莖生物量24.0倍;平均分株數增長了10.6倍,平均分株生物量增長了23.2倍。由此表明,在沒有種內和種間競爭的條件下,兩個年份賴草實驗同生群各構件數量特征均具有較大的可塑性,年份間構件生物量的增加均大于構件數量的。
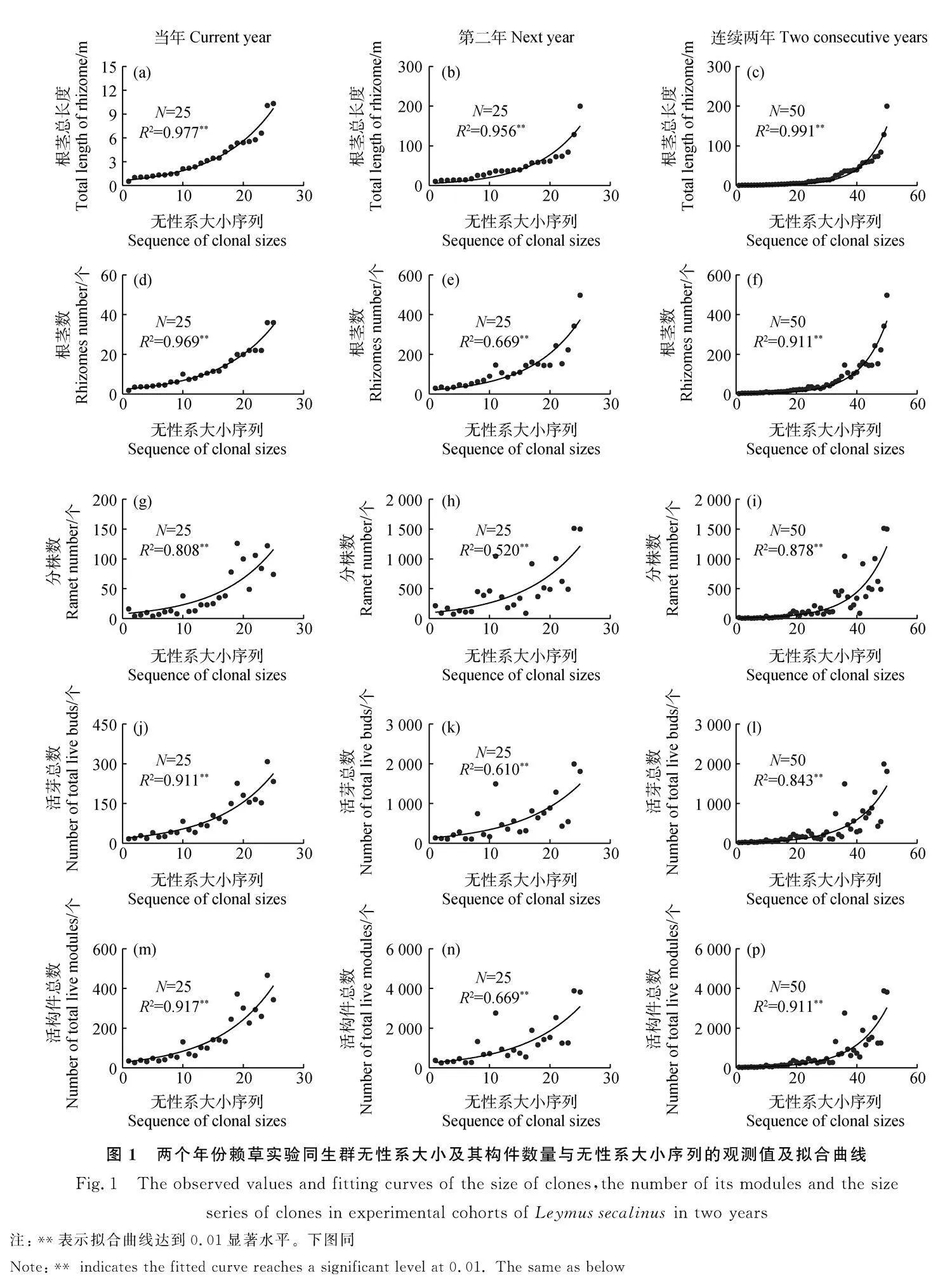
2.2" 無性系大小及其構件數量的階段性和連續性生長過程
經回歸分析和顯著性檢驗,在以根莖總長度為基準升序排列的無性系生長序列中,隨著序號增加,賴草無性系大小,即根莖總長度在當年生長階段(圖1a)、第二年生長階段(圖1b)和兩個年份的連續生長(圖1c)均呈指數增長,各種構件數量,包括根莖數(圖1d-1e)、分株數(圖1g-1i)、活芽總數(圖1j-1l)和活構件總數(圖1m-1p)在各年份的階段性和在兩個年份的連續性的生長規律均為指數形式,其回歸方程均達到了極顯著水平(Plt;0.01),確定系數(R2)介于52.0%~99.1%之間。由此表明,在沒有種內和種間競爭條件下,兩個年份賴草實驗同生群的無性系大小和各構件數量均符合經典的種群指數增長規律。
2.3" 構件數量特征與無性系大小的異速生長關系
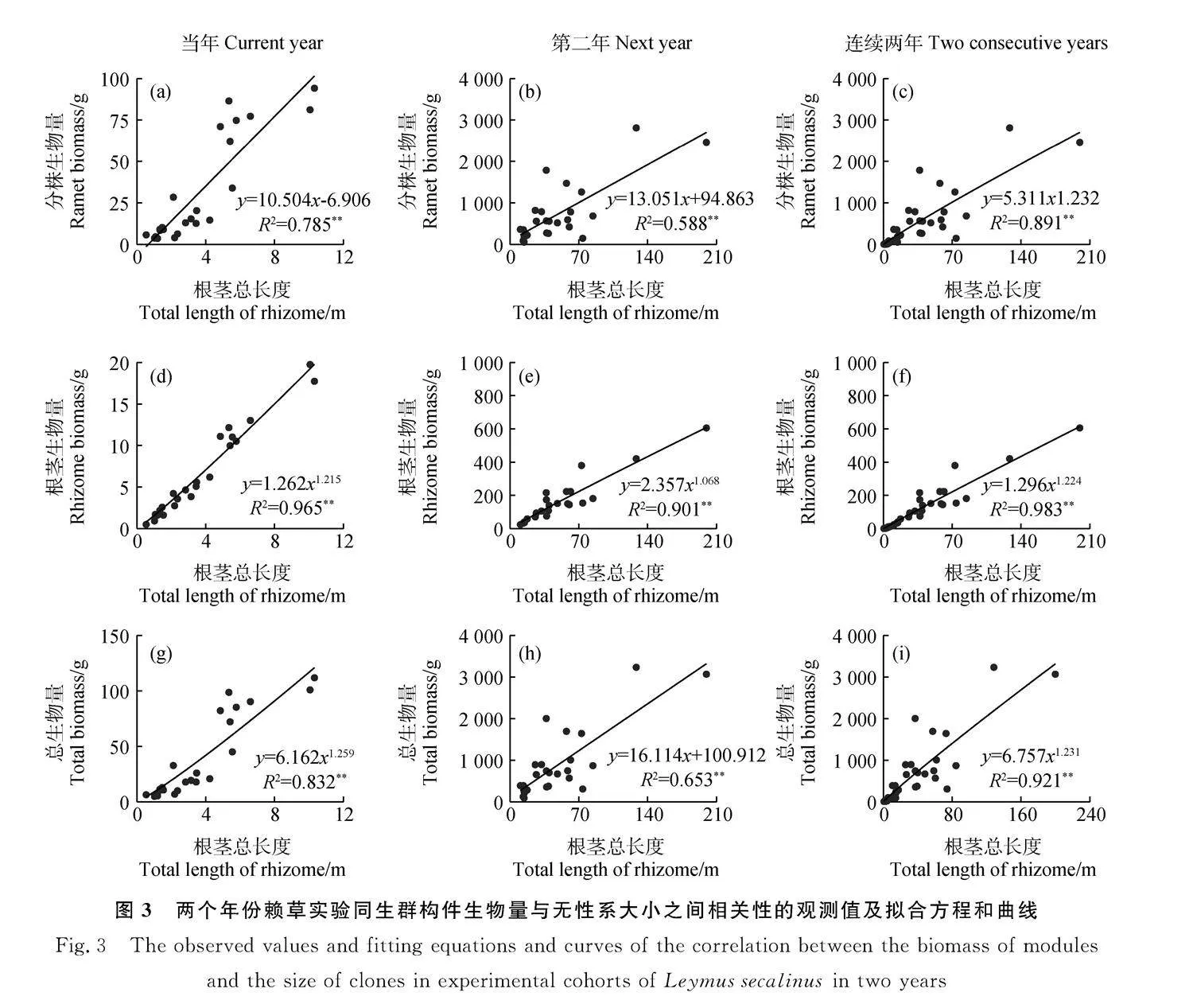
經相關性分析和顯著性檢驗,隨著無性系大小,即根莖總長度的增加,不同構件數量包括分株數(圖2a-2c)、活芽總數(圖2d-2f)和活構件總數(圖2g-2i)在當年生長階段、第二年生長階段和兩個年份的連續生長均呈冪函數增加的確定系數(R2)最大,其相關方程均達到了極顯著水平(Plt;0.01),確定系數在61.1%~92.6%。由此表明,在沒有種內和種間競爭條件下,兩個年份賴草實驗同生群的各構件數量與無性系大小之間的關系無論是階段性的還是連續性的均為經典的冪函數異速生長規律。
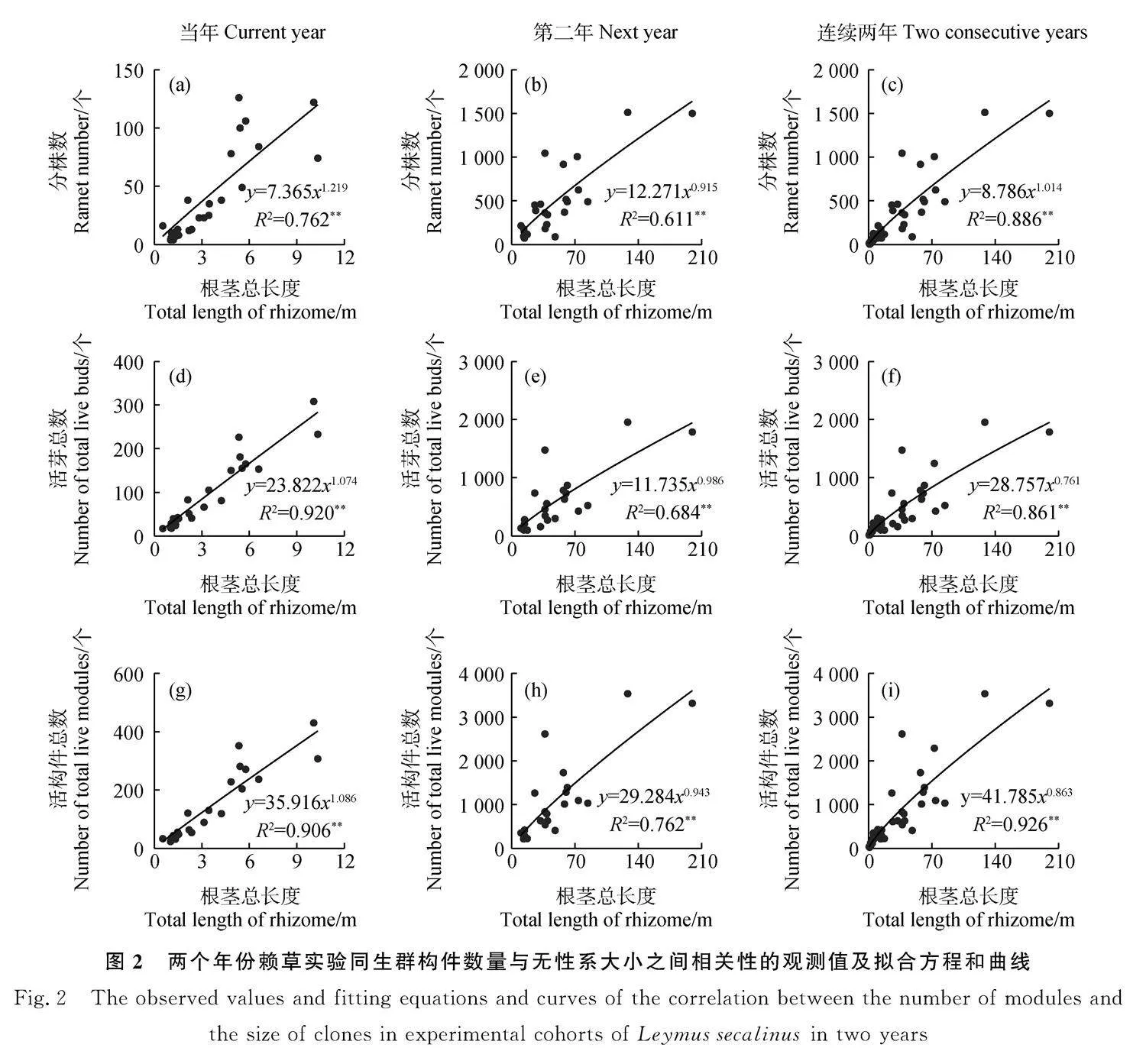
