茶葉成分EGCG與L-theanine聯合應用的神經保護作用研究
2024-11-08 00:00:00丁樹洽謝昕雅劉助生廖賢軍劉仲華蔡淑嫻
茶葉科學 2024年5期
摘要:分化的神經細胞需要在細胞靜息狀態下維護軸突的生長和功能。前期研究顯示,表沒食子兒茶素沒食子酸酯(Epigallocatechin gallate,EGCG)和L-茶氨酸(L-theanine)能夠維持神經細胞的靜息狀態并具有神經修復作用,但具體的作用機制還不清楚。在Aβ25-35誘導的PC12細胞損傷模型中,EGCG和L-theanine聯合處理改善細胞代謝和修復能力,提高了細胞活力,表現顯著的協同作用。轉錄組和網絡藥理學分析結果表明,EGCG主要通過抑制氧化應激、調節脂肪酸代謝和淀粉樣蛋白毒性應激來維持細胞靜息態;L-theanine則通過促進軸突生長、調節神經代謝和突觸功能發揮作用。兩者聯合應用對細胞網絡的調節更為廣泛和溫和,減少對細胞的刺激作用。本研究為茶葉的神經保護作用及其在老齡化社會中的飲用價值提供了理論依據。
關鍵詞:表沒食子兒茶素沒食子酸酯;L-茶氨酸;神經保護;網絡藥理;轉錄組;聯合分析
中圖分類號:S571.1;R972+.6 文獻標識碼:A 文章編號:1000-369X(2024)05-779-14
A Study on the Neuroprotective Effects of Combined EGCG and L-Theanine from Tea Leaves
DING Shuqia1,3, XIE Xinya1,3, LIU Zhusheng2*, LIAO Xianjun2, LIU Zhonghua1,3, CAI Shuxian1,3*
1. National Research Center of Engineering and Technology for Utilization of Botanical Functional Ingredients, Changsha 410128, China; 2. Guangxi Research Institute of Tea Science, Guilin 541004, China; 3. Key Lab of Education Ministry of Hunan Agricultural University for Tea Science, Changsha 410128, China
Abstracts: Differentiated neurons need to maintain axonal growth and function in a quiescent state. Previous studies have shown that epigallocatechin gallate (EGCG) and L-theanine can maintain the quiescent state of neurons and have neurorestorative effects, although the specific mechanisms are still unclear. In the Aβ25-35-induced PC12 cell damage model, combined treatment with EGCG and L-theanine improved cell metabolism and repair capacity, enhanced cell viability and showed a significant synergistic effect. Transcriptomic and network pharmacological analyses indicate that EGCG mainly maintains the quiescent state of cells by inhibiting oxidative stress, regulating fatty acid metabolism, and mitigating amyloid protein toxicity stress. L-theanine promotes axonal growth and regulates neuronal metabolism and synaptic function. The combined application of both compounds results in a broader and milder regulation of cellular networks, reducing cellular stress. This study provided theoretical support for the neuroprotective effects of tea and its value in an aging society.
