低溫脅迫下工業(yè)大麻中大麻二酚含量和差異表達(dá)基因分析
2024-09-27 00:00:00王文重張利國房郁妍鄭楠張明閆博巍張元野隋月
江蘇農(nóng)業(yè)科學(xué) 2024年15期

摘要:為研究低溫脅迫條件下龍大麻6號大麻二酚(CBD)積累的變化規(guī)律和合成途徑分子調(diào)控機制,以不同低溫脅迫和常溫條件處理大麻葉片為材料,測定CBD含量,同時對上述材料進(jìn)行轉(zhuǎn)錄組測序。通過對不同低溫脅迫和常溫條件處理大麻葉片CBD含量變化,發(fā)現(xiàn)0 ℃處理組(T1)CBD含量最高,其他低溫處理組5 ℃處理組(T2)、15 ℃處理組(T3)、20 ℃處理組(T4)大麻葉中CBD含量比常溫組(CK)CBD含量低。轉(zhuǎn)錄組數(shù)據(jù)中CBD合成途徑關(guān)鍵基因差異表達(dá)分析顯示,0 ℃處理組CBD合成途徑中大部分關(guān)鍵基因表達(dá)量都顯著上調(diào),同時其他溫度脅迫條件下CBD合成途徑中大部分關(guān)鍵基因表達(dá)量也都有變化。0 ℃能夠顯著提高龍大麻6號CBD的合成,提高植株體內(nèi)CBD合成途徑中關(guān)鍵基因表達(dá)量,從而使其更好地適應(yīng)低溫環(huán)境。
關(guān)鍵詞:大麻;大麻二酚;低溫脅迫;差異表達(dá)基因
中圖分類號:S563.301 文獻(xiàn)標(biāo)志碼:A
文章編號:1002-1302(2024)15-0072-13
收稿日期:2023-11-17
基金項目:黑龍江省省屬科研院所科研業(yè)務(wù)費項目(編號:CZKYF2023-1-B007)。
作者簡介:王文重(1979—),女,博士,副研究員,主要從事工業(yè)大麻遺傳育種研究。E-mail:wenwen0331@163.com。
通信作者:張利國,博士,研究員,主要從事麻類遺傳基礎(chǔ)研究。E-mail:zlg86@aliyun.com。
大麻(Cannabis sativa L.)是一種高附加值的多用途經(jīng)濟(jì)作物,屬于桑科大麻屬,廣泛分布于世界各地,在我國有著悠久的栽培歷史[1-3]。目前國內(nèi)已審定或認(rèn)定或登記的大麻品種,都是工業(yè)大麻,不含或含有極低神經(jīng)成癮性物質(zhì),不具毒品利用價值[4]。