經相關性分析和顯著性檢驗,隨著根莖總長度的增加,分株生物量在當年生長階段(圖3a)、第二年生長階段(圖3b)和總生物量在第二年生長階段(圖3h)均呈直線函數增加確定系數(R2)最大;其他如分株生物量在兩個年份的連續生長(圖3c)、根莖生物量在當年生長階段(圖3d)、第二年生長階段(圖3e)和兩個年份的連續生長(圖3f)、總生物量在當年生長階段(圖3g)和兩個年份的連續生長(圖3I)均呈冪函數增加確定系數最大;其相關方程均達到了極顯著水平(Plt;0.01),確定系數(R2)在58.8%~98.3%。由此表明,在沒有種內和種間競爭條件下,兩個年份賴草實驗同生群的各構件生物量與無性系大小之間在階段性上既有線性同速生長,也有冪函數異速生長,但是在其連續性生長過程中均為經典的冪函數異速生長規律。
2.4" 種群營養繁殖的芽流模型構建及其動態分析
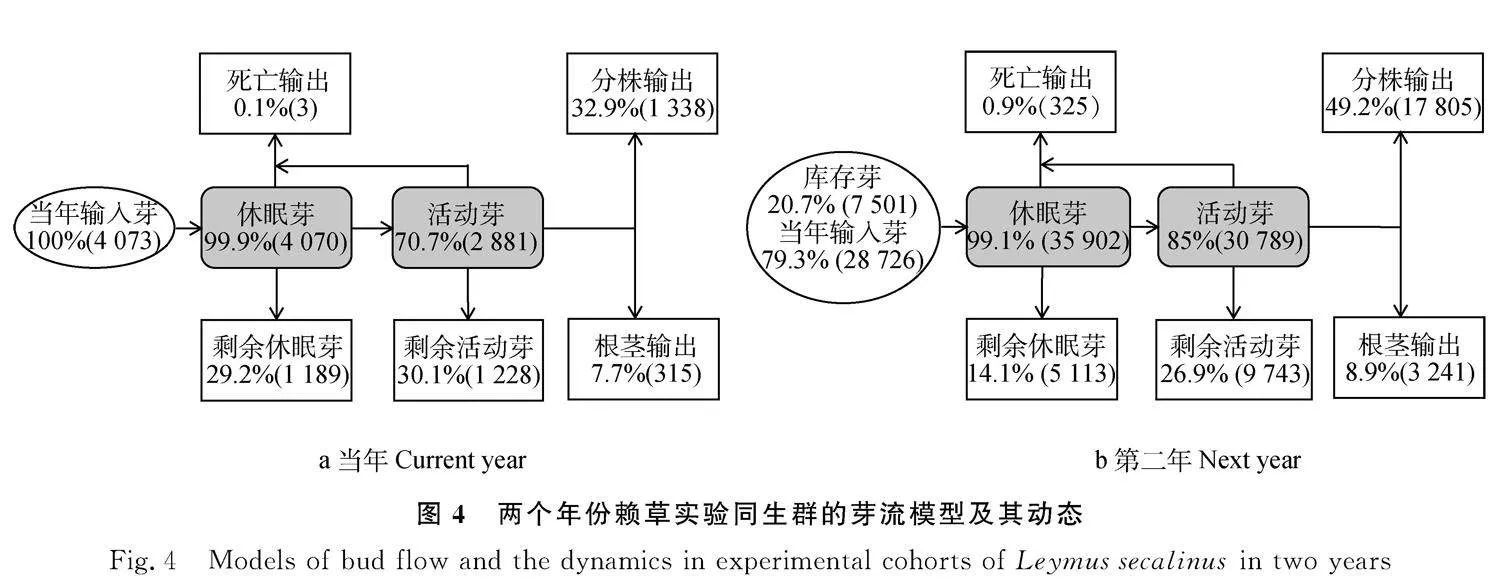
在賴草實驗同生群中,分株和根莖都是由營養繁殖的芽產生的。因此,在構建芽流模型時,把芽庫作為核心項,芽庫一般由休眠芽和活動芽組成,有輸入項和輸出項;將各組分配上相應的數據和比率,便形成一個既有流動量,又有流動率的芽流模型。經統計,賴草實驗同生群經過一個生長季的營養繁殖,全體樣本(25個無性系)共產生4 073個芽輸入芽庫中,輸入率為100%。整個生長季都有芽輸出。輸出有三個途徑,由休眠芽轉入活動芽后有些向下生長形成根莖輸出占7.7%,有些向上生長形成分株輸出占32.9%,另一個就是死亡輸出,休眠芽和活動芽都有可能生長受阻死亡,占0.1%,生長季末期芽庫中還剩余29.2%的休眠芽和30.1%的活動芽(圖4a)。經過兩個生長季的營養繁殖,將全體樣本(25個無性系)二年生根莖的7 501個芽作為庫存量,將當年生根莖產生的28 726個芽作為輸入量,輸入率為79.3%,原庫存率為20.7%。在合計的36 227個芽庫中,由分株輸出占49.2%,由根莖輸出占8.9%,死亡輸出占0.9%,生長季末期芽庫中還有14.1%的休眠芽和26.9%的活動芽(圖4b)。在兩個年份間,芽庫均以分株輸出率最大,死亡輸出最小;三種輸出率均以第二年的普遍高于當年的(圖4)。由此表明,在沒有種內和種間競爭條件下,隨著定居時間的進程,賴草實驗同生群營養繁殖中更加優先產生分株,不僅在物質生產上保證供給根莖生長等生命活動,也在占據著更多的地上空間。