Keywords: EGCG, L-theanine, neuroprotection, network pharmacology, transcriptome, joint analysis
淀粉樣蛋白病(Amyloidosis)是由內源性蛋白的異常聚集所形成的淀粉樣蛋白沉積于組織中,引發細胞毒性,最終導致組織損傷和病變[1-2]。目前有超過36種淀粉樣蛋白可沉積于心臟、腎臟、神經系統、腦組織、眼球、皮膚、血管等組織,且具有特異性[3]。研究發現,除了致病蛋白Aβ42、IAPP、α-突觸核蛋白等外,所有的蛋白質、核酸和脂肪等生物大分子均有集聚化傾向,可引發和促進大部分與衰老相關的疾病,如動脈粥樣硬化、腫瘤、2型糖尿病和阿爾茨海默病(Alzheimer's disease,AD)等。致病蛋白質聚集體的毒性可能源自其固有的錯誤折疊性質和結構異質性。這些特性導致大量異常相互作用,引起細胞損傷與死亡[4-5]。Aβ在高濃度時形成沉淀斑塊,無分解代謝途徑,因此抑制Aβ結構的形成對于預防老年性疾病至關重要。
AD及其他神經退行性疾病(Neurodegene-
rative diseases,NDs)的主要病理機制包括細胞外Aβ斑塊的形成、異常磷酸化的tau蛋白在細胞內積累、神經元突觸功能障礙以及神經元損失[6-8]。現有療法主要緩解癥狀,無法阻止疾病進程,并可能有副作用[9]。因此,開發能作用于多個生物靶點的天然產品具有重要治療潛力。
有研究認為,日常飲用適量的綠茶可顯著降低認知障礙的風險[10]。綠茶中的兒茶素,尤其是EGCG,具有多重健康益處,可通過抗炎、抗氧化等途徑對ND有防治作用[11]。EGCG能阻斷淀粉樣蛋白的聚集并分解原纖維,改善中樞記憶缺陷[12-14]。口服EGCG后,其代謝物EGC和GA促進神經軸突生長,減少腦部氧化損傷[15-16]。網絡藥理學研究已分析了EGCG及其代謝物對疾病靶點的影響[17-18]。L-茶氨酸(L-theanine)作為安全添加劑,在腦部、胃部和肝臟的炎癥損傷中顯示出其抗凋亡和保護作用[19-20]。口服L-theanine后大鼠肝臟中的乙胺和谷氨酸水平顯著提高[21],這種變化與其直接代謝和谷氨酰胺酶的作用有關[22]。L-
theanine通過抑制氧化損傷和tau蛋白過度磷酸化減輕神經毒性,降低鎘誘導的神經元死亡[23]。
本研究采用細胞試驗、轉錄組分析和網絡藥理學方法,進一步探索了EGCG和L-theanine及其主要代謝物在神經保護方面的協同效應和作用機制,通過轉錄組和網絡藥理的交集靶點分析,為揭示茶葉的神經保護機制提供重要的理論依據。
1 材料與方法
1.1 試驗材料與試劑
表沒食子兒茶素沒食子酸酯(Epigallocat-
echin Gallate,EGCG)和L-theanine(純度均為99%以上)均購自Sigma-Aldrich公司(美國密蘇里州圣路易斯市)。大鼠嗜鉻細胞瘤細胞系(PC12細胞)由北京協和醫學院細胞庫(北京)提供。Dulbecco改良Eagle培養基(DMEM)、胎牛血清(FBS)和胰蛋白酶均由美國Biological Industries提供。β淀粉樣蛋白25-35(Aβ25-35)購自美國Sigma公司。氟硼二吡咯類熒光染料(BODIPY)購自美國Invitrogen公司。
1.2 不同Aβ25-35蛋白樣品制備
將EGCG、L-theanine及其組合與Aβ25-35按等濃度混合,并在37 ℃的培養箱中孵育。孵育后,對混合液進行稀釋并加入至細胞中,以探索最佳協同濃度。在后續結果描述中,這些處理組分別標記為Aβ25-35、Aβ25-35/EGCG、Aβ25-35/L-theanine及Aβ25-35/EGCG+L-theanine。
1.3 細胞培養
PC12細胞在含10%胎牛血清、100 U·mL-1青霉素和100 μg·mL-1鏈霉素的Dulbecco改良Eagle培養基中培養,置于含5% CO2的37 ℃恒溫培養箱內。
1.4 細胞活力檢測
將PC12細胞接種于96孔板中,密度為每孔1×104個,培養24 h。然后用不同的Aβ25-35蛋白樣品(Aβ25-35、Aβ25-35/EGCG、Aβ25-35/L-
theanine和Aβ25-35/EGCG+L-theanine)處理細胞24 h,對照組加入等量的無菌水。去除上清液后,加入含0.5 g·mL-1 MTT的培養基,孵育4 h。隨后除去上清液,每孔加入150 μL DMSO。在多功能酶標儀上,以570 nm波長測量吸光度。細胞活力以處理組與對照組吸光度的平均百分比表示,對照組活力設定為100%。
1.5 熒光染色分析
將細胞以每孔5×104個的密度接種到含有蓋玻片的24孔細胞培養板中,并按1.3章節所述的方法培養。培養24 h后,用PBS在避光條件下洗滌細胞3次。然后,將蓋玻片倒置于含有DAPI的載玻片上封片,使用Zeiss公司的Axio Scope.A1熒光顯微鏡進行觀察和拍攝。
1.6 RNA測序和基因表達分析