工業(yè)大麻中存在多種大麻素,主要在大麻葉和花中,大麻二酚(cannabidiol,CBD)是其中已被聯(lián)合國世界衛(wèi)生組織認(rèn)可的健康成分,研究表明,CBD無致幻成癮性,且具有抗驚厥痙攣、癲癇、鎮(zhèn)痛、焦慮、抗菌抗炎、抗麻醉和神經(jīng)系統(tǒng)保護(hù)等藥理功效,廣泛應(yīng)用于醫(yī)藥和化妝品領(lǐng)域[5-7]。在CBD產(chǎn)業(yè)帶領(lǐng)下,工業(yè)大麻產(chǎn)業(yè)有望在未來幾年實現(xiàn)強勁增長,到2025年CBD產(chǎn)業(yè)預(yù)計將達(dá)到千億美元級別[8]。低溫是影響植物正常生長的主要障礙之一,大多數(shù)植物對低溫較為敏感,一旦遭遇低溫脅迫,將導(dǎo)致生長受限,產(chǎn)量下降影響植物的生長發(fā)育和次生代謝產(chǎn)物的合成[9]。溫度是調(diào)控植物生長發(fā)育和代謝物質(zhì)積累的重要條件,國內(nèi)外關(guān)于溫度對工業(yè)大麻生長和CBD積累研究的相關(guān)報道較少。黑龍江省地處我國北方,春季播種時間溫度較低,對大麻的生長和CBD含量會造成一定的影響。因此需要開展大麻對低溫脅迫響應(yīng)方面的研究,了解大麻在CBD含量變化規(guī)律和合成途徑分子機制。目前,黑龍江省工業(yè)大麻原料來源多樣,溫度與CBD含量、原料產(chǎn)率存在密切的關(guān)系,掌握溫度與大麻CBD有效成分含量之間的關(guān)系,也為今后利用植物基因工程技術(shù)提高工業(yè)大麻的品質(zhì)奠定研究基礎(chǔ),將有效提升花葉用工業(yè)大麻產(chǎn)業(yè)的市場競爭力與經(jīng)濟(jì)效益。
1 材料與方法
1.1 植物材料與處理
試驗大麻品種為龍大麻6號,由黑龍江省農(nóng)業(yè)科學(xué)院經(jīng)濟(jì)作物研究所提供。CK代表常溫(25 ℃)處理組;T1代表0 ℃(處理1)處理組;T2代表5 ℃(處理2)處理組;T3代表15 ℃(處理3)處理組;T4代表20 ℃(處理4)處理組。
試驗材料2021年種植于黑龍江省農(nóng)業(yè)科學(xué)院經(jīng)濟(jì)作物研究所實驗室人工氣候溫箱內(nèi),培養(yǎng)條件設(shè)定為相對濕度80%、光—暗周期14 h—10 h,全程充分供水。分別在0 ℃(處理1)、5 ℃(處理2)、15 ℃(處理3)、20 ℃(處理4)、CK(25 ℃)下進(jìn)行培養(yǎng)。待植株長大至花期進(jìn)行取樣,為了消除個體差異,選擇植株長勢、外觀表型、部位盡量一致的大麻葉取樣,每株大麻取5~8張葉片,3次生物學(xué)重復(fù),用錫箔紙包好后于-80 ℃保存?zhèn)溆谩?/p>