3" 討論
3.1" 構件數量可塑性與實際營養繁殖力
植物數量性狀均具有生態可塑性,其可塑性大小是適應性的重要指標。由于植物生長的生境異質性程度高,任何一種生境也應該存在不同的實際營養繁殖力。本研究以無性系為計測單位,將所有無性系產生的營養繁殖器官(根莖或芽)的數量指標的平均值視為營養繁殖力,將單個無性系產生的營養繁殖器官(根莖或芽)的數量指標視為實際營養繁殖力,據此可利用最大值與最大值、最小值與最小值的比值來估計和比較實際營養繁殖力在年際間的增長狀況。譬如賴草無性系第二年的營養繁殖力是當年的13.9倍,而實際營養繁殖力最大值增長了19.4倍,最小值增長了21.0倍,等等。總體看,年份間營養繁殖力的增長倍數在6.1~13.9之間,實際營養繁殖力的增長倍數最大的在6.5~19.4之間,最小的在6.2~36.5之間。表明在沒有種內和種間競爭的條件下,兩個年份賴草實驗同生群無性系實際營養繁殖力的增長大于營養繁殖力的增長,其中,最小無性系的實際營養繁殖力增長程度更大,尤其產生分株的潛力是最大無性系的2.6倍。由此也表明,即使是同生群,不同無性系對其內部構件結構均有各自調節,最小無性系在第二年所擁有的空間相對更大,其優先產生分株,既可以有效占據地上空間,也可以保證無性系養分生產的供給,此外,分株也具有產生根莖等營養繁殖功能。如果探究這各自調節的機理,無不蘊含著與種群一樣的無性系生存與發展策略。
3.2" 無性系大小的尺度及其與構件數量增長模式
植物種群大小的唯一衡量標準是個體數量的多少。但無性系大小的尺度在以往研究中則具多樣性,除了數量外,還有用分蘗叢徑[18]、分蘗叢占據的面積[19]、根莖擴展空間的直徑和面積[20]等,都可以有效地定量描述無性系構件的數量增長和物質生產過程。由于根莖型無性系植物擴展迅速,在一、兩年之內所占據的斑塊就已經很大,即使在撂荒地或水淹跡地恢復演替植物重新定居初期,為了避免破壞,一般采用測量面積和計數地上分株的方法,很難進行整個無性系挖出做全部構件組成的數量與生物量測量。本研究設計的為期兩年,建立的賴草實驗同生群既無種間競爭,又尚未產生種內競爭的全體無性系分兩次破壞性完整取樣,獲得了全部構件組分數量特征的測定,重新界定以根莖總長度作為無性系大小的尺度,獲得了賴草無性系大小及構件組分的數量在階段性和連續生長過程均為經典的指數增長模式。
