基于微衛(wèi)星標(biāo)記的雪豹種群結(jié)構(gòu)及遺傳多樣性
2024-11-28 00:00:00周穎娜李金霖馬躍王震
野生動(dòng)物學(xué)報(bào) 2024年4期

摘 要 掌握雪豹(Panthera uncia)的遺傳多樣性信息對(duì)其保護(hù)和管理有重要意義。對(duì)104只雪豹個(gè)體的組織樣品提取DNA,結(jié)合文獻(xiàn)中提供的70個(gè)雪豹基因分型數(shù)據(jù),利用9個(gè)微衛(wèi)星多態(tài)性位點(diǎn)進(jìn)行遺傳多樣性研究。結(jié)果顯示:當(dāng)樣本量從70個(gè)增加到174個(gè)時(shí),雪豹種群有分化為2個(gè)種群的趨勢(shì),即西北部群和中部群;共檢測(cè)到45個(gè)等位基因(Na),中部、西北部2個(gè)種群平均每個(gè)位點(diǎn)Na分別為4. 556和4. 667,平均期望雜合度(He)分別為0. 574和0. 591,平均觀測(cè)雜合度(Ho)分別為0. 517和0. 498。雪豹群體的平均近交系數(shù)(FIS)為0. 137,平均遺傳分化系數(shù)(FST)為0. 067,平均多態(tài)信息含量(PIC)為0. 602,種群間遺傳變異為15. 24%,種群內(nèi)遺傳變異為84. 76%。9個(gè)微衛(wèi)星位點(diǎn)具有中高程度多態(tài)性,與其他雪豹研究相比,2個(gè)雪豹種群遺傳多樣性處于中等水平,種群間遺傳分化水平處于中等程度,遺傳變異大部分來源于群體內(nèi)部。研究結(jié)果對(duì)全球雪豹的種群結(jié)構(gòu)和遺傳多樣性研究具有重要意義,也為雪豹未來的科學(xué)保護(hù)提供可靠的遺傳背景。
關(guān)鍵詞:雪豹;種群結(jié)構(gòu);遺傳多樣性;微衛(wèi)星標(biāo)記
中圖分類號(hào):Q953
文獻(xiàn)標(biāo)志碼:A
文章編號(hào):2310 - 1490(2024)- 04 - 0699 - 10
DOI:10.12375/ysdwxb.20240402
雪豹(Panthera uncia)是全球廣泛關(guān)注的瀕危物種之一,隸屬于食肉目(Carnivora)貓科(Felidae)豹屬(Panthera)[1],通常出現(xiàn)在海拔3 000 ~ 4 500 m的區(qū)域,但在海拔900 ~ 1 500 m的戈壁沙漠的北部地區(qū)及海拔高至5 800 m的喜馬拉雅或青藏高原區(qū)域也有活動(dòng)[2]。長期以來,因棲息地喪失、獵物減少、氣候變化以及非法盜獵等因素的影響,雪豹種群持續(xù)下降[3?4],已被列為國家一級(jí)重點(diǎn)保護(hù)野生動(dòng)物和《中國生物多樣性紅色名錄:脊椎動(dòng)物卷》瀕危(EN)等級(jí)[5?6]。目前,雪豹分布于我國青藏高原及其周邊的12個(gè)國家,是山地生物多樣性的旗艦物種[7]。在中國,雪豹分布在8個(gè)省區(qū),其棲息地面積占全球雪豹棲息地面積的60%以上,中國在雪豹的物種保護(hù)方面有重要地位[1]。
了解并掌握雪豹的遺傳背景、致危因素及遺傳多樣性水平,對(duì)雪豹的科學(xué)保護(hù)具有重要意義。由于雪豹活動(dòng)的特殊性,樣品采集面臨著諸多挑戰(zhàn)。隨著技術(shù)的發(fā)展,非損傷性遺傳取樣法提供了解決方案,通過收集野生動(dòng)物脫落的毛發(fā)及排泄物等樣品,使用分子標(biāo)記如擴(kuò)增片段長度多態(tài)性[8]、隨機(jī)擴(kuò)增多態(tài)性DNA[9]、線粒體基因[10]及微衛(wèi)星[11]等,在這些分子標(biāo)記中,微衛(wèi)星因其高度多態(tài)性、選擇中性和共顯性等優(yōu)點(diǎn),在個(gè)體識(shí)別、遺傳多樣性評(píng)估和種群遺傳結(jié)構(gòu)分析等方面具有廣泛的應(yīng)用前景[12]。