1.2 大麻二酚(CBD)含量測定
分別取不同環(huán)境下培養(yǎng)后的鮮大麻葉經(jīng)85 ℃烘3 h并粉碎后的大麻葉0.200 g,加入甲醇4 mL,采用常溫超聲提取,提取時間為10 min,靜置1 h。4 000 r/min 離心5 min,取上清液轉(zhuǎn)移至10 mL容量瓶中進(jìn)行定容。使用2 mL注射器裝樣過 0.45 μm 有機濾膜注射至液相進(jìn)樣瓶,采用HPLC色譜條件分析CBD含量。
1.3 RNA的提取及cDNA文庫的合成
使用全式金生物技術(shù)有公司試劑盒分別提取處理組和對照組總RNA,并用Bioanalyzer 2100及Agilent RNA6000 Nano Kit對總RNA的完整性和純度進(jìn)行分析。
測序由哈爾濱博泰生物科技有限公司完成,使用Illumina HiSeqTM X TEN測序儀進(jìn)行測序。使用HISAT2將clean reads比對到參考基因組(https://gofile-3663784363.cn4.quickconnect.cn/fsdownload/QZM0L6zNu/Cannatonic.genome.fasta.gz)。篩選差異表達(dá)基因條件為:表達(dá)差異倍數(shù) |log2FoldChange|>1,顯著性P<0.01。
1.4 qRT-PCR驗證
使用全式金生物技術(shù)有公司試劑盒提取總RNA后反轉(zhuǎn)錄為cDNA,利用Primer 5軟件設(shè)計引物,采用Actin為內(nèi)參基因(表1)。
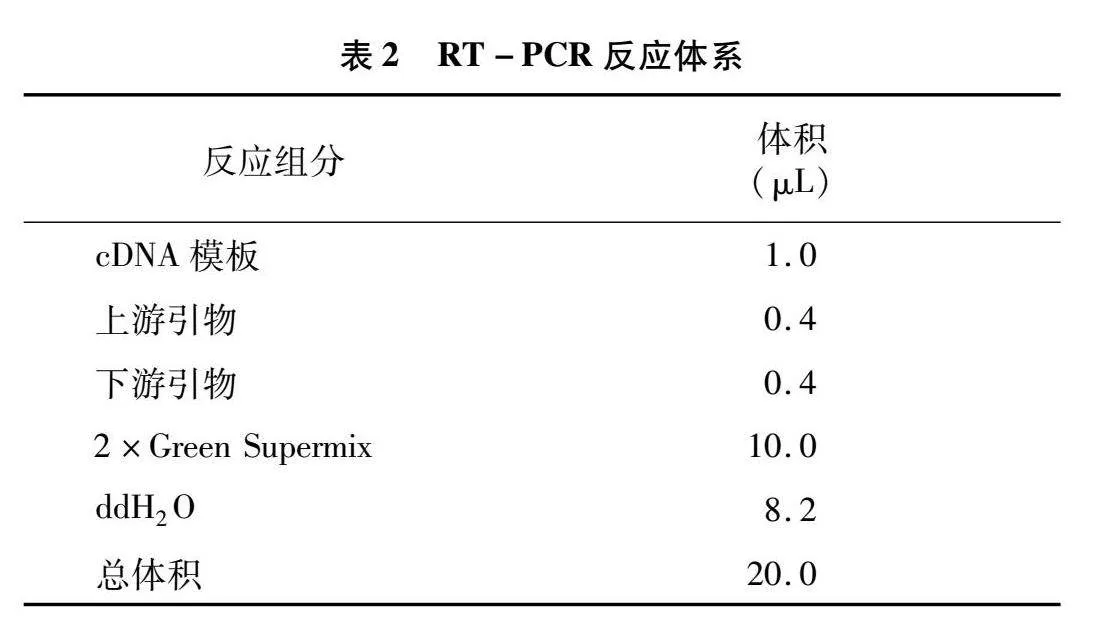
應(yīng)用全式金公司熒光定量試劑盒進(jìn)行熒光定量試驗,反應(yīng)體系見表2。反應(yīng)程序為:94 ℃ 30 s;94 ℃ 5 s,57 ℃ 15 s,72 ℃ 10 s,40個循環(huán)。應(yīng)用2-ΔΔCT方法計算相對定量結(jié)果。相關(guān)基因qRT-PCR引物如表1所示。對篩選的關(guān)鍵基因進(jìn)行 qRT-PCR,測定相對表達(dá)量,比熒光定量試驗結(jié)果與轉(zhuǎn)錄組數(shù)據(jù)趨勢是否一致。
1.5 數(shù)據(jù)分析
每組試驗重復(fù)3次,使用Excel軟件處理數(shù)據(jù)及對結(jié)果作圖,SPSS 25.0軟件對數(shù)據(jù)進(jìn)行顯著性分析(α=0.05)。