試驗結束后,立即將收集的細胞樣品在液氮中預凍2 min,隨后在干冰上快速冷凍并送至深圳華大基因股份有限公司進行轉錄組測序(RNA-seq)。對每個基因的表達水平進行標準化處理后,使用每千堿基轉錄物的片段數(Fragments per kilobase of transcript per million mapped reads,FPKM)來計算基因表達水平。為了在轉錄組水平上全面探索作用機制,以|Fold Change|≥1.2和Q值≤0.05為條件,篩選差異表達基因(Differentially expressed genes,DEGs)進行GO生物過程(Biological process,BP)富集分析。
1.7 EGCG和L-theanine及其主要代謝物藥理學信息收集
采用Lipinski的五法則來評估所設計化合物的體內吸收潛力[24],包括分子量(MW)<500,氫鍵供體數量(Hdon)≤5,氫鍵受體數量(Hacc)≤10,脂水分配系數(LogP)≤5,以及可旋轉鍵數(Rbon)≤10。使用SwissADME網絡工具(www.swissadme.ch)評估化合物的藥物代謝動力學(Absorption,distribution,metabolism,and excretion,ADME)[25],同時測定拓撲極性表面積(Topological polar surface area,TPSA)、脂水分布系數(LogS)和皮膚滲透率(LogKp)。
1.8 EGCG和L-theanine及其主要代謝物相關靶標的收集
將主要代謝物的化學結構導入SwissTar-
getPrediction(www.swisstargetprediction.ch)和TCMSP(https://old.tcmsp-e.com/tcmsp.php)數據庫,以獲取各自的作用靶點。
1.9 篩選神經退行性疾病(NDs)靶標
使用GeneCards數據庫(www.genecards.
org)[26],以“神經退行性疾病”作為關鍵詞,篩選與NDs相關的疾病靶標。使用Venny 2.1.0軟件評估EGCG和L-theanine及其主要代謝物相關靶點與NDs相關靶點的交集,從而定義抗NDs相關靶點。在文中,EGCG組、L-theanine組和EGCG+L-theanine組的抗NDs相關靶點分別標記為EGCG/ND、L-theanine/ND和EGCG+L-theanine/ND。
1.10 蛋白質-蛋白質相互作用(PPI)網絡與聚類分析
使用STRING數據庫(https://string-db.org)構建PPI網絡,選定的生物體為智人(Homo sapiens)。PPI網絡由代表目標蛋白的節點和表示蛋白質-蛋白質相互作用的邊組成。使用Cytoscape軟件(v.3.8.2)確定核心目標,其插件MCODE用于分析網絡中的聚類模塊[27]。
1.11 轉錄組學與網絡藥理學聯合分析
使用Draw Venn Diagram在線程序,由網絡藥理學篩選出的EGCG和L-theanine協同治療神經退行性疾病的潛在靶點,以及轉錄組學數據中協同組與模型組相比較的差異表達mRNA基因取交集,此分析幫助識別了兩種方法共同支持的關鍵靶基因,其中弦圖和熱圖分析由微生信(www.bioinformatics.com.cn)在線平臺完成。
1.12 數據處理方法
所有數據均以平均值±標準差(Mean±SD)表示,取自3次重復的獨立試驗。不同組間的統計比較采用單因素方差分析(ANOVA)和多重對比后檢驗。
2 結果與分析
2.1 不同處理組的PC12細胞比較分析
2.1.1 EGCG與Aβ25-35/EGCG處理的PC12細胞比較分析
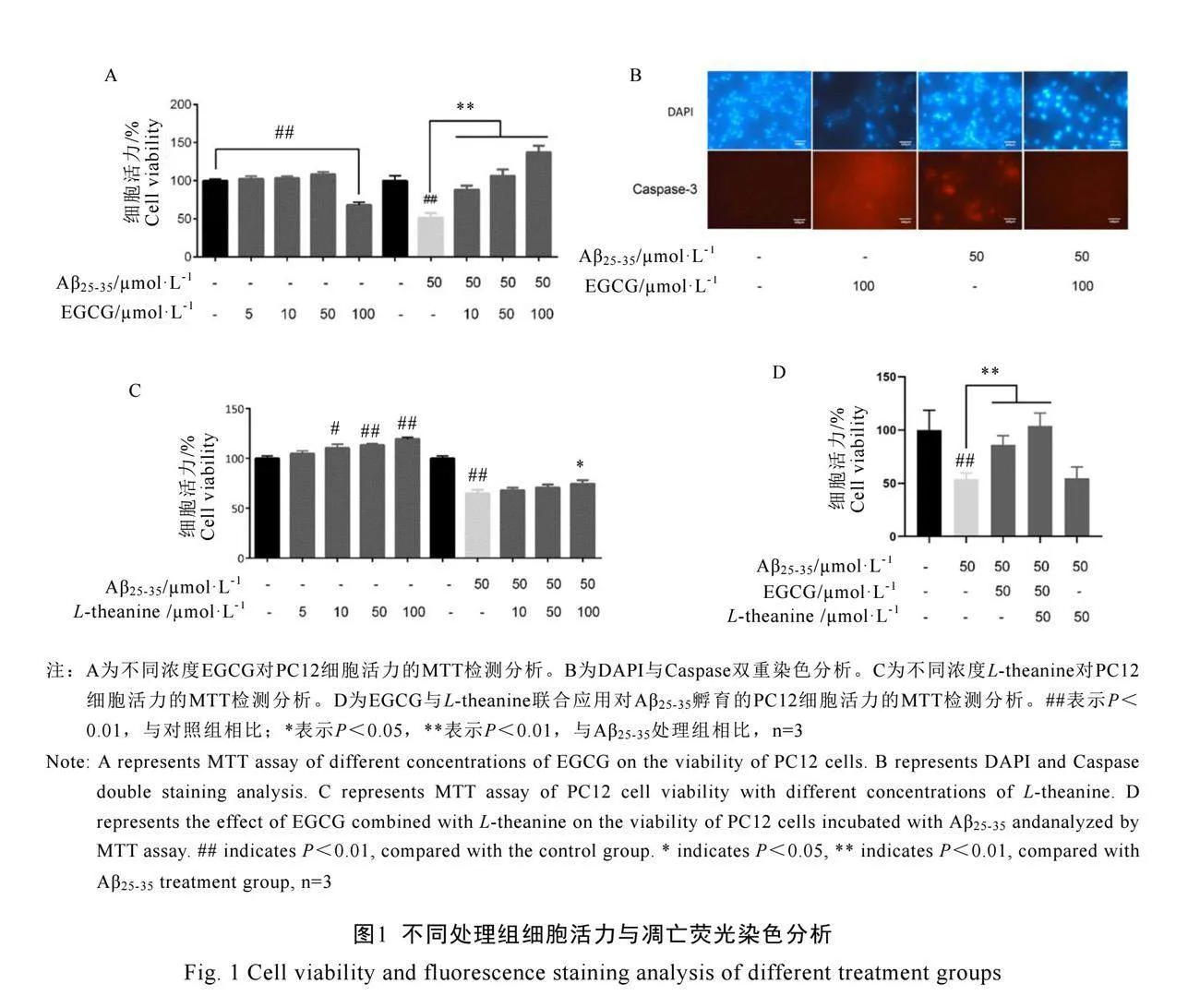
不同濃度EGCG分別孵育PC12細胞24 h,結果顯示在50 μmol·L-1濃度范圍內,EGCG對細胞活力影響不顯著;而在100 μmol·L-1濃度時,EGCG顯著降低細胞活力(圖1A)。相比之下,Aβ25-35/EGCG孵育PC12細胞24 h后,與Control組相比,50 μmol·L-1 Aβ25-35細胞活力下降約50%。隨著EGCG濃度增加,Aβ25-35/EGCG處理組的細胞活力顯著增加,尤其在EGCG濃度為100 μmol·L-1時,細胞活力超過Control組,表明Aβ25-35/EGCG處理組在EGCG高濃度時具有促進細胞生長的作用(圖1A)。
細胞凋亡熒光染色結果表明,EGCG單獨孵育PC12細胞24 h,與Control組相比,100 μmol·L-1 EGCG處理的細胞核結構不完整,caspase-3染色強烈,表現出促凋亡活性(圖1B)。50 μmol·L-1 Aβ25-35處理組的DAPI染色顯示細胞異染色質集聚,caspase-3染色增強,細胞核集聚,早期凋亡增加;而50 μmol·L-1 Aβ25-35/100 μmol·L-1 EGCG處理組細胞核增大,caspase-3幾乎沒有陽性染色,細胞未表現凋亡性,這與100 μmol·L-1 EGCG單獨處理的促凋亡作用相反(圖1B)。