目前,通過采集雪豹糞便獲得遺傳信息,并使用微衛(wèi)星分子標(biāo)記對(duì)雪豹群體遺傳多樣性進(jìn)行了較廣泛的研究,已經(jīng)覆蓋中國、蒙古國和印度等9 個(gè)國家[13?20]。有研究推測(cè)全球雪豹應(yīng)分為3個(gè)亞種,西部亞種(P. uncia uncia)分布于天山山脈、帕米爾高原、喀喇昆侖山脈和喜馬拉雅山脈西段等;中部亞種(P. u. uncioides)分布于青藏高原和喜馬拉雅山中東段;北部亞種(P. u. irbis)分布于阿爾泰山脈和蒙古國南部戈壁[16]。綜合以上研究,與豹屬其他物種相比,雪豹的遺傳多樣性處于較低水平[1],但雪豹的遺傳多樣性及亞種劃分可能因樣本采集位置及數(shù)量受限而導(dǎo)致結(jié)果差異。因此,本研究進(jìn)一步擴(kuò)大雪豹的采樣范圍及數(shù)量,以獲得更準(zhǔn)確的遺傳學(xué)數(shù)據(jù),為未來的雪豹種群劃分及遺傳學(xué)保護(hù)提供科學(xué)依據(jù)。
1 材料與方法
1. 1 試驗(yàn)樣本
以東北林業(yè)大學(xué)野生動(dòng)植物檢測(cè)中心實(shí)驗(yàn)室在執(zhí)法鑒定和行業(yè)監(jiān)管中采樣得到的104個(gè)雪豹個(gè)體組織樣本(101個(gè)來源于四川西部、西藏和青海等地,3個(gè)來自蒙古國進(jìn)境執(zhí)法檢查)為研究對(duì)象,將其置于密封袋中,-20 ℃保存。由文獻(xiàn)[16]獲得70個(gè)已知來源地的3個(gè)雪豹種群微衛(wèi)星基因分型數(shù)據(jù)。
1. 2 DNA 提取
使用AxyPrep Genomic DNA Miniprep Kit(Axy?gen,美國)試劑盒按說明書對(duì)104個(gè)雪豹組織樣本進(jìn)行DNA 提取。使用1. 0% 瓊脂糖凝膠電泳檢測(cè)DNA純度,用紫外分光光度計(jì)儀器(Implen,德國)測(cè)定DNA濃度,并置于-20 ℃保存?zhèn)溆谩?/p>
1. 3 PCR 擴(kuò)增
根據(jù)雪豹已有研究成果[16],選用9對(duì)微衛(wèi)星引物(PUN1157、PUN82、PUN100、PUN8972、PUN1283、PUN80、PUN1270、PUN272、PUN44)用于肌肉樣本DNA的擴(kuò)增(表1)。PCR反應(yīng)體系和反應(yīng)條件與文獻(xiàn)[16]一致。取3 μL PCR產(chǎn)物進(jìn)行1. 0%瓊脂糖凝膠檢測(cè),其中成功擴(kuò)增的PCR產(chǎn)物用ABI 3730XL測(cè)序儀(Applied Biosystems Inc,美國)進(jìn)行毛細(xì)管電泳檢測(cè),得到等位基因大小。以上實(shí)驗(yàn),除DNA提取外,均由北京六合華大基因科技有限公司完成。
1. 4 數(shù)據(jù)處理
利用9個(gè)微衛(wèi)星位點(diǎn)分別對(duì)70個(gè)文獻(xiàn)中的樣本和合并104個(gè)樣本后的全部樣本(174個(gè))進(jìn)行種群結(jié)構(gòu)分析。主成分分析(principal component analy?sis,PCA)使用R 3. 2. 4的adegenet 1. 2. 4包[21]。種群結(jié)構(gòu)分析使用Structure 2. 3. 4軟件[22]中的貝葉斯聚類方法,K 值1 ~ 8,每個(gè)K 值計(jì)算15次并確定最佳K值[23]。