2 結(jié)果與分析
2.1 不同溫度處理大麻二酚(CBD)含量變化
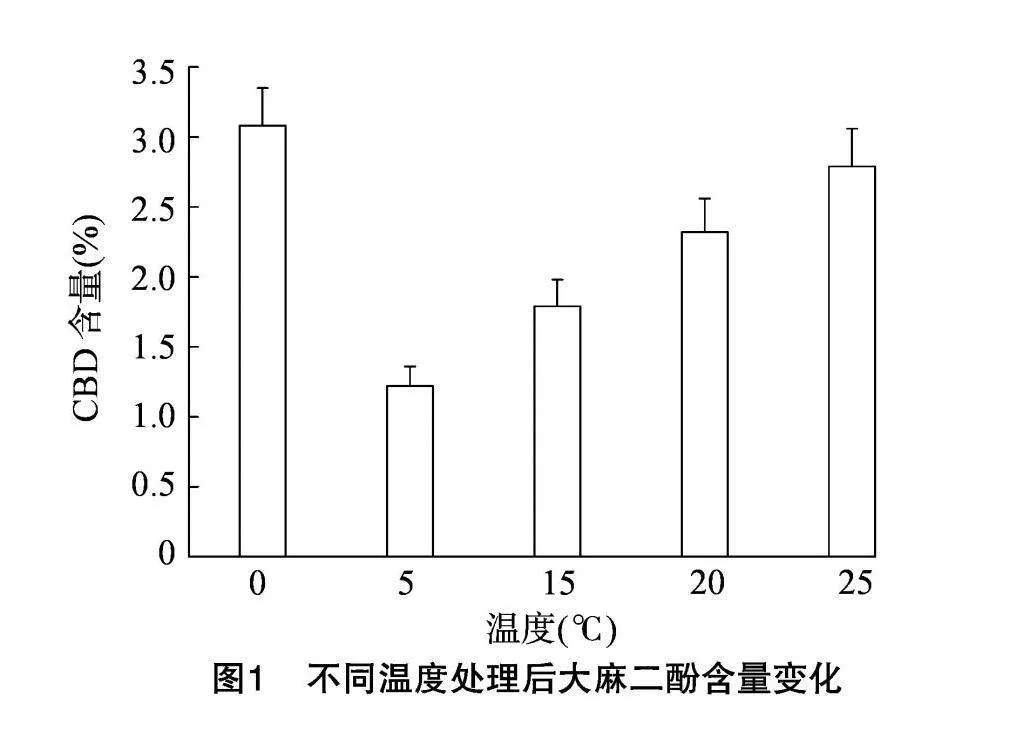
由圖1可知,0 ℃處理后CBD含量最高,5 ℃處理后含量最低。處理溫度從5 ℃開始,隨著處理溫度升高,CBD含量逐漸增加,但15℃和20℃處理后,CBD含量差異不顯著。
2.2 轉(zhuǎn)錄組分析
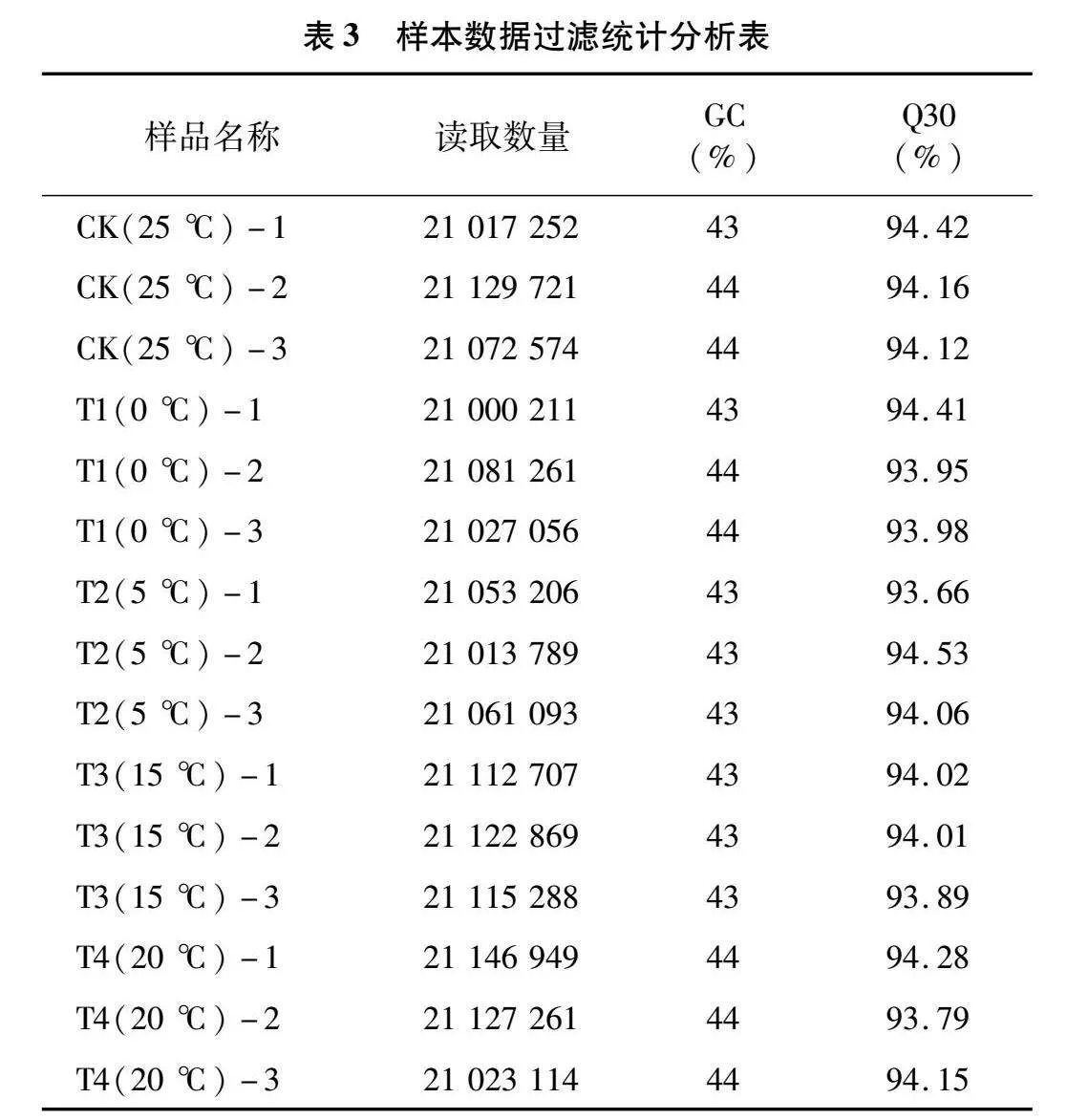
2.2.1 數(shù)據(jù)整理 工業(yè)大麻植株經(jīng)過不同溫度處理之后,進(jìn)行相應(yīng)的轉(zhuǎn)錄組測序分析。對每個樣品的下機數(shù)據(jù)(Raw Data)分別進(jìn)行統(tǒng)計,包括Q30(%)、模糊堿基和GC(%)。統(tǒng)計結(jié)果如表3所示,Q30均在90%以上,說明轉(zhuǎn)錄組測序數(shù)據(jù)質(zhì)量較好,可用于進(jìn)一步分析。
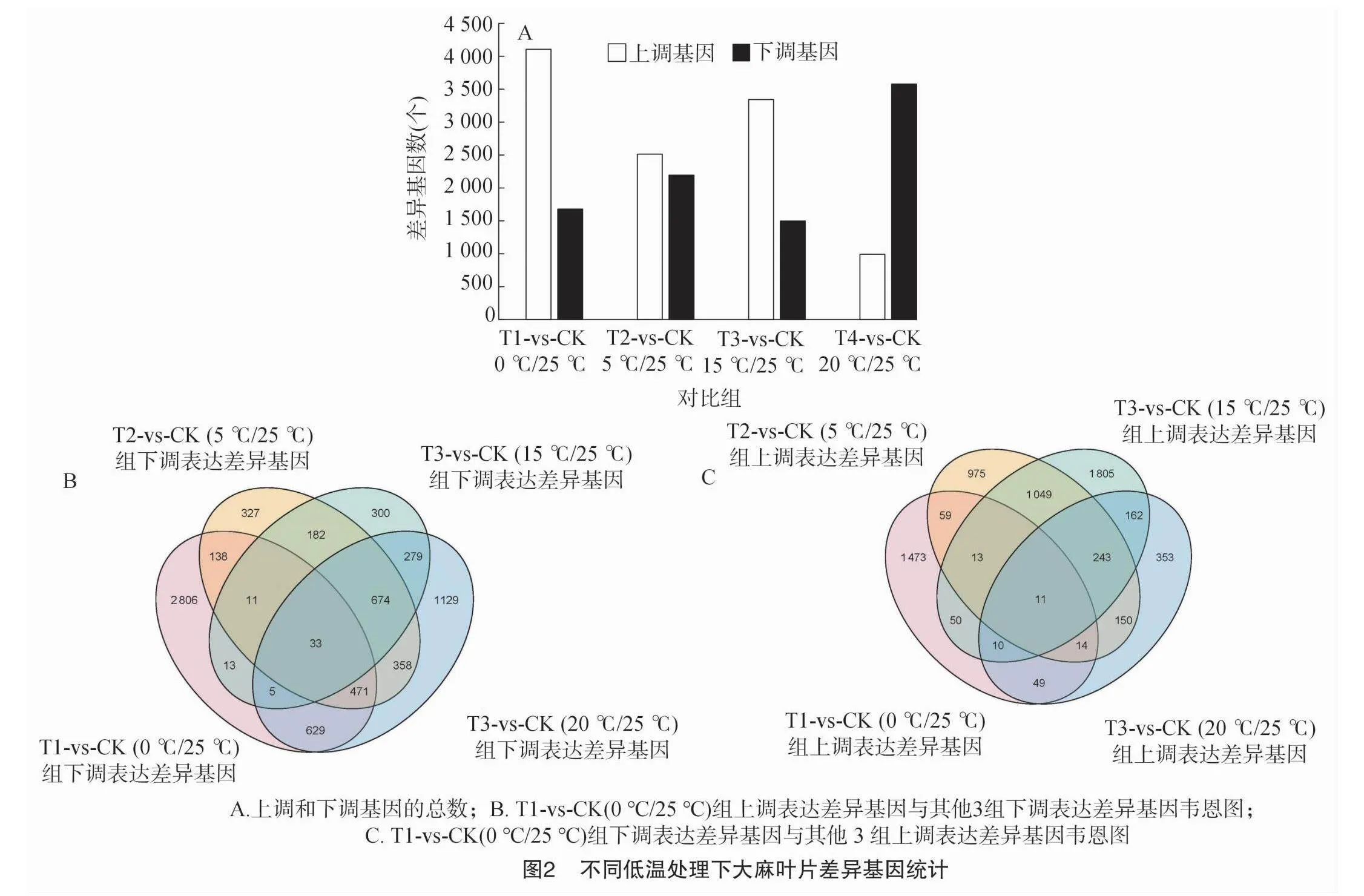
2.2.2 差異基因表達(dá)及富集分析 edgeR以count計數(shù)為起始數(shù)據(jù),經(jīng)過TMM標(biāo)準(zhǔn)化后,進(jìn)行差異表達(dá)分析,結(jié)果如圖2所示。T1與對照組(CK)之間共有5 785個差異基因,其中4 106個上調(diào)基因,1 679 個下調(diào)基因;T2與對照組(CK)之間共有 4 707 個差異基因,其中2 513個上調(diào)基因,2 194個下調(diào)基因;T3與對照組(CK)之間共有48 440個差異基因,其中3 343個上調(diào)基因,1 497個下調(diào)基因;T4與對照組(CK)之間共有4 570個差異基因,其中992個上調(diào)基因,3 578個下調(diào)基因(圖2-A)。結(jié)果顯示,不同低溫處理后,T1-vs-CK(0 ℃/25 ℃)、T2-vs-CK(5 ℃/25 ℃)和T3-vs-CK(15 ℃/25 ℃)上調(diào)差異表達(dá)基因數(shù)大于下調(diào)差異表達(dá)基因數(shù)。T4-vs-CK(20 ℃/25 ℃)下調(diào)差異表達(dá)基因數(shù)遠(yuǎn)遠(yuǎn)大于上調(diào)差異表達(dá)基因數(shù)(圖2-A)。