3.3" 構件數量特征之間同速和異速生長過程的變化及生長策略調節
研究發現,無論廣義還是狹義構件的數量特征之間的關系均可以定量地表達出在相對動態中的調節過程與生長策略[21-25]。本研究中賴草分株和芽兩個組分及其合計構件數量與無性系大小之間,在兩個年份的生長階段和連續生長過程均為相同的冪函數增長模式(圖2)。按照冪函數的性質,將其雙對數直線化后b值不變,對角線的斜率b=1,當bgt;1時,縱坐標的指標增長更快,當blt;1時,橫坐標的指標增長更快。依此判斷,當年生長階段方程的增長速率均為bgt;1,其中分株數的最大,表明賴草無性系在移植當年是優先產生營養繁殖構件數量,尤其優先產生分株數量;第二年生均為blt;1,表明第二年轉向擴大無性系空間優先生長根莖長度;連續生長過程分株數的bgt;1,芽數和構件總數為blt;1,表明在連續生長過程中仍然體現優先產生分株數量。而在生物量分配上,在當年和第二年的生長階段,既有同速生長過程,又有異速生長過程,而在連續生產過程中又均趨同于冪函數異速生長,這一方面反映了冪函數是植物構件之間異速生長的普遍規律,另一方面也反映了線性函數只是異速生長中的某一個環節,也是不同階段在生長與物質分配之間的調節。
3.4" 芽流的內涵與賴草種群的發展策略
芽流是對芽庫與構件之間在運轉過程中既有流通量又有流通率的定量描述模型[3,26]。芽流與構件結構的差異僅在于有無死亡組分,在流通率上,芽流是包括死亡在內芽周轉的全體構件組成譜,其內涵比結構譜更完整更豐富。譬如,芽死亡的內涵是營養繁殖失敗,芽周轉為分株和根莖的內涵是營養繁殖成功。在本研究賴草實驗同生群的芽庫中,在移植當年的營養繁殖成功率為40.6%。其中,轉為現實種群的分株占32.9%,轉為營養繁殖和養分貯藏的根莖占7.7%,而營養繁殖失敗率僅占0.1%;在第二年的營養繁殖成功率高達58.1%,其中分株占49.2%,根莖占8.9%,營養繁殖失敗率僅占0.9%。兩年的營養繁殖成功率中,分株的比率分別是根莖比率的4.3倍和5.5倍。