使用GenAlEx 6. 5[24]統(tǒng)計(jì)3個(gè)群體的等位基因數(shù)(Na)、有效等位基因(Ne)、觀測(cè)雜合度(Ho)、期望雜合度(He)、香農(nóng)信息指數(shù)(I)、近交系數(shù)(FIS)和群體遺傳分化系數(shù)(FST)。使用PowerMarker 3. 25[25]統(tǒng)計(jì)遺傳多態(tài)性信息含量(PIC)以及個(gè)體間的Nei’s遺傳距離。基于174個(gè)雪豹樣本間Nei’s遺傳距離,使用MEGA11 軟件[26]的個(gè)體鄰接法(neighborjoining,NJ)構(gòu)建聚類樹。使用Arlequin 3. 5. 2. 2 軟件[27]進(jìn)行分子方差分析(analysis of molecular vari?ance,AMOVA)。
2 結(jié)果與分析
2. 1 PCR 擴(kuò)增結(jié)果
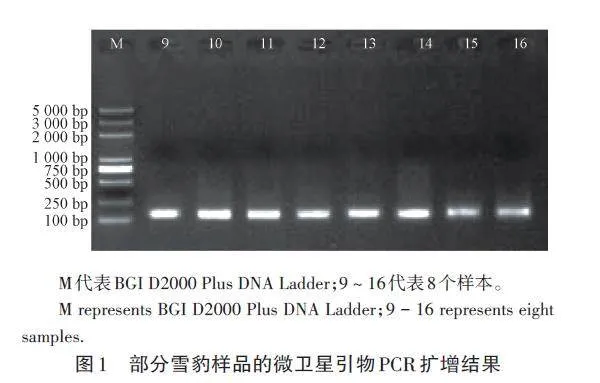
用9對(duì)微衛(wèi)星引物對(duì)104個(gè)雪豹樣本DNA進(jìn)行擴(kuò)增,獲得長度在100 ~ 150 bp的擴(kuò)增條帶,擴(kuò)增條帶清晰,大小一致(圖1),可以用于后續(xù)多態(tài)性分析。
2. 2 種群結(jié)構(gòu)
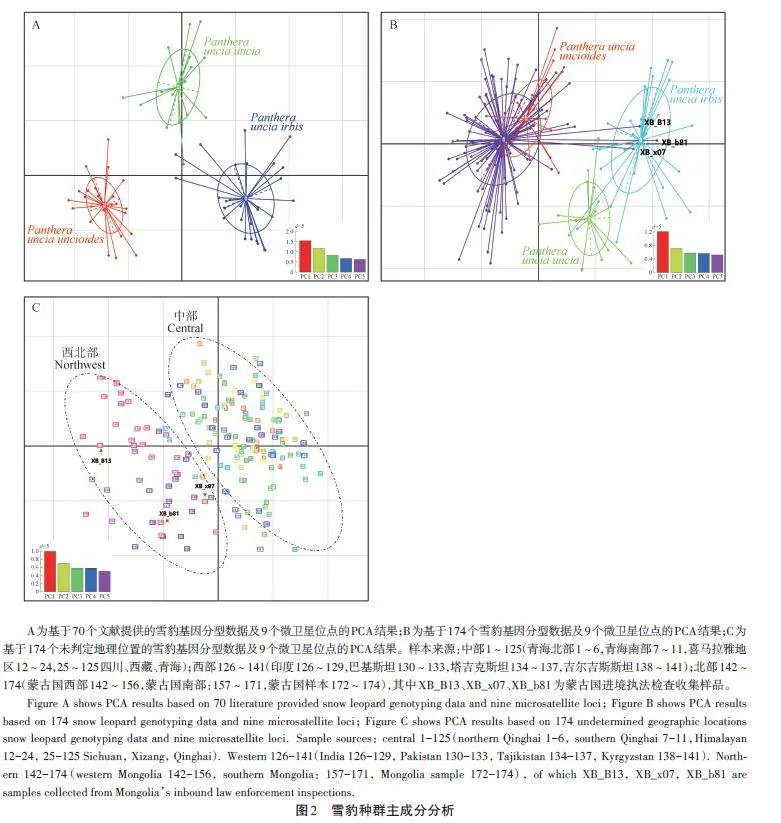
從圖2A可知,9個(gè)微衛(wèi)星位點(diǎn)可將70個(gè)已知樣本數(shù)據(jù)按照3個(gè)雪豹種群清晰分開,表明這些微衛(wèi)星位點(diǎn)具有較高的靈敏度。
全部樣本的PCA結(jié)果顯示:101個(gè)樣本與文獻(xiàn)[16]中的中部樣本聚集在一起,3個(gè)樣本與文獻(xiàn)[16]中的北部樣本聚集在一起,與樣本來源地相符(圖2B);同時(shí)相對(duì)于中部樣本,西部樣本與北部樣本呈現(xiàn)出聚集的趨勢(shì)。為排除地理標(biāo)簽對(duì)PCA分析的影響,去除地理標(biāo)簽限制后繪制的PCA結(jié)果顯示:整體數(shù)據(jù)主要分為2個(gè)聚類群(圖2C),即西部與北部種群數(shù)據(jù)并為一個(gè)群體,中部種群數(shù)據(jù)并為一個(gè)群體。
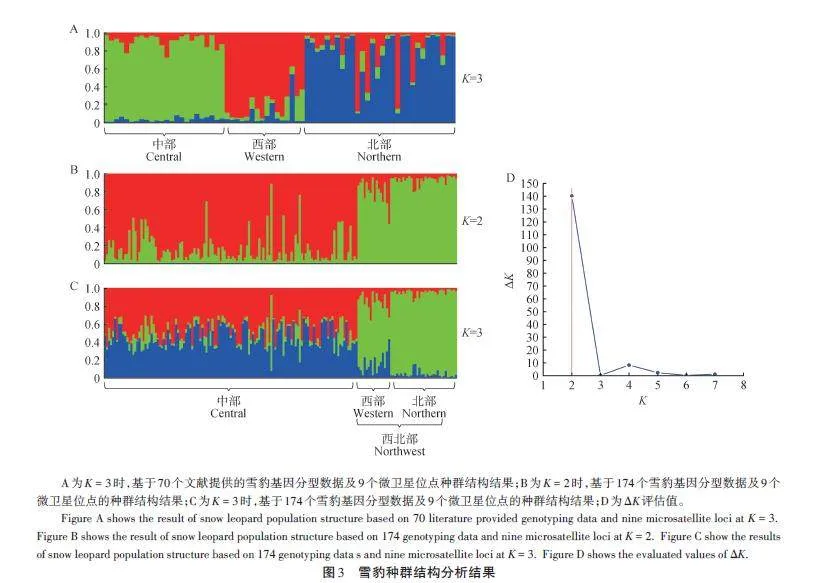
基于PCA 結(jié)果,對(duì)雪豹的種群結(jié)構(gòu)進(jìn)行分析。由圖3A可知,對(duì)于70個(gè)已知樣本,當(dāng)K = 3時(shí),中部、西部和北部種群祖先成分呈不同結(jié)構(gòu),與文獻(xiàn)[16]中的結(jié)果基本一致。174個(gè)樣本的分析結(jié)果顯示:當(dāng)K = 2和K = 3時(shí),兩種結(jié)果均體現(xiàn)出西部種群和北部種群祖先成分具有一致性(圖3B,C),且當(dāng)K = 2時(shí),ΔK 呈現(xiàn)最大值(圖3D)。結(jié)果表明,雪豹分為2個(gè)種群具有一定的合理性。
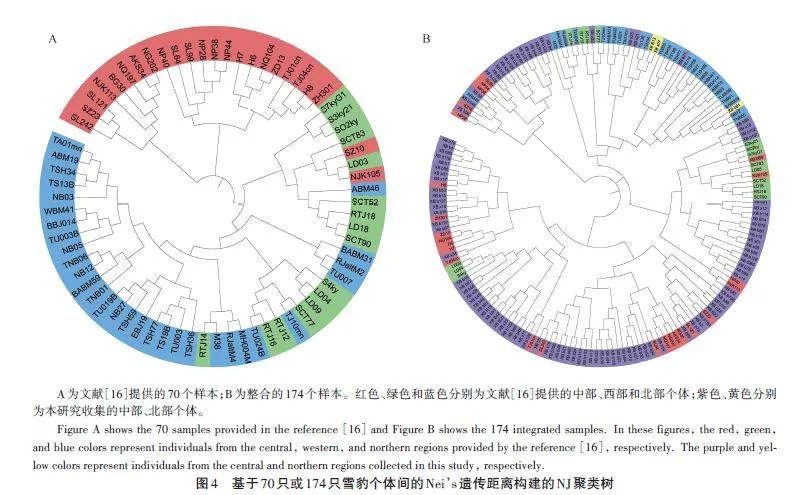
此外,基于70個(gè)樣本和174個(gè)樣本的Nei’s遺傳距離分別構(gòu)建了NJ聚類樹(圖4)。70個(gè)樣本的聚類樹顯示(圖4A),西部與北部個(gè)體混合在一支,中部個(gè)體主要分為兩支;174 個(gè)樣本的聚類樹顯示(圖4B),西部與北部大部分個(gè)體仍主要聚集在一支,不過一部分中部個(gè)體與西部個(gè)體混在一支。雖然兩個(gè)NJ聚類樹均存在少量個(gè)體的混合穿插,但均表明西部與北部個(gè)體的遺傳距離較近,也支持雪豹西部與北部種群并為一個(gè)群體。