整體看,處理溫度從0 ℃逐漸升高至20 ℃,不同處理組與對照組相比上調(diào)基因數(shù)逐漸減少,下調(diào)基因數(shù)逐漸增加,其中T4-vs-CK(20 ℃/25 ℃)組下調(diào)基因數(shù)是上調(diào)基因數(shù)的3.6倍。在4個比較組中,T1-vs-CK(0 ℃/25 ℃)組上調(diào)表達(dá)差異基因,與T2-vs-CK(5 ℃/25 ℃)、T3-vs-CK(15 ℃/25 ℃)和T4-vs-CK(20 ℃/25 ℃)組下調(diào)基因進(jìn)行比較,結(jié)果顯示33個基因為共有的顯著差異基因(圖2-B)。T1-vs-CK(0 ℃/25 ℃)組下調(diào)表達(dá)差異基因,與T2-vs-CK(5 ℃/25 ℃)、T3-vs-CK(15 ℃/25 ℃)和T4-vs-CK(20 ℃/25 ℃)組上調(diào)基因進(jìn)行比較,結(jié)果顯示11個基因為共有的顯著差異基因(圖2-C)。這些結(jié)果表明,工業(yè)大麻在應(yīng)對不同的低溫脅迫時,可以觸發(fā)相似的反應(yīng),同時處理1可能還存在獨特的反應(yīng)和機制。
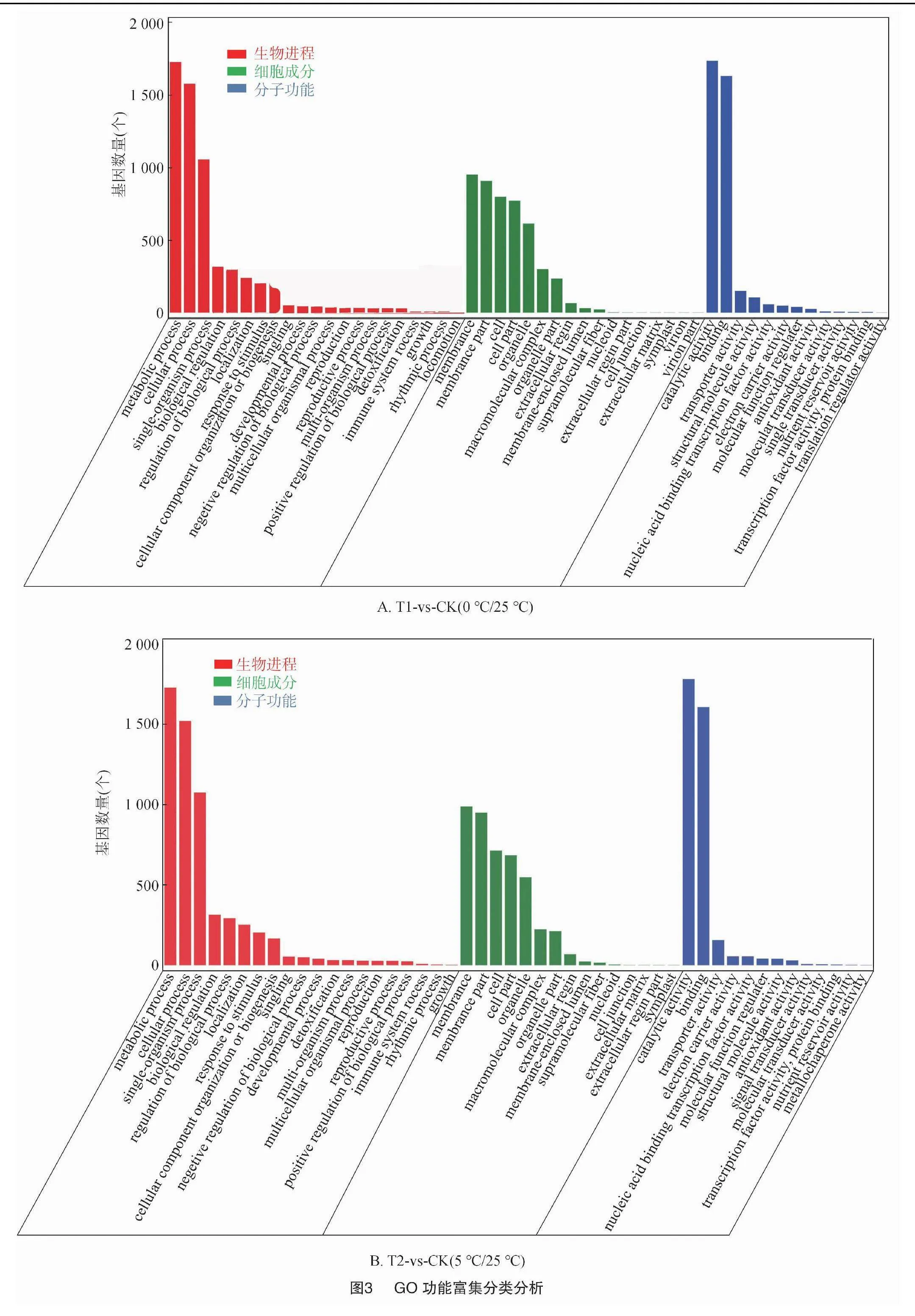
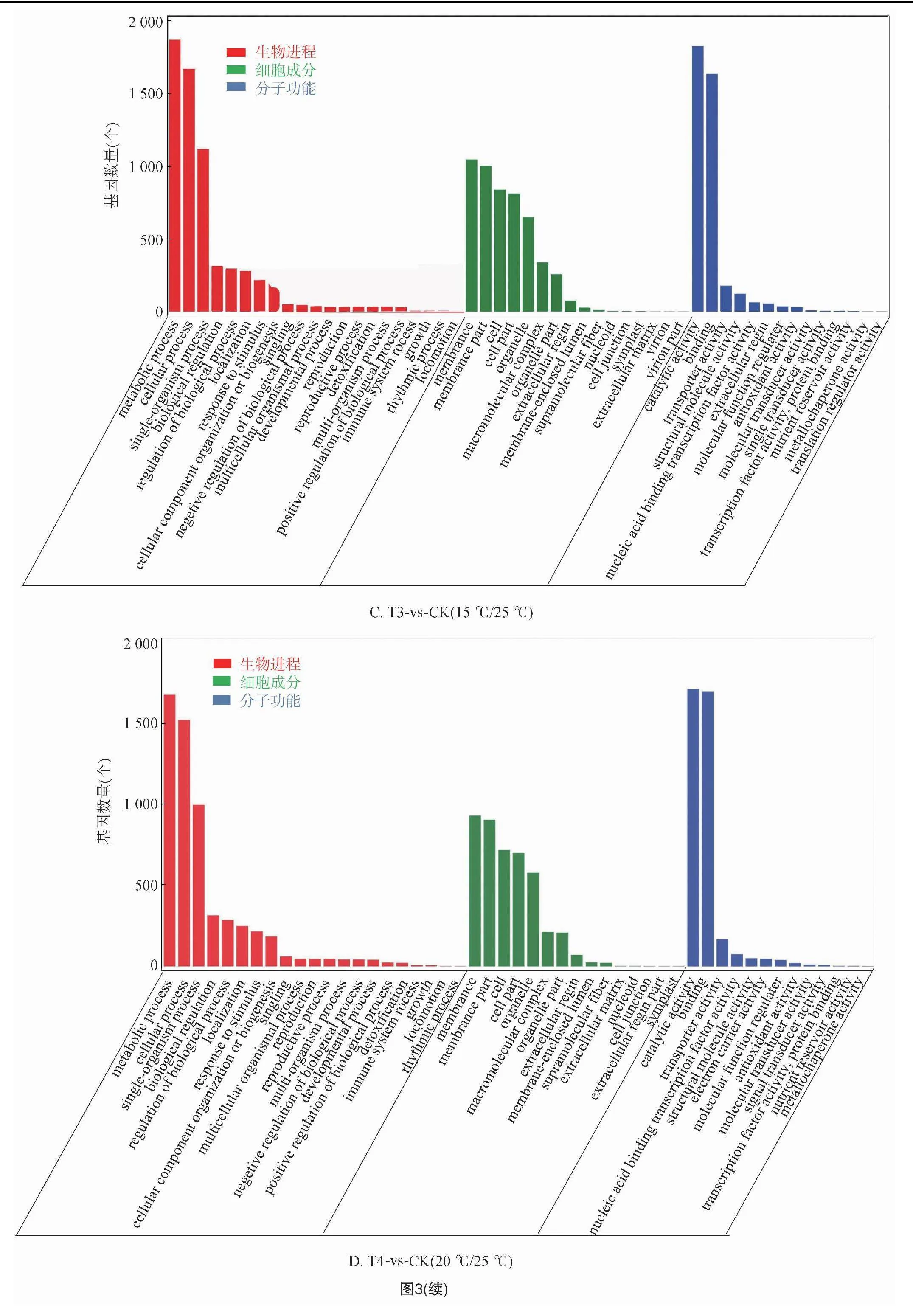
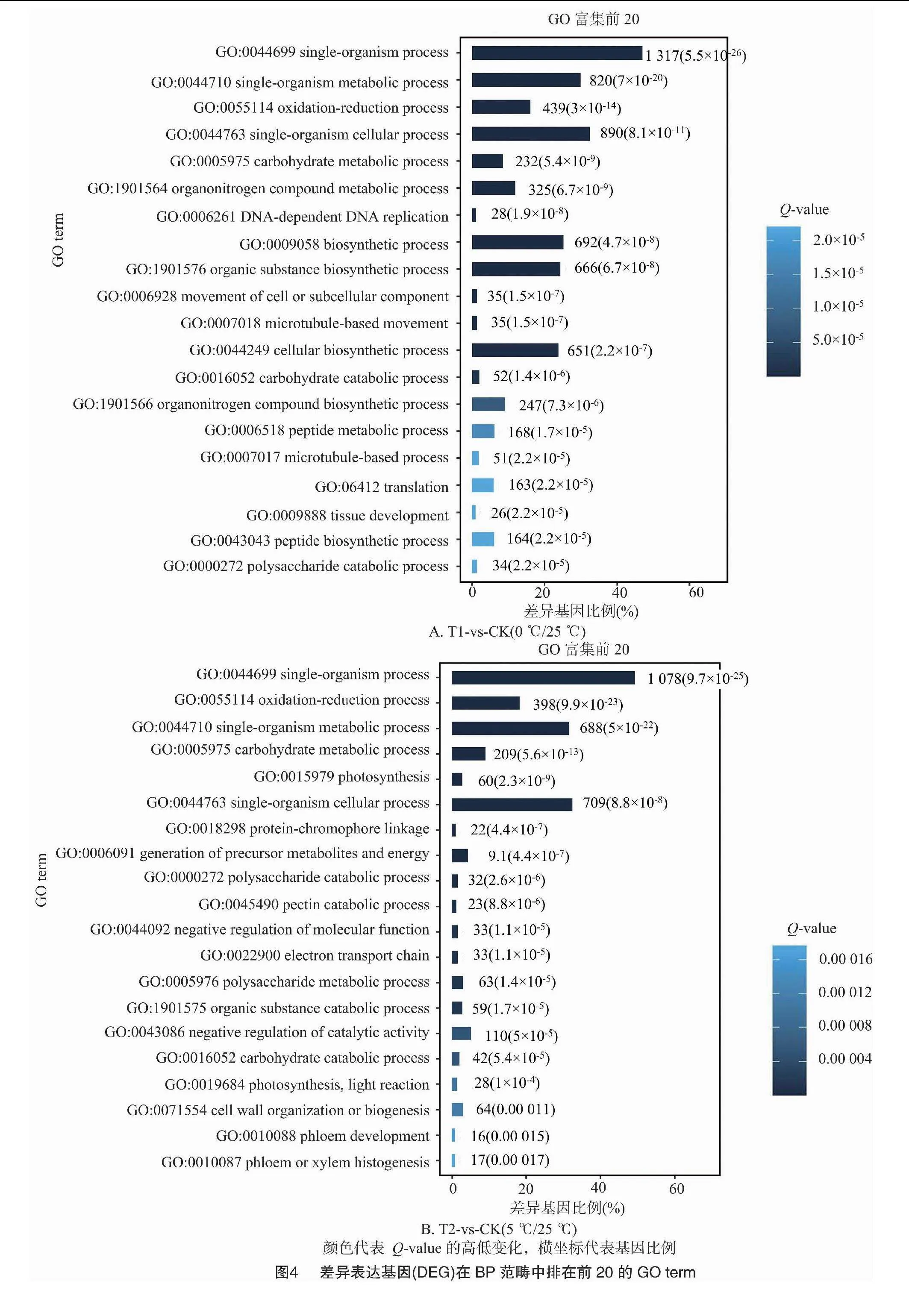
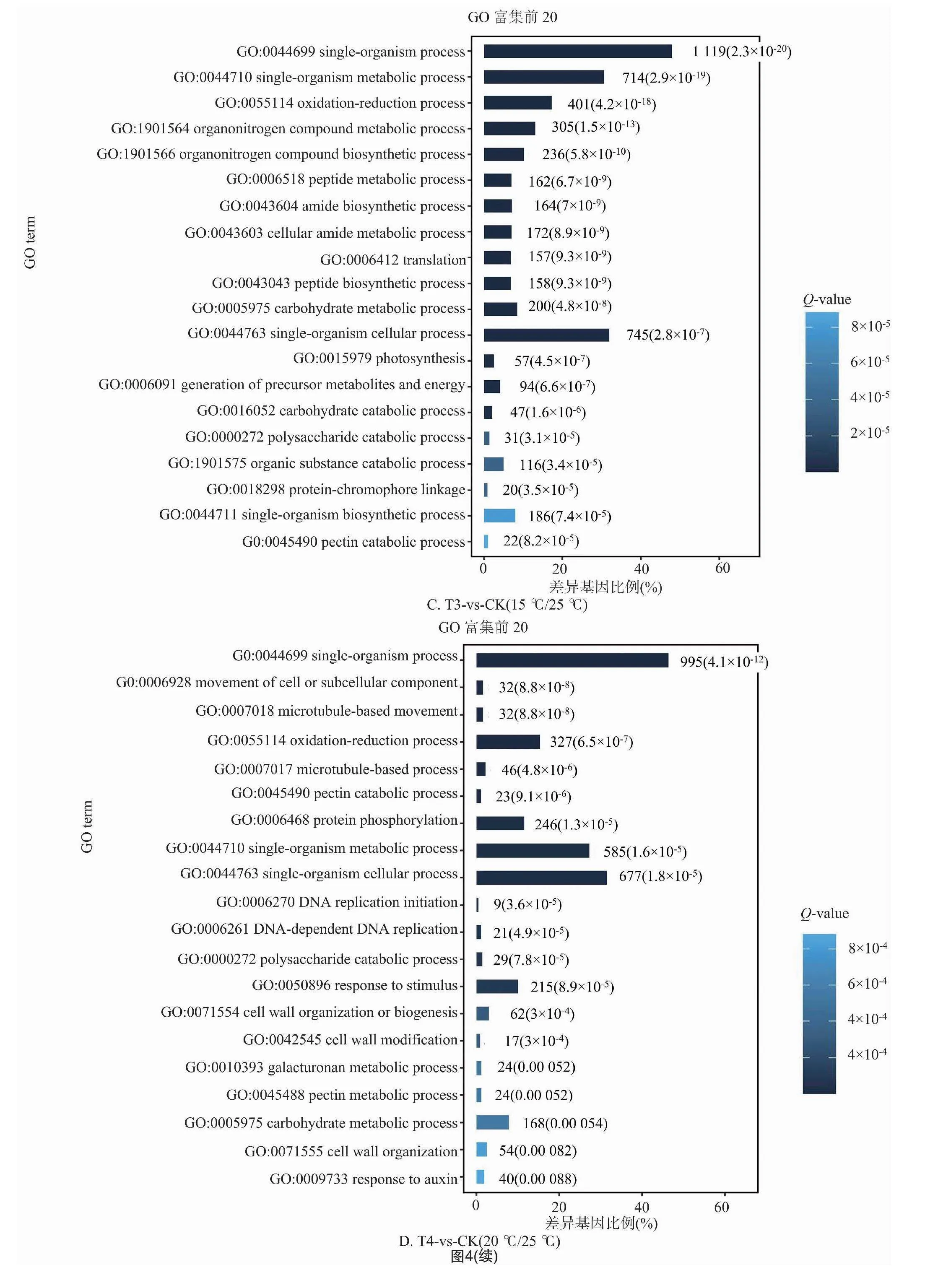
2.2.3 差異基因的GO富集分析 對不同溫度處理工業(yè)大麻差異表達(dá)基因所具有的功能進(jìn)行GO功能富集分析,結(jié)果發(fā)現(xiàn)4組間的差異表達(dá)基因分別富集55、48、51、49條GO terms中,這些GO terms屬于三大功能類別:細(xì)胞成分(CC)、分子功能(MF)、生物進(jìn)程(BP)。4組分析獲得的GO terms很相似。CC中最豐富的成分被歸類為膜(GO:0016020,membrane)、細(xì)胞膜組分(GO:0044425,membrane part)以及細(xì)胞(GO:0005623,cell)。在MF中富集最多的2條GO terms是催化活性(GO:0003824,catalytic activity)和結(jié)合(GO:0005488,binding)。在BP中富集最多的3條GO terms是代謝過程(GO:0008152,metabolic process)、細(xì)胞進(jìn)程(GO:0009987,cellular process)和單一有機體過程(GO:0044699,single-organism process)(圖3)。