2.1.2 L-theanine與Aβ25-35/L-theanine處理的PC12細胞比較分析
MTT試驗結果表明,不同濃度的L-theanine孵育PC12細胞,L-theanine濃度低于100 μmol·L-1時,細胞活力隨其濃度增加而提高(圖1C)。PC12細胞用50 μmol·L-1 Aβ25-35孵育處理7 d,再分別采用不同濃度的L-theanine處理后發現,與未加L-theanine相比,加L-theanine濃度低于100 μmol·L-1時對Aβ25-35沒有表現顯著的保護作用(圖1C)。
2.1.3 EGCG與L-theanine的協同作用分析
通過MTT檢測可得,模型組顯著抑制細胞活力(P<0.05),在添加了EGCG和L-theanine之后,細胞的活力得到了顯著提高,其中L-theanine的效果更為顯著(圖1D)。當同時添加這兩種藥物時,細胞活力達到了最高水平。因此,我們推測EGCG和L-theanine可能具有協同作用,在同時使用時可以顯著提高細胞活力。
2.2 轉錄組測序結果與差異基因分析
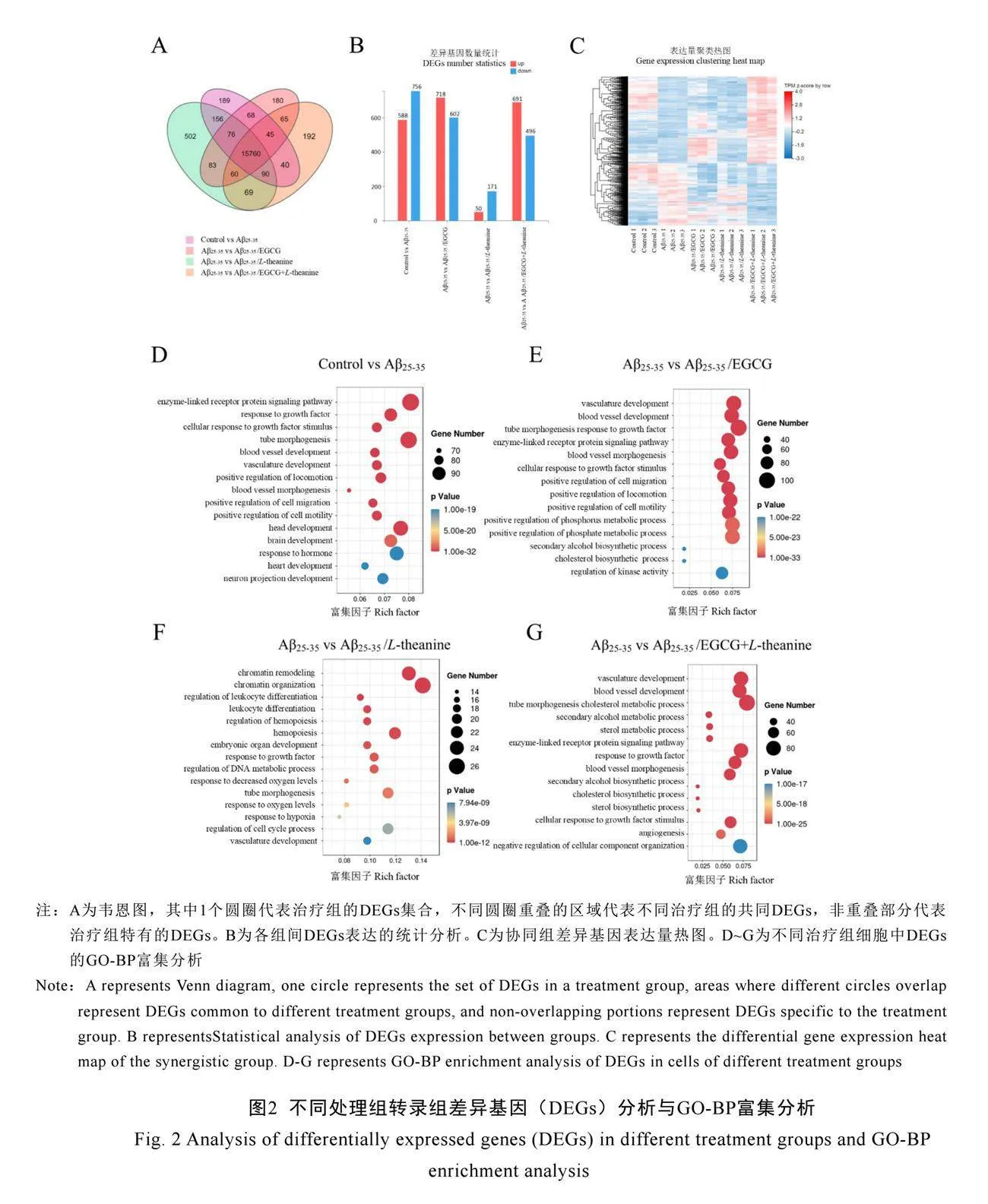
基于圖1D的細胞協同試驗結果,進行了轉錄組測序分析。從15個轉錄組cDNA文庫中獲得了69 987萬個原始讀數,過濾后有效讀數占比超過91.23%。將這些讀數與小鼠基因組進行比對,總比對率超過81.7%,唯一比對率超過76.65%,表明測序質量高,適用于后續分析。韋恩圖顯示,所有處理組中共有15 760個共表達基因(圖2A)。根據|Fold Change|≥1.2和Q值≤0.05篩選差異表達基因(DEGs),與對照組相比,Aβ25-35建模組鑒定出1 344個DEGs(上調588個,下調756個);Aβ25-35/EGCG組鑒定出1 320個DEGs(上調718個,下調602個);Aβ25-35/L-theanine組鑒定出221個DEGs(上調50個,下調171個);Aβ25-35/EGCG+L-theanine組鑒定出1 187個DEGs(上調691個,下調496個)(圖2B)。結果表明,與Aβ25-35相比,Aβ25-35/EGCG對細胞轉錄組的影響最為顯著,約是Aβ25-35/L-theanine的6倍。
圖2D至2G展示了不同組別差異基因的KEGG富集分析結果,揭示了Aβ25-35處理、EGCG處理、L-theanine處理及其聯合處理對細胞的多方面影響。Aβ25-35處理顯著干擾了血管發育、染色質重塑、細胞對生長因子反應和細胞遷移等多個關鍵生物過程,表明Aβ25-35通過影響細胞信號傳導、代謝調節、發育和免疫應答,可能導致神經退行性改變和組織損傷(圖2D)。EGCG處理顯著影響了酶聯受體蛋白信號通路、磷酸代謝和膽固醇生物合成等關鍵通路,通過促進細胞增殖和修復、調節代謝平衡,
提高組織修復能力和抗逆性(圖2E)。L-theanine處理調控了染色質重塑、DNA代謝和免疫過程,顯著影響了基因表達、細胞周期和低氧反應,表明其在基因調控和免疫增強方面具有重要作用(圖2F)。EGCG和L-theanine聯合處理擴展了單獨處理的作用范圍,顯著調控了血管發育、膽固醇和次級醇代謝、氧化還原過程等通路,展現出顯著的協同效應,可能通過多機制、多途徑的聯合作用,增強神經保護,改善細胞代謝和修復能力,顯示出在神經退行性疾病防治中的潛在應用價值(圖2G)。
KEGG分析結果表明,P值越低,通路越顯著,富集因子反映了通路中差異基因的數量。與EGCG相比,L-theanine對細胞轉錄組的影響相對較弱,但聯合處理展示了更強的調控能力,為神經保護機制及臨床應用研究提供了重要依據。
2.3 EGCG和L-theanine及其主要代謝物的網絡藥理學分析
2.3.1 EGCG和L-theanine及其主要代謝物ADME性質
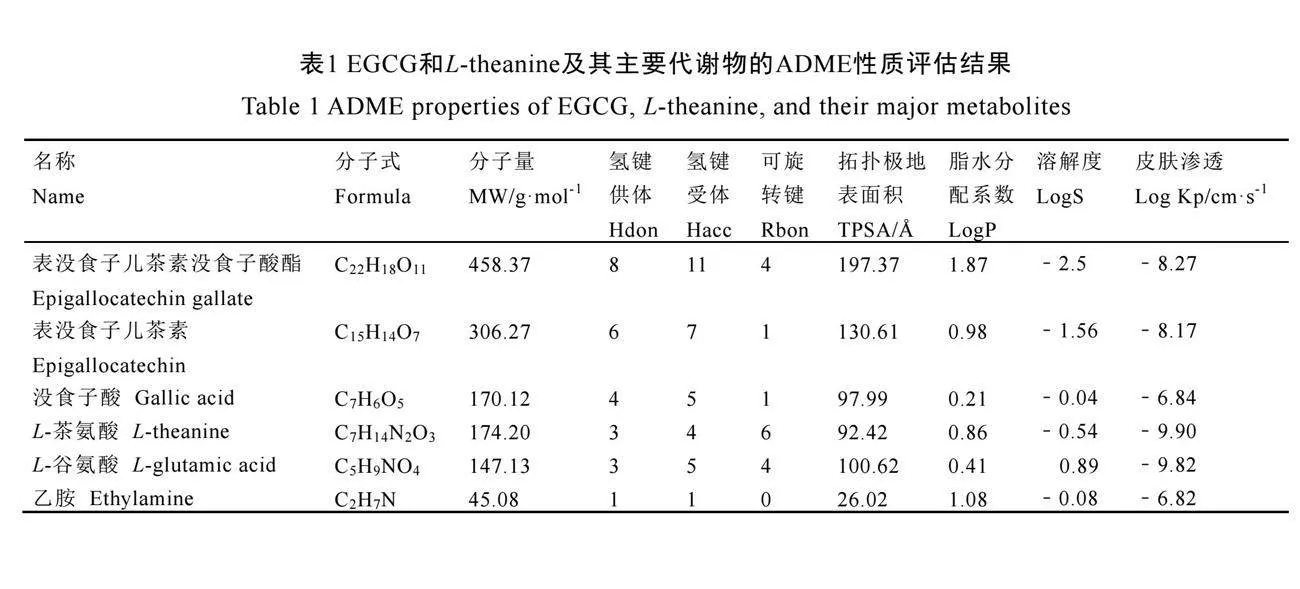
EGCG和L-theanine及其主要代謝物的化學結構從PubChem數據庫中獲得(圖3)。隨后,使用在線工具SwissADME對這些代謝產物的吸收、分布、代謝和排泄(ADME)性質進行了深入評估(表1)。分析結果顯示,拓撲極地表面積(Topological polar surface area,TPSA)和膜滲透性具有良好的相關性。一般來說,較大的TPSA值可能會導致較低的膜滲
透性。脂溶性是藥物化學中小分子的重要參數,脂水分配系數(logP)大于零時化合物親脂性比較強,反之則表明親水性較強。溶解度(LogS)則決定了EGCG等物質在腸道中的吸收與口服利用度。表1的數據表明,EGCG和L-theanine的代謝物在跨細胞膜滲透性方面表現良好。這些特性有助于它們在體內的有效吸收和利用,支持其在神經保護中的潛在應用。
2.3.2 EGCG與L-theanine及其主要代謝物抗NDs靶標篩選和PPI分析
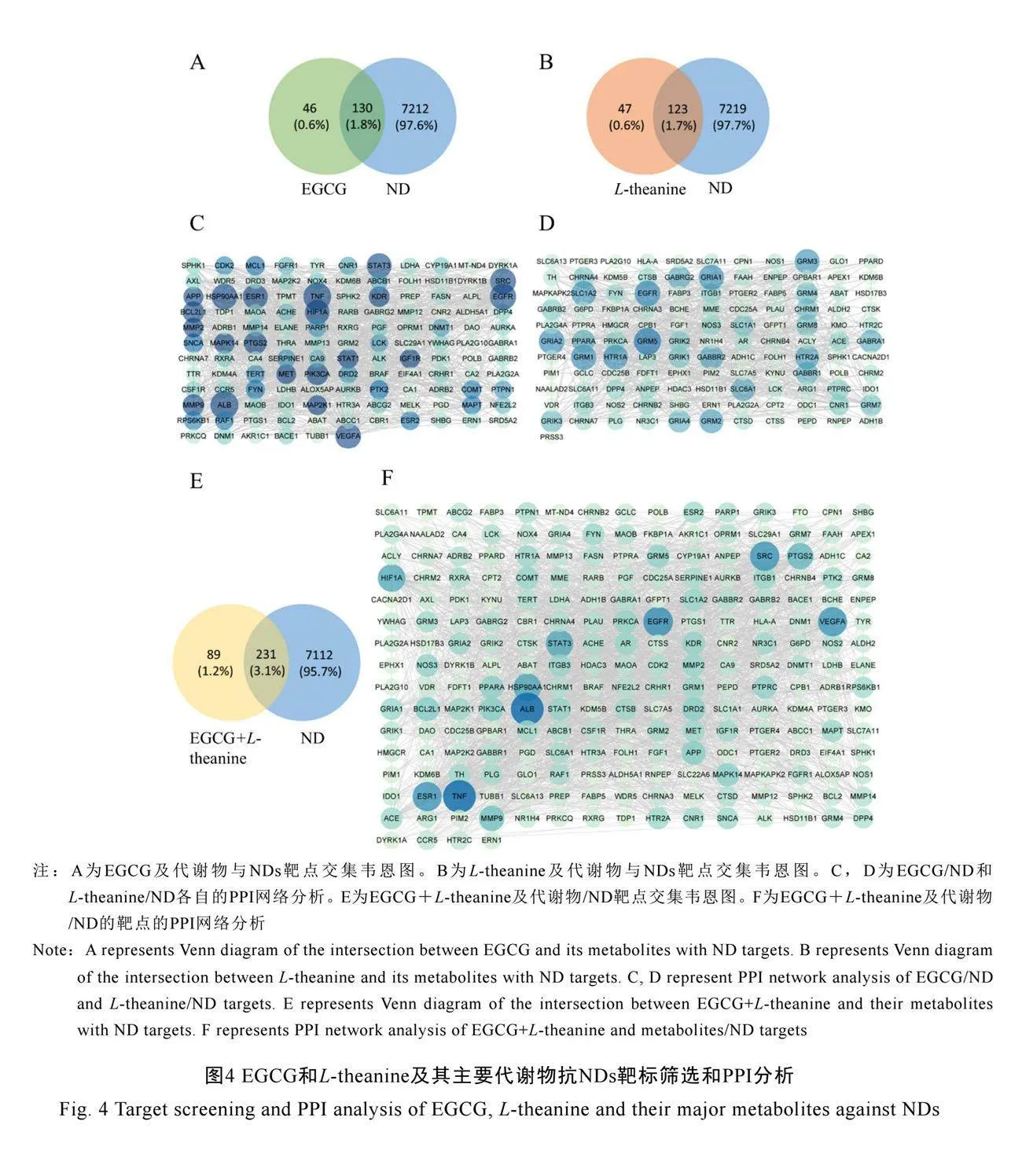