2. 3 遺傳多樣性分析
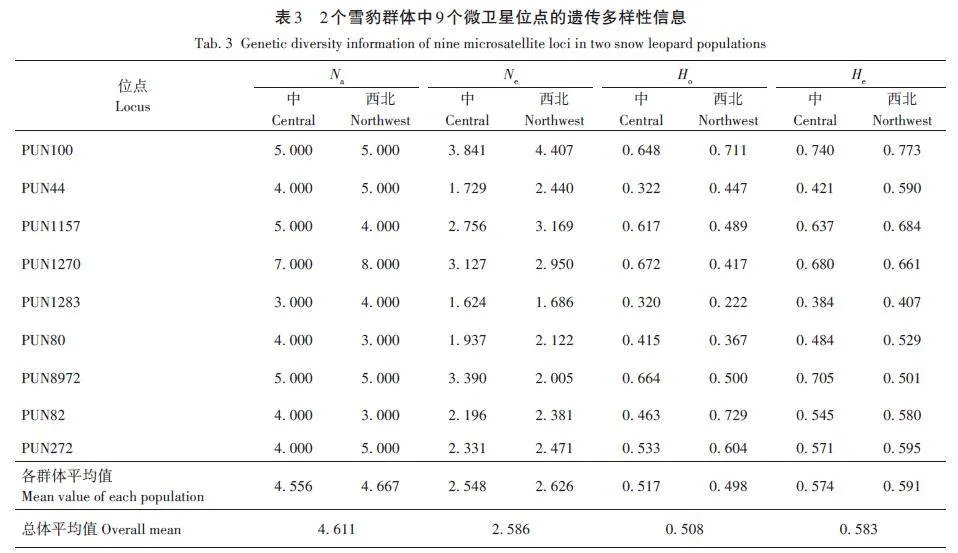
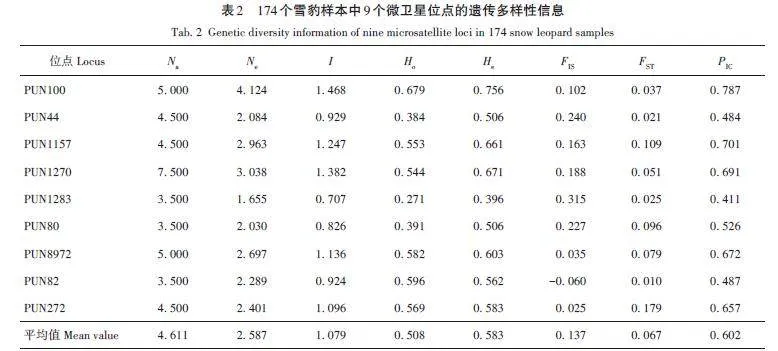
首先分析全部樣本的總體遺傳多樣性,共識(shí)別出45個(gè)Na。由表2可知,所有微衛(wèi)星位點(diǎn)的Na平均值為4. 611(3. 500 ~ 7. 500);Ne 平均值為2. 587(1. 655 ~4. 124);I 平均值為1. 079(0. 707 ~ 1. 468);Ho 平均值為0. 508(0. 271 ~ 0. 679);He 平均值為0. 583(0. 396 ~ 0. 756);FIS平均值為0. 137,其中PUN82為負(fù)值,其余8個(gè)位點(diǎn)為正值;FST平均值為0. 067(0. 010 ~0. 179);PIC平均值為0. 602(0. 411 ~ 0. 787)。結(jié)果表明,9個(gè)位點(diǎn)均呈現(xiàn)中高程度的遺傳多樣性,其中3個(gè)位點(diǎn)表現(xiàn)為中度多態(tài)性(0. 25 lt; PIC lt; 0. 50),分別為PUN44、PUN1283和PUN82,其余位點(diǎn)均為高度多態(tài)性位點(diǎn)(PIC gt; 0. 50),遺傳多樣性較豐富。
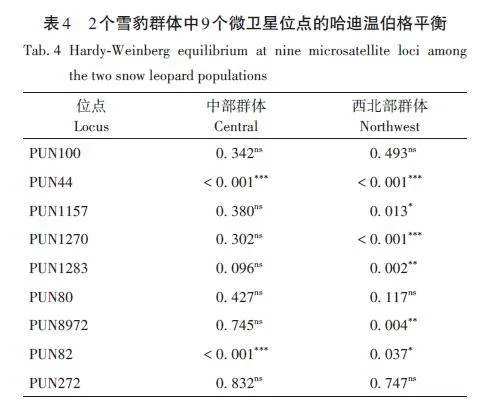
中部群和西北部群的遺傳多樣性結(jié)果顯示(表3):9 個(gè)位點(diǎn)在2 個(gè)種群中檢測(cè)到3 ~ 8 個(gè)Na,其中PUN1270位點(diǎn)檢測(cè)的Na數(shù)量最多,PUN1283、PUN80和PUN82位點(diǎn)檢測(cè)到的Na數(shù)量最少;中部和西北部2個(gè)種群的平均Na分別為4. 556和4. 667;平均Ne分別為2. 548和2. 626;平均Ho分別為0. 517和0. 498;平均He分別為0. 574和0. 591。哈迪溫伯格平衡檢驗(yàn)結(jié)果顯示:在中部種群中,PUN44和PUN82兩個(gè)位點(diǎn)不符合哈迪溫伯格平衡;西北部種群中PUN44、PUN1157、PUN1270、PUN1283、PUN8972 和PUN82六個(gè)位點(diǎn)偏離哈迪溫伯格平衡(表4)。
2. 4 微衛(wèi)星標(biāo)記的遺傳分化
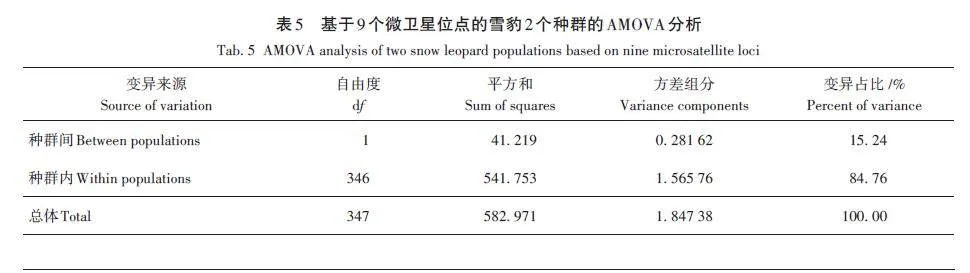
AMOVA結(jié)果顯示,在雪豹2個(gè)種群中,種群間的遺傳變異占15. 24%,種群內(nèi)的遺傳變異占84. 76%(表5),表明變異大部分發(fā)生在種群內(nèi)部,種群之間的遺傳變異所占比例較小,同時(shí)也表明2個(gè)雪豹種群間的基因交流較少,大部分的變異來源于群體內(nèi)部。
3 討論
3. 1 雪豹種群遺傳結(jié)構(gòu)
關(guān)于雪豹群體或亞種的劃分一直存在爭議。最早在2015年,Riordan et al.[28]根據(jù)模型分析雪豹種群的連通性,建議將雪豹分為南、北2個(gè)地理種群。在2017 年,Kitchener et al.[29]在對(duì)貓科動(dòng)物重新分類時(shí)沒有采納該意見,依然判斷全球雪豹為1個(gè)種群。同年,Janecka et al.[16]基于33個(gè)微衛(wèi)星位點(diǎn),對(duì)70個(gè)雪豹糞便樣本進(jìn)行基因分型,結(jié)合遺傳結(jié)構(gòu)和地理隔離,支持將全球雪豹分為3個(gè)亞種。