可見,賴草營養繁殖的失敗率甚低,意味著親本的無效投資極少,而營養繁殖成功率之高是任何植物有性繁殖所無法比擬的。如果推而廣之到天然草地,營養繁殖成功率高還體現在芽苗的競爭力上。芽苗與親株相連接,在沒有長出綠色葉片或生產的有機質尚不足以滿足自身生理代謝需要之前,親株均供給所需物質,不僅保證了芽苗存活,也提高了與種子實生苗之間的競爭力,同時也起到了排他作用。這也是在天然草地一些典型的無性系植物經常形成大面積單一優勢種群落的植物領域性策略與原理[27]。對于賴草營養繁殖周轉中優先生產更多分株,與構件生長分析中分株冪律增長速率均為bgt;1相契合,意味著賴草無性系具有優先占據更多地上空間的種群發展策略。
4" 結論
在初始移植單生根莖芽苗的賴草實驗同生群中,當年和第二年所產生的全體營養繁殖構件數量特征均具有較大的可塑性,其變異系數均大于74%。在沒有種內和種間競爭的條件下,賴草無性系在年份間實際營養繁殖力的增長大于營養繁殖力的增長。賴草實驗同生群無性系大小及各構件組分的數量及構件總數在兩個年份生長階段和連續生長過程均為經典的指數增長模式。在芽流中,兩個年份賴草實驗同生群的營養繁殖失敗率均不足1%,營養繁殖成功率均較高,移植當年為40.6%,第二年為58.1%,其中,分株的比率分別是根莖比率的4.3倍和5.5倍。賴草實驗同生群內無性系實際營養繁殖力在最大值和最小值之間的可塑性變化是種群生存對策的體現,小的無性系具有優先占據更多地上空間的發展策略。
參考文獻
[1]" KLIMESˇOV J,NOBIS M P,HERBEN T. Senescence,ageing and death of the whole plant:morphological prerequisites and constraints of plant immortality[J]. New Phytologist,2015,206:14-18
[2]" NIKLAS K J,COBB E D. The evolutionary ecology (evo-eco) of plant asexual reproduction[J]. Evolutionary Ecology,2017,31(3):317-332