通過SwissTargetPrediction數據庫對EGCG及其主要代謝物EGC和GA的潛在靶點進行了預測,共獲得了176個靶點。對于L-theanine及其主要代謝物谷氨酸和乙胺的潛在靶點進行預測,共獲得了170個靶點。EGCG與L-theanine及其主要代謝物的共有潛在靶點320個。同時,從GeneCards數據庫中確定了7 341個與NDs相關的靶點。使用維恩圖展現EGCG和L-theanine主要代謝物與NDs相關的交集靶點(圖4A、圖4B與圖4E)。
為了探究交集靶點之間的關系,使用STRING數據庫進行了蛋白質-蛋白質相互作用(Protein-protein interaction,PPI)分析。EGCG/ND的靶點生成了包含127個節點和
1 010條邊的PPI網絡,平均節點度值為15.5(圖4C)。L-theanine/ND的靶點生成了包含122個節點和602條邊的PPI網絡,平均節點度值為9.79(圖4D)。EGCG+L-theanine/ND靶點生成了229個節點和2 372條邊的PPI網絡(圖4F)。靶點之間連線越多,表示該靶點與其他靶點的關系越密切,度值越大,被確定為核心靶點。核心靶點可能在NDs治療中發揮重要作
用。EGCG和L-theanine共有潛在靶點之間的連接更為緊密。
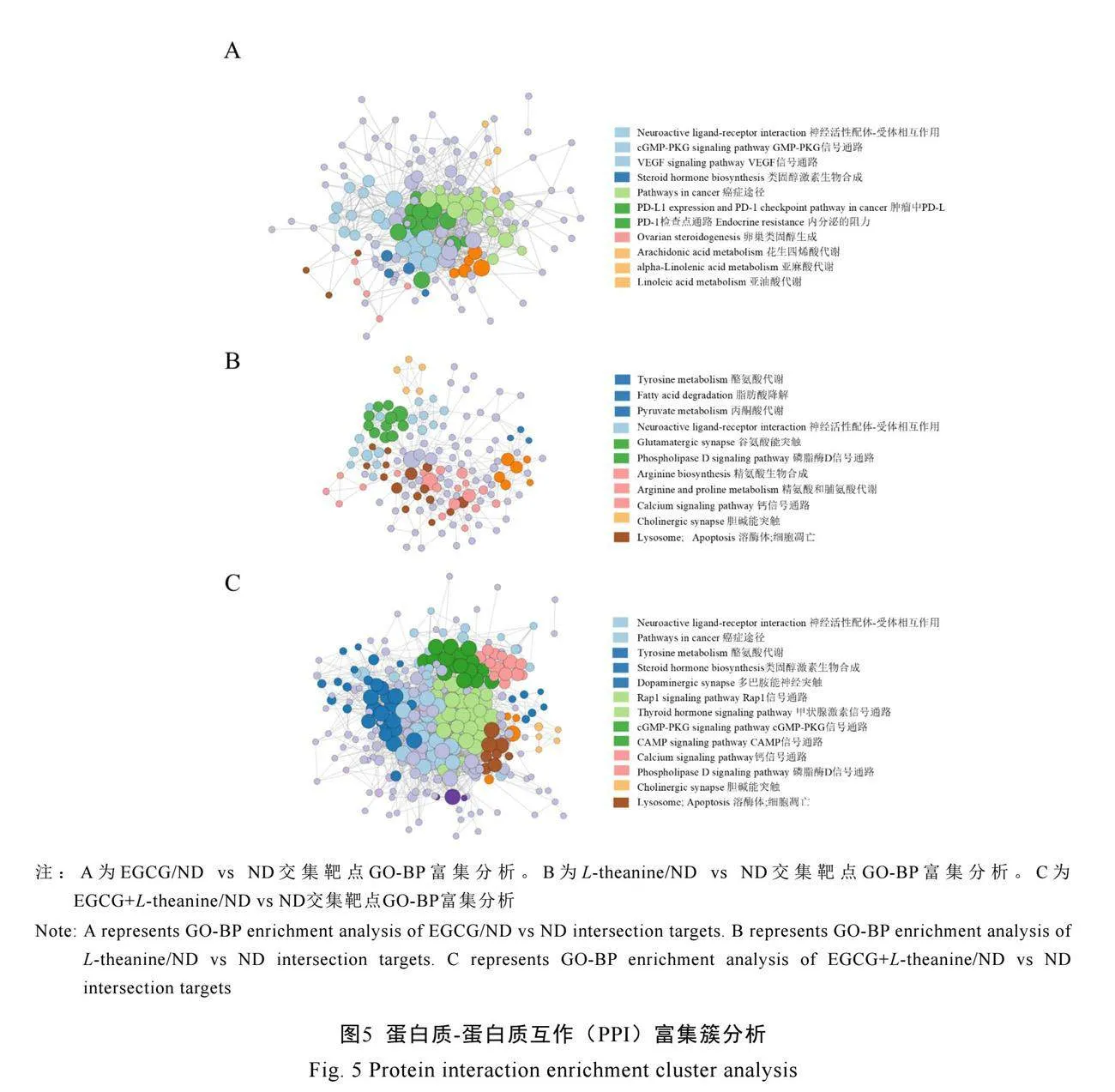
2.3.3 EGCG與L-theanine及其主要代謝物抗NDs的蛋白質互作富集簇分析
蛋白質互作富集簇分析結果表明,EGCG/ND組的交集靶點主要富集在表皮生長因子受體(EGFR)酪氨酸激酶抑制劑、黏附功能、神經配體-受體相互作用和PI3K-Akt信號通路(圖5A)。L-theanine/ND組的交集靶點主要富集在松弛素信號通路、精氨酸和脯氨酸代謝、神經活性配體受體相互作用和脂肪酸代謝途徑(圖5B)。EGCG+L-theanine/ND交集靶點主要富集在上述簇中,其中神經配體-受體相互作用、磷脂酶D信號通路、DNA結合轉錄因子(Rap1)信號通路、環磷腺苷(cAMP)信號通路、細胞凋亡和溶酶體相關富集DE靶點數量和連接緊密度明顯增加(圖5C)。
EGCG及其代謝物可能通過調節細胞穩態
并減輕細胞凋亡途徑的損傷,從而發揮治療NDs的作用。L-theanine及其代謝物則具有調節代謝和促進生長的作用,能夠產生興奮性神經遞質,作用于中樞神經系統。EGCG與L-theanine協同作用,能夠增強EGCG和L-theanine各自的效果。
2.4 轉錄組學和網絡藥理學聯合分析
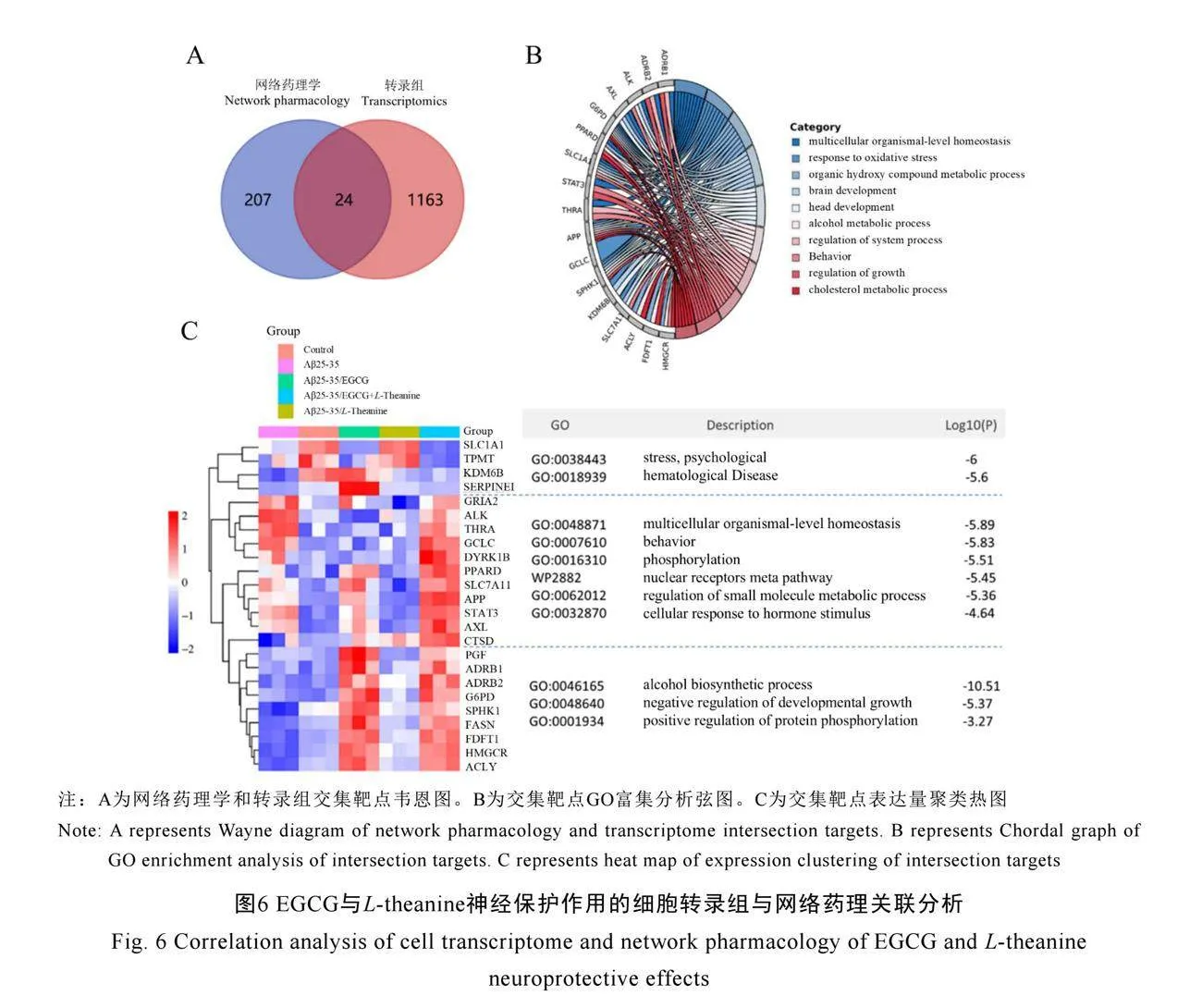
本研究通過細胞轉錄組測序和網絡藥理學方法,對Aβ25-35/EGCG+L-theanine協同處理組的差異基因進行交互分析,識別出24個關鍵交互靶基因(圖6A),這些基因在Aβ25-35處理組與Aβ25-35/EGCG+L-theanine聯合處理組之間顯示出顯著的表達差異。GO分析顯示,這些基因主要富集在膽固醇代謝、生長調節、多細胞有機體水平的穩態、氧化應激反應、腦發育和酒精代謝過程等關鍵生物通路中(圖6B、圖6C)。
同時,EGCG+L-theanine聯合處理上調了AXL、APP、CTSD、GCLC等基因的表達,這些基因主要富集在多細胞有機體穩態、行為調控、磷酸化反應、核受體元途徑和小分子代謝調節等生物過程中。AXL和APP是已知的神經保護因子,參與調節神經元的存活和信號傳導,
表明EGCG和L-theanine可能通過這些基因促進神經元的修復和功能恢復。GCLC編碼谷胱甘肽合成酶,在抗氧化應激反應中起重要作用,其上調可能增強細胞的抗氧化能力,減輕Aβ25-35誘導的氧化損傷。
此外,研究發現,這些基因還富集在酒精生物合成過程、發育生長的負向調節和蛋白質磷酸化的正向調節等通路中,這些過程主要受到EGCG的影響。EGCG通過調控這些通路,維持細胞的靜息狀態和代謝平衡,減少異常細胞生長和增殖[30]。聯合L-theanine的應用,則進一步促進細胞的能量代謝和生長修復,增強了神經保護的效果。表明EGCG和L-theanine在協同減輕細胞應激、維持細胞穩態方面發揮了顯著作用[31-32]。這種協同作用為進一步探索EGCG和L-theanine在神經退行性疾病中的治療機制及其臨床應用提供了重要的理論依據。
3 討論
NDs是老年人的常見疾病,隨著人口老齡化的加劇,其患病率迅速上升[33]。迫切需要更有效的治療策略來遏制NDs的進展,同時深入了解每種治療策略的原因和機制。網絡藥理學的進展為闡明某些藥物治療復雜疾病提供了全新的機會[34]。基于前期研究結果[35],本研究分析了EGCG和L-theanine協同抑制神經退行性改變的作用,并通過網絡藥理與細胞轉錄組的交集靶點分析,揭示了茶葉中的EGCG與L-theanine聯合應用在神經退行性改變中的協同作用。