本研究利用9個(gè)微衛(wèi)星位點(diǎn),先后對(duì)文獻(xiàn)[16]中70只雪豹個(gè)體數(shù)據(jù)和174只個(gè)體數(shù)據(jù)進(jìn)行分析,得到了不同的種群結(jié)構(gòu)。70只雪豹數(shù)據(jù)的種群結(jié)構(gòu)與文獻(xiàn)[16]結(jié)果相近(圖2A,圖3A),支持雪豹3個(gè)群體的劃分。但是,在174個(gè)樣本的結(jié)果中,西部群體和北部群體接近,形成一個(gè)群體,與中部種群相對(duì)應(yīng)(圖2B、C);種群結(jié)構(gòu)上也支持了2個(gè)群體的劃分(圖3B、C,圖4)。出現(xiàn)這樣的變化,可能歸因于Janecka et al.[16]研究的樣本量較小、地理覆蓋不全,進(jìn)而放大了小群體之間的差異。當(dāng)樣本量增大時(shí),西部群體和北部群體逐步聚集。實(shí)際上,Janecka et al.[16]的研究結(jié)果也支持這一觀點(diǎn),例如,在其小樣本分析中,雖然都存在少量混合,但北方群體的蒙古國西部群(15個(gè)樣本)和南部群(17個(gè)樣本)形成了2個(gè)不同的集群(文獻(xiàn)[16]圖2i);中部群體也分成了2個(gè)群體(文獻(xiàn)[16]圖2g),青海地區(qū)群(北部和南部,共11個(gè)樣本)與西藏地區(qū)群(含不丹和尼泊爾,共13 個(gè)樣本)。Senn et al.[30]也認(rèn)為Janecka et al. 的研究缺乏足夠的樣本量,不足以支持3個(gè)亞種的劃分。兩個(gè)NJ聚類樹也支持北部與西部群體并為一個(gè)群體,聚類樹上少量個(gè)體的混合穿插,可能與群體間的基因流動(dòng)有關(guān),也可能與微衛(wèi)星位點(diǎn)數(shù)量偏少、精度不夠有關(guān)。因此,本研究認(rèn)為目前的研究結(jié)果不支持Jan?ecka et al. 雪豹3個(gè)亞種的劃分,支持將雪豹按照2個(gè)群體劃分,即中部群和西北部群。在地理上,雪豹中部群體與北部、西部群體也存在一定的阻隔。雪豹中部群體,在北方有戈壁沙漠與北部群體相隔[1],南部有喜馬拉雅山脈與其南麓相隔,西部有塔克拉瑪干沙漠、塔里木盆地和南部的喀喇昆侖山脈等[31],也可能阻隔了其與西部群體的交流。不過,相對(duì)于中部樣本(125個(gè)),西部(16個(gè))和北部(33個(gè))的樣本數(shù)量仍較少,樣本數(shù)量不平衡可能影響了研究結(jié)果。
3. 2 雪豹種群遺傳多樣性
遺傳多樣性水平可以反映物種的進(jìn)化潛力[32],能為物種現(xiàn)狀評(píng)估和保護(hù)提供重要信息[33]。目前,使用微衛(wèi)星標(biāo)記的方法對(duì)雪豹種群進(jìn)行遺傳多樣性研究,樣本范圍覆蓋了9個(gè)國家,樣本數(shù)共達(dá)到211只,但單次研究中每個(gè)地區(qū)有效的樣本采集數(shù)量不超過30只[1],而且關(guān)于雪豹群體遺傳多樣性的不同研究結(jié)果出現(xiàn)了一定的波動(dòng)。
多態(tài)信息含量是表示微衛(wèi)星位點(diǎn)變異程度高低、衡量基因片段多態(tài)性的一個(gè)指標(biāo)[34],包括低度多態(tài)性位點(diǎn)(PIC lt; 0. 25)、中度多態(tài)性位點(diǎn)(0. 25 0. 50)[35]。本研究使用的9個(gè)微衛(wèi)星位點(diǎn)中,不存在低度多態(tài)性位點(diǎn),適用于雪豹遺傳學(xué)分析。全部雪豹群體在9個(gè)微衛(wèi)星位點(diǎn)上檢測(cè)到的Na 為3 ~ 8,Ne 為1. 655 ~4. 124,在其他雪豹研究中,Na為2. 571 ~ 6. 286[13?20],Ne為2. 031 ~ 2. 800[16,20],本研究結(jié)果與已有研究結(jié)果基本一致。本研究結(jié)果中Na 和Ne 的數(shù)值范圍更大,可能與樣本量大有關(guān)。此外,Na值均大于Ne,表明等位基因分布不均勻,存在差異,可能受到群體間潛在的近親繁殖或遺傳漂變等因素的影響,未來種群也存在等位基因丟失以及遺傳多樣性下降的風(fēng)險(xiǎn)。