在工業(yè)大麻葉片相應(yīng)溫度脅迫處理的基因功能富集中,在BP分類中排在前20的GO terms中主要與合成和代謝相關(guān),包括單一生物過程(GO:0044699,single-organism process)、單生物代謝過程(GO:0044710,single-organism metabolic process)、氧化還原過程(GO:0055114,oxidation-reduction process),此外還有單生物細(xì)胞過程(GO:0044763,single-organism cellular process)、細(xì)胞或亞細(xì)胞成分的運動(GO:0006928 movement of cell or subcellular component)、有機氮化合物代謝過程(GO:1901564,organonitrogen compound metabolic process)、碳水化合物代謝過程(GO:0005975,carbohydrate metabolic process)、果膠分解代謝過程(GO:0045490,pectin catabolic process)(圖4)。
2.2.4 差異共表達(dá)基因的GO分析
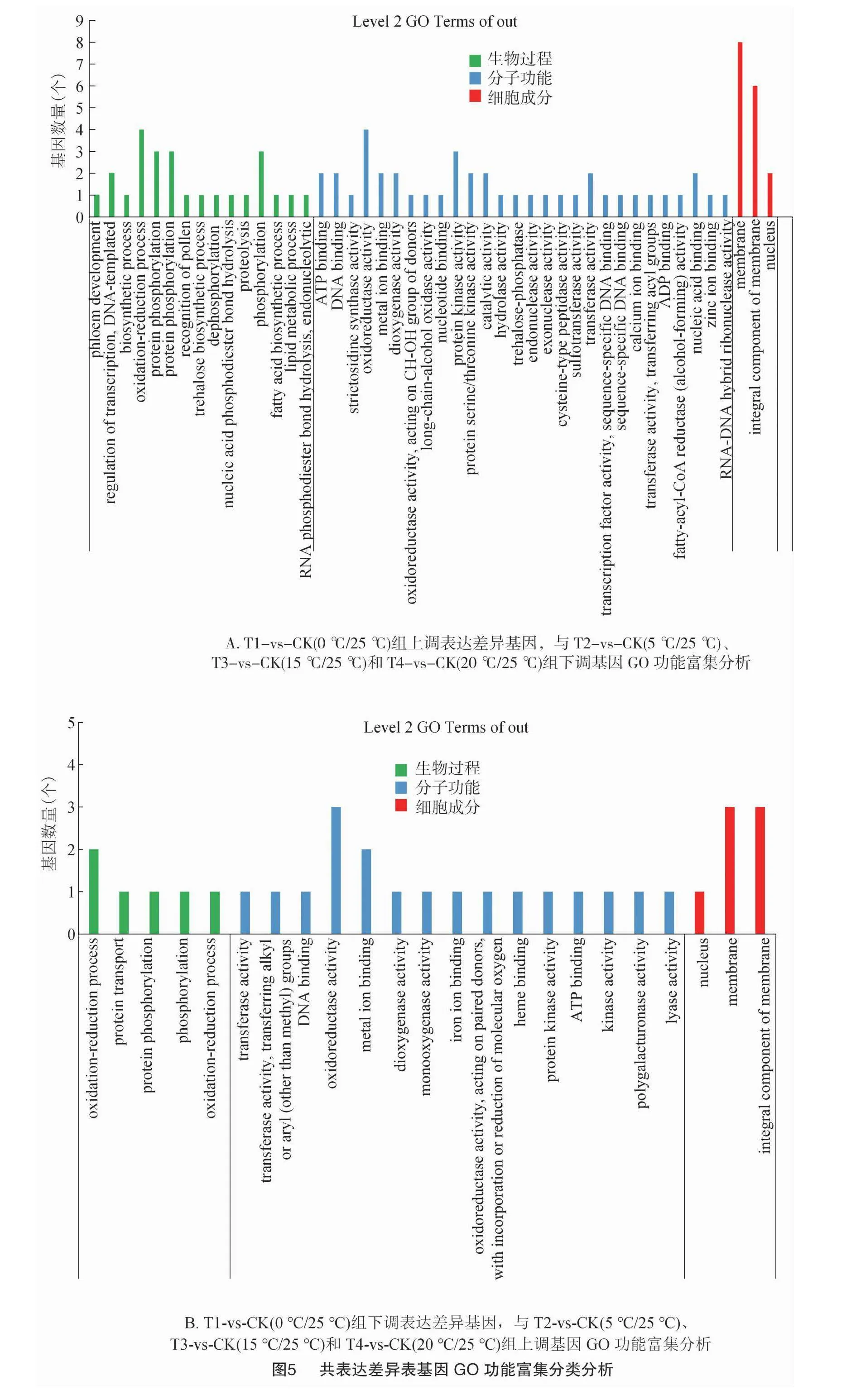
T1-vs-CK(0 ℃/25 ℃)組上調(diào)表達(dá)差異基因,與T2-vs-CK(5 ℃/25 ℃)、T3-vs-CK(15 ℃/25 ℃)和 T4-vs-CK(20 ℃/25 ℃)組下調(diào)基因共表達(dá)的33個基因進(jìn)行GO分析發(fā)現(xiàn),CC中最豐富的成分被歸類為膜(GO:0016020,membrane),在MF中是氧化還原酶活性(GO:0016491,oxidoreductaseactivity),在BP中富集最多的是氧化還原過程(GO:0055114,oxidation-reduction process)(圖5-A)。