細胞試驗驗證了EGCG和L-theanine能有效抑制由Aβ25-35誘導的PC12細胞損傷,且聯合應用時表現出明顯的協同效應。EGCG主要通過抑制淀粉樣蛋白的毒性應激和維持細胞靜息狀態來發揮作用,而L-theanine則通過抑制DNA損傷和調節谷氨酸代謝來提供神經保護(圖1和圖2)。這些結果與之前的研究一致[35-38],進一步證實了EGCG和L-theanine在防治NDs中的潛在價值。
細胞試驗顯示,EGCG和L-theanine聯合應用能夠顯著提高細胞活力。轉錄組分析表明,協同組上調的基因與膽固醇和脂質代謝相關,這些基因可能作為神經退行性疾病中的治療靶點。下調基因與蛋白質合成相關(圖2)。在NDs中,蛋白質穩態失衡可能導致錯誤折疊蛋白積累,形成有毒聚集體,導致神經細胞損傷和死亡[39]。因此,通過靜息態效應減少蛋白質的合成可能具有腦保護效果。
EGCG和L-theanine聯合應用展現了顯著的神經保護作用。EGCG在高濃度下(100 μmol·L-1)顯示出促凋亡活性,而在與Aβ25-35聯合處理時卻表現出相反的結果(圖1)。這種現象可能是由于EGCG在高濃度下具有雙重作用:既可以引發氧化應激導致細胞凋亡,又能在Aβ25-35存在情況下通過抗氧化和抗炎機制緩解細胞毒性,從而對細胞產生保護作用。轉錄組分析結果進一步支持這一觀點,顯示EGCG能調控多條與抗氧化應激和脂質代謝相關的信號通路,減輕Aβ25-35的毒性作用,保護細胞免受凋亡(圖2)。這進一步強調了EGCG在神經退行性疾病防治中的復雜作用機制,為其臨床應用提供了重要理論依據。
L-theanine單獨處理未能顯著改善Aβ25-35誘導的PC12細胞損傷(圖1C和圖1D),轉錄組分析也支持這一點,顯示L-theanine處理組的DEGs數量較少,對細胞整體基因表達影響較弱(圖2F)。L-theanine主要通過調節染色質重塑、DNA代謝和免疫過程發揮作用,但在抵抗Aβ25-35誘導的急性毒性應激方面作用有限。相比之下,EGCG處理顯著影響了氧化應激反應、脂質代謝和細胞凋亡路徑(圖2E),增強了細胞的抗逆性。當EGCG與L-theanine聯合應用時,顯著擴展了單獨處理的作用范圍,調控多個關鍵生物過程(圖2G),增強神經保護效果,提高細胞活力(圖1D)。因此,L-theanine單獨應用在神經代謝和突觸功能調控方面有潛力,但在急性細胞毒性應激抵抗方面作用有限,聯合EGCG應用則顯著增強了細胞的抗逆性和修復能力,體現出更強的神經保護效果。這一發現為進一步探索EGCG與L-theanine在神經退行性疾病中的協同機制提供了理論依據。
網絡藥理學分析顯示,EGCG在調節細胞網絡中起著顯著作用,其潛在靶點主要涉及氧化應激、β-淀粉樣蛋白的處理以及細胞凋亡路徑。L-theanine通過參與神經元的突觸信號傳遞,有助于恢復功能失調的谷氨酸穩態。當EGCG和L-theanine聯合應用時,它們在NDs相關的細胞網絡中展示了更廣泛且溫和的調控作用(圖4~圖5)。這種協同效應可能提高了它們在治療NDs中的潛在效果,支持了進一步的臨床研究與應用。
此外,轉錄組學和網絡藥理學的交互分析進一步揭示了EGCG和L-theanine在NDs中的潛在協同機制(圖6)。本研究共識別出24個關鍵交互靶基因,這些基因主要富集在膽固醇代謝、生長調節、細胞穩態、氧化應激反應和腦發育等通路。EGCG通過調節這些通路,維持細胞靜息狀態,減少異常細胞生長和增殖。聯合L-theanine的應用則進一步促進了細胞的能量代謝和生長修復,表明EGCG和L-theanine在減輕細胞應激、維持細胞穩態方面發揮了顯著作用。
本研究通過體外Aβ25-35誘導的PC12細胞AD模型和利用人類疾病靶點數據庫進行的網絡藥理分析,發現EGCG與L-theanine協同抑制淀粉樣蛋白應激、調節代謝并促進神經細胞軸突功能,通過多途徑多靶點抑制神經退行性改變。這項研究首次揭示了EGCG與L-theanine協同作用,通過維護細胞穩態來促進神經細胞修復和再生,為茶葉的神經保護的分子機制提供了基礎研究數據。
然而,本研究還存在一定的局限性,后續的研究可以進一步探索如何優化EGCG和L-theanine的聯合治療策略,以提高其療效和應用范圍。另外,需要進一步聚焦EGCG與L-theanine聯合應用對神經細胞代謝的具體調節機制,并利用神經退行性改變的動物模型,深入研究EGCG和L-theanine在體內的協同作用機制。
參考文獻
[1] Powers E T, Morimoto R I, Dillin A, et al. Biological and chemical approaches to diseases of proteostasis deficiency [J]. Annual Review of Biochemistry, 2009, 78(1): 959-991.
[2] Taylor R C, Dillin A. Aging as an event of proteostasis collapse [J]. Cold Spring Harbor Perspectives in Biology, 2011, 3(5): 328-342. doi: 10.1101/cshperspect.a004440.