雜合度是衡量群體遺傳多樣性程度最重要的指標(biāo)之一,數(shù)值越大代表種群間遺傳多樣性越豐富。此前關(guān)于雪豹的遺傳多樣性研究中,不同調(diào)查群體的Ho在0. 371 ~ 0. 887,He在0. 330 ~ 0. 781[13?20]。本結(jié)果中,中部和西北部2 個(gè)雪豹群體的Ho 分別為0. 517和0. 498,He分別為0. 574和0. 591,與此前的研究結(jié)果相比,處于中等水平,可能是樣本量增大之后得到了更接近于雪豹整體實(shí)際情況的結(jié)果。在這些研究中,周蕓蕓等[19]在青海2個(gè)縣以及甘肅阿克塞縣調(diào)查的雪豹遺傳多樣性偏高(平均Ho為0. 843,平均He為0. 759),可能歸因于樣本量小而產(chǎn)生的差異。通過雜合度數(shù)值,與豹屬其他物種[1]相比,雪豹的遺傳多樣性較低。
結(jié)果中所有位點(diǎn)的FIS 平均值為0. 137,遠(yuǎn)低于巴基斯坦地區(qū)雪豹的FIS 平均值(0. 38)[18]。可能歸因于在巴基斯坦進(jìn)行雪豹樣本采集的5個(gè)地區(qū)相距較近,而且該國雪豹分布范圍較小、數(shù)量較少[7],因此,可能加劇了群體內(nèi)部近親繁殖的概率。當(dāng)種群數(shù)量擴(kuò)大時(shí),近交系數(shù)下降,表明在更大的雪豹分布區(qū)中,雪豹的近交程度可能沒有達(dá)到更高的水平。相比于近交程度較低的大熊貓(Ailuropoda melano?leuca)(種群平均FIS為0. 081)[12,36]和近交程度很高的華南虎(Panthera tigris amoyensis)(0 lt; FIS lt; 0. 42)[12,37],雪豹種群近交程度偏低。
以上分析中,不同研究得到的雪豹遺傳多樣性指標(biāo)存在著一定的波動(dòng),這可能受到樣本數(shù)量及采樣范圍大小的影響,當(dāng)樣本量大、地理來源更廣時(shí),得到的結(jié)果可能更接近于實(shí)際情況。不過局部小樣本量的研究,對(duì)當(dāng)?shù)匮┍》N群的保護(hù)管理也具有一定的指導(dǎo)意義。
本研究利用174只雪豹個(gè)體9個(gè)位點(diǎn)的微衛(wèi)星數(shù)據(jù),對(duì)雪豹種群結(jié)構(gòu)和遺傳多樣性進(jìn)行分析,現(xiàn)有的結(jié)果支持將雪豹?jiǎng)澐譃?個(gè)群體,即中部群體和西北部群體。與其他雪豹有關(guān)研究相比,這2個(gè)種群的遺傳多樣性處于中等水平,近交水平也較低(低于Aruge et al.[18]的研究),略高于大熊貓。本研究通過大樣本微衛(wèi)星數(shù)據(jù)對(duì)全球雪豹的遺傳結(jié)構(gòu)進(jìn)行了分析,對(duì)雪豹的種群劃分和遺傳多樣性提供了更全面的數(shù)據(jù)支持,對(duì)雪豹的遺傳多樣性研究和指導(dǎo)保護(hù)實(shí)踐具有重要意義。
參考文獻(xiàn):
[1] 郝宇波, 金紅, 楊林,等. 雪豹保護(hù)遺傳學(xué)和基因組學(xué)研究進(jìn)展[J]. 獸類學(xué)報(bào), 2022, 42(5): 508-518.