T1-vs-CK(0 ℃/25 ℃)組下調(diào)表達(dá)差異基因,與T2-vs-CK(5 ℃/25 ℃)、T3-vs-CK(15 ℃/25 ℃)和T4-vs-CK(20 ℃/25 ℃)組上調(diào)基因共表達(dá)的11個基因進(jìn)行GO分析發(fā)現(xiàn),CC中最豐富最多的2條GO terms是膜(GO:0016020,membrane)和細(xì)胞膜組成成分(GO:0016021,integral component of membrane),在MF中是氧化還原酶活性(GO:0016491,oxidoreductase activity),在BP中富集最多的是氧化還原過程(GO:0055114,oxidation-reduction process)(圖5-B)。
2.2.5 差異基因的KEGG富集分析
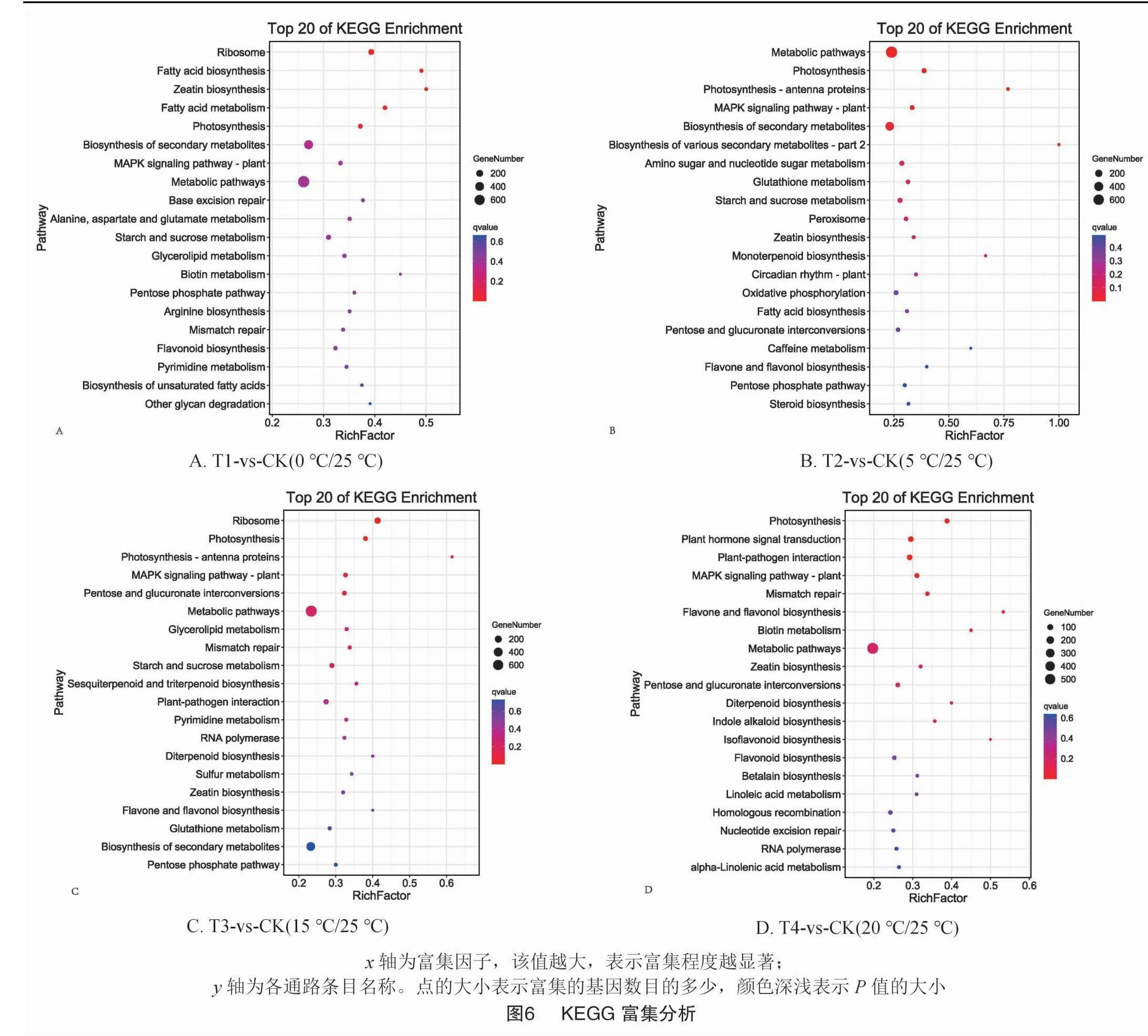
根據(jù)差異表達(dá)基因的KEGG富集分析結(jié)果,分別有139、136、134、127個代謝途徑參與了T1-vs-CK(0 ℃/25 ℃)、T2-vs-CK(5 ℃/25 ℃),T3-vs-CK(15 ℃/25 ℃)和T4-vs-CK(20 ℃/25 ℃)組。主要包括玉米素生物合成(zeatin biosynthesis)、光合作用(photosynthesis)、植物MAPK信號通路(MAPK signaling pathway-plant)、代謝通路(metabolic pathways),還有次級代謝物的生物合成(biosynthesis of secondary metabolites)、淀粉和蔗糖代謝(starch and sucrose metabolism)、黃酮和黃酮醇生物合成(flavone and flavonol biosynthesis)、戊糖磷酸途徑(pentose phosphate pathway),戊糖和葡萄糖酸鹽相互轉(zhuǎn)化(pentose and glucuronate interconversions)和錯配修復(fù)(mismatch repair)。本研究中由不同溫度處理后調(diào)控的排在前20的代謝途徑如圖6所示。這些結(jié)果表明,在不同溫度處理下,工業(yè)大麻的葉片會積極調(diào)整自身代謝以增加環(huán)境適應(yīng)的能力。
2.2.6 CBD合成途徑基因響應(yīng)不同溫度脅迫
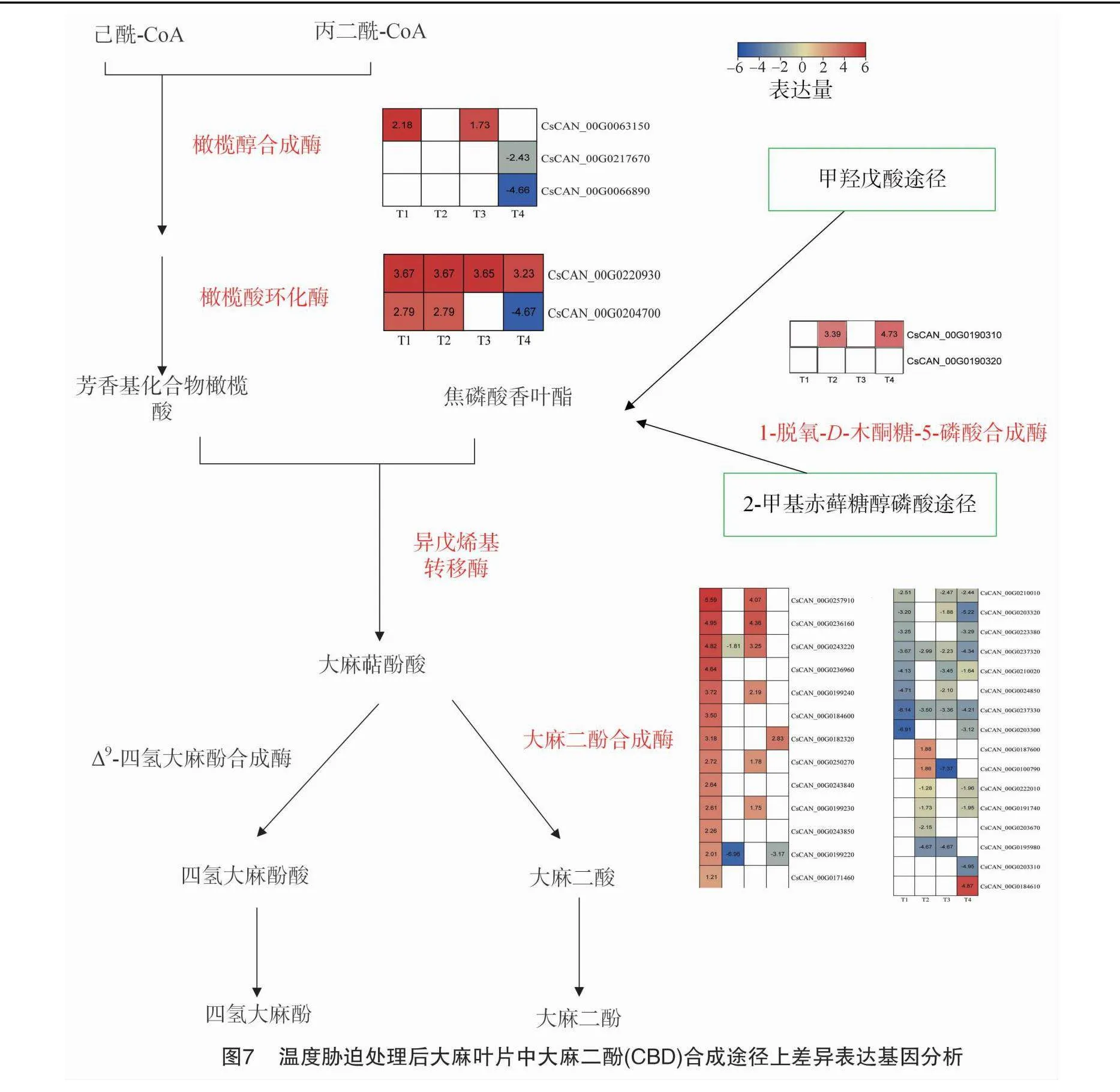