[3] Mok K H, Pettersson J, Orrenius S, et al. HAMLET, protein folding, and tumor cell death [J]. Biochemical & Biophysical Research Communications, 2007, 354(1): 1-7.
[4] Chiti F, Dobson C M. Protein misfolding, functional amyloid, and human disease [J]. Annual Review of Biochemistry, 2006, 75(1): 333-366.
[5] Soto C. Alzheimer's and prion disease as disorders of protein conformation: implications for the design of novel therapeutic approaches [J]. Journal of Molecular Medicine, 1999, 77(5): 412-418.
[6] Wallace R A, Dalton A J. What can we learn from study of Alzheimer's disease in patients with down syndrome for early-onset Alzheimer's disease in the general population? [J]. Alzheimer's Reseach &Therapy, 2011, 3(2): 13. doi: 10.1186/alzrt72.
[7] Asaad M, Lee J H. A guide to using functional magnetic resonance imaging to study Alzheimer's disease in animal models [J]. Disease Models and Mechanisms, 2018, 11(5): dmm031724. doi: 10.1242/dmm.031724.
[8] Sheng J G, Zhou X Q, Mrak R E, et al. Progressive neuronal injury associated with amyloid plaque formation in Alzheimer disease [J]. Journal of Neuropathology and Experimental Neurology, 1998(7): 714-717.
[9] Inglis F. The tolerability and safety of cholinesterase inhibitors in the treatment of dementia [J]. International Journal of Clinical Practice Supplement, 2002, 127(127): 45. doi: .1016/S0924-8579(02)00114-0.
[10] Pervin M, Unno K, Takagaki A, et al. Function of green tea catechins in the brain: epigallocatechin gallate and its metabolites [J]. International Journal of Molecular Sciences, 2019, 20(15): 3630. doi: 10.3390/ijms20153630.
[11] Afzal O, Dalhat M H, Altamimi A S A, et al. Green tea catechins attenuate neurodegenerative diseases and cognitive deficits [J]. Molecules, 2022, 27(21): 7604. doi: 10.3390/molecules27217604.
[12] Youn K, Ho C T, Jun M. Multifaceted neuroprotective effects of (-)-epigallocatechin-3-gallate (EGCG) in Alzheimer's disease: an overview of pre-clinical studies focused onβ-amyloid peptide [J]. Food Science and Human Wellness, 2022, 11(3): 11. doi:10.1016/j.fshw.2021.12.006.