HAO Y B, JIN H, YANG L, et al. Research advances in conser?vation genetics and genomics of snow leopard (Panthera uncia)[J]. Acta Theriologica Sinica, 2022, 42(5): 508-518.
[2] JACKSON R, MALLON D, MISHRA C, et al. Snow leopard sur?vival strategy: revised version 2014. 1 [R]. Seattle:Snow Leop?ard Network, 2014: 17.
[3] MAHESHWARI A, NIRAJ S K. Monitoring illegal trade in snowleopards: 2003-2014[J]. Global Ecology and Conservation,2018, 14: e00387.
[4] 洪洋,張晉東. 臥龍自然保護(hù)區(qū)雪豹的生境選擇偏好與食源結(jié)構(gòu)特征[J]. 野生動(dòng)物學(xué)報(bào),2021,42(2):295-305.
HONG Y, ZHANG J D. Habitat selection and food source of snowleopard (Panthera uncia) in Wolong National Nature Reserve, Si?chuan Province, China[J]. Chinese Journal of Wildlife,2021,42(2):295-305.
[5] 國家林業(yè)和草原局,農(nóng)業(yè)農(nóng)村部. 國家重點(diǎn)保護(hù)野生動(dòng)物名錄(2021 年2 月1 日修訂)[J]. 野生動(dòng)物學(xué)報(bào),2021,42(2):605-640.
National Forestry and Grassland Administration, Ministry of Agri?culture and Rural Affairs. List of national key protected wild ani?mals (revised on February 1, 2021) [J]. Chinese Journal ofWildlife, 2021, 42(2): 605-640.
[6] 生態(tài)環(huán)境部, 中國科學(xué)院. 中國生物多樣性紅色名錄: 脊椎動(dòng)物卷(2020)[EB/OL].(2023-05-18)[2024-04-10]. https://www. mee. gov. cn/xxgk2018/xxgk/xxgk01/202305/t20230522_1030745. html.
Ministry of Ecology and Environment, Chinese Academy ofSciences. China’s red list of biodiversity: vertebrate volume(2020)[EB/OL]. (2023-05-18)[2024-04-10]. https://www.mee. gov. cn/xxgk2018/xxgk/xxgk01/202305/t20230522_1030745.html.
[7] 楊子文,韓姝伊,李壹,等. 氣候變化對(duì)雪豹全球潛在適生區(qū)分布的影響與評(píng)估[J]. 生態(tài)學(xué)報(bào),2023,43(4):1412-1425.YANG Z W, HAN S Y, LI Y, et al. Impacts and assessment ofclimate change on the global distribution of potentially suitablehabitats for Panthera uncia[J]. Acta Ecologica Sinica, 2023,43(4):1412-1425.
[8] PFAU R S, KOZORA A N, GATICA-COLIMA A B, et al. Popu?lation genetic structure of a Chihuahuan Desert endemic mammal,the desert pocket gopher, Geomys arenarius[J]. Ecology and Evo?lution, 2023, 13(10): e10576.
[9] HUSSEIN M A A, EID M, RAHIMI M, et al. Comparative as?sessment of SSR and RAPD markers for genetic diversity in somemango cultivars[J]. PeerJ, 2023, 11: e15722.
[10] RAJ N, SUKUMARAN S, JOSE A, et al. Population geneticstructure of Randall’s threadfin bream Nemipterus randalli in In?dian waters based on mitochondrial and nuclear gene sequences[J]. Scientific Reports, 2024, 14(1): 7556.
[11] SHANG K, ZHANG L W, SONG J R, et al. Characterization ofthe microsatellite landscape provides insights into the evolution?ary dynamics of the mammals based on the chromosome-level ge?nomes[J]. Gene, 2023, 851: 146965.
[12] 魏輔文, 馬天笑, 胡義波. 中國瀕危獸類保護(hù)遺傳學(xué)研究進(jìn)展與展望[J]. 獸類學(xué)報(bào), 2021, 41(5): 571-580.
WEI F W, MA T X, HU Y B. Research advances and perspec?tives of conservation genetics of threatened mammals in China[J]. Acta Theriologica Sinica, 2021, 41(5): 571-581.
[13] JACKSON R, AHLBORN G. Snow leopards (Panthera uncia)in Nepal: home range and movements[J]. National GeographicResearch, 1989, 5(2): 161-175.
[14] JANECKA J E, JACKSON R, MUNKHTSOG B, et al. Charac?terization of 9 microsatellites and primers in snow leopards and aspecies-specific PCR assay for identifying noninvasive samples[J]. Conservation Genetics Resources, 2014, 6(2): 369-373.
[15] JANE?KA J E, JACKSON R, YUQUANG Z, et al. Populationmonitoring of snow leopards using noninvasive collection of scatsamples: a pilot study[J]. Animal Conservation, 2008, 11(5):401-411.