[3]" 楊允菲,魏春雁,張寶田,等.松嫩平原堿化草甸旱地生境蘆葦種群的芽流和芽庫動態[J].應用生態學報,2005,16(5):854-858
[4]" HARPER J L. Population biology of plants[M]. New York:Academic Press,1977:5-30
[5]" SILVERTOWN J W. Introduction to plant population ecology[M]. London and New York:Longman Press,1982:3-20
[6]" 董鳴.克隆植物生態學[M].北京:科學出版社,2011:80-81
[7]" 李建東,鄭慧瑩.松嫩平原鹽堿化草地治理及其生物生態機理[M].北京:科學出版社,1997:1-30
[8]" 盛軍,朱瑤,李海燕,等.長期刈割及圍欄封育方式對大油芒種群構件特征的影響[J].草地學報,2018,26(3):545-550
[9]" 張露丹,李海燕,楊允菲.松嫩平原單優種群落斑塊拂子茅種群構件的年齡結構[J].草業學報,2016,25(9):20-27
[10]宋月媛,楊允菲.松嫩平原林緣草地羽茅無性系構件結構與生長分析[J].草業學報,2019,28(7):168-174
[11]初麗爽,李海燕,楊允菲.松嫩平原異質生境羊草種群營養繁殖特征[J].應用生態學報,2020,31(1):83-88
[12]豐吉,劉志擴,李海燕,等.圍欄封育和長期刈割對松嫩草地羊草和野古草種群營養繁殖特征的影響[J].草業學報,2023,32(5):50-60
[13]趙凌平,王占彬,程積民.草地生態系統芽庫研究進展[J].草業學報,2015,24(7):172-179
[14]楊持.生態學[M].第3版.北京:高等教育出版社,2015:52-63
[15]GUO J,LI H Y,YANG Y F. Phenotypic plasticity in sexual reproduction based on nutrients supplied from vegetative ramets in a Leymus chinensis population[J]. Frontiers in Plant Science,2020,10:1681