[13] Miren E, Amanda C, Patricia R M, et al. Epigallocatechin-3-gallate (EGCG) improves cognitive deficits aggravated by an obesogenic diet through modulation of unfolded protein response in APPswe/PS1dE9 mice [J]. Molecular Neurobiology, 2020, 57(4): 1814-1827.
[14] Walker J M, Klakotskaia D, Ajit D, et al. Beneficial effects of dietary EGCG and voluntary exercise on behavior in an Alzheimer's disease mouse model [J]. Journal of Alzheimers Disease, 2015, 44(2): 561-572.
[15] Unno K, Pervin M, Nakagawa A, et al. Blood-brain barrier permeability of green tea catechin metabolites and their neuritogenic activity in human neuroblastoma SH-SY5Y cells [J]. Molecular Nutrition & Food Research, 2017, 61(12): 1700294. doi:10.1002/mnfr.201700294.
[16] Pervin M, Unno K, Nakagawa A, et al. Blood brain barrier permeability of (-)-epigallocatechin gallate, its proliferation-enhancing activity of human neuroblastoma SH-SY5Y cells, and its preventive effect on age-related cognitive dysfunction in mice [J]. Biochemistry and Biophysics Reports, 2017, 9: 180-186.
[17] Du A Z, Rong D, Cyrollah L, et al. Epigallocatechin-3-gallate, an active ingredient of traditional Chinese medicines, inhibits the 3CLpro activity of SARS-CoV-2 [J]. International Journal of Biological Macromolecules, 2021, 176(1): 1-12.
[18] Wu Z, Yu W, Ni W, et al. Improvement of obesity by Liupao tea is through the IRS-1/PI3K/AKT/GLUT4 signaling pathway according to network pharmacology and experimental verification [J]. Phytomedicine, 2023, 110: 154633. doi:10.1016/j.phymed.2022.154633
[19] Gong Z, Liu Q, Lin L, et al. L-Theanine prevents ETEC-induced liver damage by reducing intrinsic apoptotic response and inhibiting ERK1/2 and JNK1/2 signaling pathways [J]. European Journal of Pharmacology, 2017, 818: 184-190.
[20] Tsai, Wen H, Chung H, et al. L-Theanine inhibits proinflammatory PKC/ERK/ICAM-1/IL-33 signaling, apoptosis, and autophagy formation in substance P-Induced hyperactive bladder in rats [J]. Neurourology and Urodynamics, 2017, 36(2): 297-307.
[21] Unno T, Suzuki Y, Kakuda T, et al. Metabolism of theanine, gamma-glutamylethylamide, in rats [J]. Journal of Agricultural and Food Chemistry, 1999, 47(4): 1593-1596.
[22] Nedergaard M, Takano T, Hansen A J. Beyond the role of glutamate as a neurotransmitter [J]. Nature Reviews Neuroscience, 2002, 3(9): 748-755.
[23] Ben P, Zhang Z, Zhu Y, et al. L-Theanine attenuates cadmium-induced neurotoxicity through the inhibition of oxidative damage and tau hyperphosphorylation [J]. NeuroToxicology, 2016, 57: 95-103.
[24] Lipinski C A, Lombardo F, Dominy B W, et al. Experimental and computational approaches to estimate solubility and permeability in drug discovery and development settings [J]. Advanced Drug Delivery Reviews, 2001, 46(1/2/3): 3-26.
[25] Daina A, Michielin O, Zoete V. SwissADME: a free web tool to evaluate pharmacokinetics, drug-likeness and medicinal chemistry friendliness of small molecules [J]. Scientific Reports. 2017, 7: 42717. doi: 10.1038/srep42717.
[26] Wang Y X, Zhang S, Li F C, et al. Therapeutic target database 2020: enriched resource for facilitating research and early development of targeted therapeutics [J]. Nucleic Acids Research, 2020, 48(D1): 1031-1041.
[27] Bader G D, Hogue C W V. An automated method for finding molecular complexes in large protein interaction networks [J]. BMC Bioinformatics, 2003, 4: 2. doi: 10.1186/1471-2105-4-2.
[28] Zhou Y, Zhou B, Pache L, et al. Metascape provides a biologist-oriented resource for the analysis of systems-level datasets [J]. Nature Communications, 2019, 10(1): 1523. doi: 10.1038/s41467-019-09234-6.
[29] Papassotiropoulos A, Gerhards C, Heck A, et al. Human genome-guided identification of memory-modulating drugs [J]. Proceedings of the National Academy of Sciences, 2013, 110(46): 4369-4374.
[30] Gon?alves P B, Sodero A C R, Cordeiro Y. Green tea epigallocatechin-3-gallate (EGCG) targeting protein misfolding in drug discovery for neurodegenerative diseases [J]. Biomolecules, 2021, 11(5): 767. doi: 10.3390/biom11050767.
[31] 劉寶貴, 陳致印, 張楊玲, 等. L-茶氨酸與表沒食子兒茶素沒食子酸酯預防肥胖及高膽固醇血癥的協同調節作用[J]. 食品工業科技, 2022, 43(3): 341-350.
Liu B G, Chen Z Y, Zhang Y L,et al. Synergistic moderating effects of L-theanine and EGCG for the prevention of obesity and Hypercholesterolemia [J]. Science and Technology of Food Industry, 2022, 43(3): 341-350.
[32] 彭影琦, 袁冬寅, 林玲, 等. 表沒食子兒茶素沒食子酸酯對L-茶氨酸調節小鼠血清及腸道游離氨基酸的影響[J]. 食品科學, 2020, 41(19): 154-160.
Peng Y Q, Yuan D Y, Lin L, et al. Influence of epigallocatechin gallate on the regulation effect of L-theanine on eerum and intestinal free amino acids in mice [J]. Food Science, 2020, 41(19): 154-160.
[33] Bianchi V E, Herrera P F, Laura R. Effect of nutrition on neurodegenerative diseases. A systematic review [J]. Nutr Neurosci, 2021, 24(10): 810-834.
[34] Zhang W, Bai Y, Wang Y, et al. Polypharmacology in drug discovery: a review from systems pharmacology perspective [J]. Current Pharmaceutical Design, 2016, 22(21): 3171-3181.
[35] Xie X, Wan J, Zheng X, et al. Synergistic effects of epigallocatechin gallate and L-theanine in nerve repair and regeneration by anti-amyloid damage, promoting metabolism, and nourishing nerve cells [J]. Frontiers in Nutrition, 2022, 9: 951415. doi: 10.3389/fnut.2022.951415.
[36] Mandel S, Amit T, Reznichenko L, et al. Green tea catechins as brain-permeable, natural iron chelators-antioxidants for the treatment of neurodegenerative disorders [J]. Molecular Nutrition & Food Research, 2006, 50(2): 229-234.
[37] Unno K, Yamada H, Iguchi K, et al. Anti-stress effect of green tea with lowered caffeine on humans: a pilot study [J]. Biological & Pharmaceutical Bulletin, 2017, 40(6): 902. doi: 10.1248/bpb.b17-00141.
[38] Kim T I, Lee Y K, Park S G, et al. L-Theanine, an amino acid in green tea, attenuates beta-amyloid-induced cognitive dysfunction and neurotoxicity: reduction in oxidative damage and inactivation of ERK/p38 kinase and NF-kappaB pathways [J]. Free Radical Biology and Medicine, 2009, 47(11): 1601-1610.
[39] Morley J E, Armbrecht H J, Farr S A, et al. The senescence accelerated mouse (SAMP8) as a model for oxidative stress and Alzheimer's disease [J]. Biochim Biophys Acta, 2012, 1822(5): 650-656.