[16] JANECKA J E, ZHANG Y G, LI D Q, et al. Range-wide snowleopard phylogeography supports three subspecies[J]. Journal ofHeredity, 2017, 108(6): 597-607.
[17] ROZHNOV V V, ZVYCHAINAYA E Y, KUKSIN A N, et al.Noninvasive molecular genetic analysis in studying the ecology ofthe snow leopard: problems and prospects[J]. Russian Journalof Ecology, 2011, 42: 439-444.
[18] ARUGE S, BATOOL H, KHAN F M, et al. A pilot study:ge?netic diversity and population structure of snow leopards of Gilgit-Baltistan, Pakistan, using molecular techniques[J]. PeerJ,2019, 7: e7672.
[19] 周蕓蕓, 朵海瑞, 薛亞東, 等. 雪豹的微衛(wèi)星DNA遺傳多樣性[J]. 動(dòng)物學(xué)雜志, 2015, 50(2): 161-168.ZHOU Y Y, DUO H R, XUE Y D, et al. Genetic diversityanalysis of microsatellite DNA in snow leopard (Panthera uncia)[J]. Chinese Journal of Zoology, 2015, 50(2): 161-168.
[20] 張于光, HACKER C, 張宇,等. 三江源和祁連山國家公園雪豹種群的遺傳結(jié)構(gòu)分析[J]. 獸類學(xué)報(bào), 2019, 39(4):442-449.
ZHANG Y G,HACKER C, ZHANG Y, et al. An analysis of ge?netic structure of snow leopard populations in Sanjiangyuan andQilianshan National Parks [J]. Acta Theriologica Sinica, 2019,39( 4): 442-449.
[21] JOMBART T. adegenet: a R package for the multivariate analy?sis of genetic markers[J]. Bioinformatics, 2008, 24(11):1403-1405.
[22] PRITCHARD J K, STEPHENS M, DONNELLY P. Inference ofpopulation structure using multilocus genotype data[J]. Genet?ics, 2000, 155(2): 945-959.
[23] LI Y L, LIU J X. StructureSelector: a web?based software to se?lect and visualize the optimal number of clusters using multiple methods[J]. Molecular Ecology Resources, 2018, 18(1):176-177.
[24] PEAKALL R, SMOUSE P E. GenAlEx 6. 5: genetic analysis inExcel. Population genetic software for teaching and research: anupdate[J]. Bioinformatics, 2012, 28(19): 2537-2539.
[25] LIU K J, MUSE S V. PowerMarker: an integrated analysis envi?ronment for genetic marker analysis[J]. Bioinformatics, 2005,21(9): 2128-2129.
[26] TAMURA K, STECHER G, KUMAR S. MEGA11: molecularevolutionary genetics analysis version 11[J]. Molecular Biologyand Evolution, 2021, 38(7): 3022-3027.
[27] EXCOFFIER L, LISCHER H E L. Arlequin suite ver 3. 5: anew series of programs to perform population genetics analysesunder Linux and Windows[J]. Molecular Ecology Resources,2010, 10(3): 564-567.
[28] RIORDAN P, CUSHMAN S A, MALLON D, et al. Predictingglobal population connectivity and targeting conservation actionfor snow leopard across its range[J]. Ecography, 2016, 39(5):419-426.
[29] KITCHENER A C, BREITENMOSER-WüRSTEN C, EIZIRIKE, et al. A revised taxonomy of the Felidae: the final report ofthe cat classification task force of the IUCN/SSC cat specialistgroup[J]. Cat News, 2017(Special Issue 11):1-80.
[30] SENN H, MURRAY-DICKSON G, KITCHENER A C, et al.Response to Janecka et al. 2017[J]. Heredity, 2018, 120(6):581-585.
[31] LI J, MCCARTHY T M, WANG H, et al. Climate refugia ofsnow leopards in High Asia[J]. Biological Conservation, 2016,203: 188-196.
[32] FRANKEL O H, SOULé M E. Conservation and evolution[M].Cambridge: Cambridge University Press, 1981:358.
[33] HOGBIN P M, PEAKALL R. Evaluation of the contribution ofgenetic research to the management of the endangered plant Zie?ria prostrata[J]. Conservation Biology, 1999, 13(3): 514-522.
[34] 耿廣耀,由玉巖,劉群秀. 豚鹿SSR標(biāo)記開發(fā)與遺傳多樣性分析[J]. 野生動(dòng)物學(xué)報(bào),2022,43(3):816-820.
GENG G Y, YOU Y Y, LIU Q X. SSR molecular marker devel?opment and genetic diversity analysis of Axis porcinus[J]. Chi?nese Journal of Wildlife,2022,43(3):816-820.
[35] 方黎明,付春正,李慧,等. 浙江長興揚(yáng)子鱷種群遺傳多樣性研究[J]. 野生動(dòng)物學(xué)報(bào),2022,43(3):741-751.
FANG L M, FU C Z, LI H, et al. Research on population ge?netic diversity of Alligator sinensis in the Changxing City, Zheji?ang Province[J]. Chinese Journal of Wildlife,2022,43(3):741-751.
[36] SHAN L, HU Y B, ZHU L F, et al. Large-scale genetic surveyprovides insights into the captive management and reintroductionof giant pandas[J]. Molecular Biology and Evolution, 2014, 31(10): 2663-2671.
[37] XU Y C, FANG S G, LI Z K. Sustainability of the south Chinatiger: implications of inbreeding depression and introgression[J]. Conservation Genetics, 2007, 8(5): 1199-1207.
基金項(xiàng)目:國家林業(yè)和草原局機(jī)關(guān)業(yè)務(wù)委托項(xiàng)目(HFW220200050)