[16]YUAN J H,LI H Y,YANG Y F. The compensatory tillering in the forage grass Hordeum brevisubulatum after simulated grazing of different severity[J]. Frontiers in Plant Science,2020,11:792
[17]楊允菲,宋向華.科爾沁草原區火電廠儲灰池植被恢復階段賴草種群構件的生物量年齡結構[J].草地學報,2005,13(2):130-133
[18]宋金枝,楊允菲.松嫩平原堿化草甸朝鮮堿茅無性系冬眠構件的結構及生長分析[J].生態學雜志,2006,25(7):743-746
[19]楊允菲,張寶田,李建東.松嫩平原人工草地野大麥無性系冬眠構件的結構及形成規律[J].生態學報,2004,24(2):268-273
[20]楊允菲,張寶田,張春華.松嫩平原賴草無性系構件的形成與空間擴展實驗[J].應用生態學報,2007,18(5):979-984
[21]WESTOBY M,WRIGHT I J. The leaf size-twig size spectrum and its relationship to other important spectra of variation among species[J]. Oecologia,2003,135:621-628
[22]WEST G B,BROWN J H,ENQUIST B J. A general model for the structure and allometry of plant vascular systems[J]. Nature,1999,400:664-667
[23]BROUAT C,MCKEY D. Leaf-stem allometry,hollow stems,and the evolution of caulinary domatia in myrmecophytes[J]. New Phytologist,2001,151:391-406
[24]SUN S C,JIN D M,SHI P L. The leaf size-twig size spectrum of temperate woody species along an altitudinal gradient:An invariant allometric scaling relationship[J]. Annals of Botany,2006,97:97-107
[25]ZHANG J,ZHOU C,YANG Y F. Growth of clonal modules on Agropyron michnoi in the Songnen Plain of Northeast China[J]. Phyton,International Journal of Experimental Botany,2015,84(2):417-422
[26]楊允菲,張寶田.松嫩平原貝加爾針茅無性系構件的結構及生長規律[J].應用生態學報,2005,16(12):2311-2315
[27]鄭慧瑩,李建東.松嫩平原的草地植被及其利用保護[M].北京:科學出版社,1993:45-108
(責任編輯" 劉婷婷)
猜你喜歡
今日農業(2021年11期)2021-08-13 08:53:34
小讀者(2021年2期)2021-03-29 05:03:48
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
中國生殖健康(2019年8期)2019-01-07 01:18:24
幸福(2018年33期)2018-12-05 05:22:46
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
作文周刊·小學一年級版(2017年17期)2017-06-27 19:32:55
廣東第二課堂·小學(2017年2期)2017-02-20 15:10:08