植物大麻素生物合成途徑分為3個催化步驟:(1)聚酮合成(polyketide formation);(2)異戊烯化(prenylation);(3)氧化環(huán)化(oxidative cyclization)。本研究中發(fā)現(xiàn)低溫脅迫也能夠誘導(dǎo)CBD合成途徑相關(guān)基因表達(dá)。橄欖醇合成酶(olivetol synthase,OLS)和橄欖酸環(huán)化酶(olivetolic acid cyclase,OAC)是聚酮合成途徑中的關(guān)鍵酶。在所有的顯著差異表達(dá)基因中,有3個基因被注釋為OLS,為T1-vs-CK(0 ℃/25 ℃)組和T3-vs-CK(15 ℃/25 ℃)組共有的差異基因CsCAN_00G0063150,低溫脅迫后顯著上調(diào)表達(dá)。基因CsCAN_00G0217670和CsCAN_00G0066890在受到低溫脅迫時下調(diào)表達(dá)(圖7)。基因CsCAN_00G0063150,CsCAN_00G0066890和CsCAN_00G0217670 GO富集為BP條目生物合成的過程(GO:0009058,biosynthetic process)。基因CsCAN_00G0063150還參與查爾酮生物合成過程(GO:0009715,chalcone biosynthetic process)和大麻素生物合成過程(GO:1901696,cannabinoid biosynthetic process)。
在所有的顯著差異表達(dá)基因中,1個OAC(CsCAN_00G0220930)為4個溫度處理共有的差異基因,還有另外1個OAC(CsCAN_00G0204700)為T1-vs-CK(0 ℃/25 ℃)組和T2-vs-CK(5 ℃/25 ℃)組共有的差異基因,上述基因在受到低溫脅迫之后均顯著上調(diào)表達(dá)。
大麻戊烯基轉(zhuǎn)移酶1(cannabis sativa prenyltransferase 1,CsPT1)和烯戊烯基轉(zhuǎn)移酶4(cannabis sativa prenyltransferase 4,CsPT4)已被證明在大麻素生物合成途徑中催化產(chǎn)生大麻二酚酸(cannabigerolic acid,CBGA)的步驟,CBGA是產(chǎn)生大麻二酸(CBDA)和四氫大麻酚酸(THCA)的終點酶的底物。在所有的顯著差異表達(dá)基因中,有8個基因被注釋為PT,這些基因中沒有響應(yīng)4個脅迫所共有的差異基因。在T1-vs-CK(0 ℃/25 ℃)組和T3-vs-CK(15 ℃/25 ℃)組溫度脅迫處理下,有5個PT共有的差異基因,除CsCAN_00G0027410之外其他均顯著上調(diào)表達(dá)(圖7)。T4-vs-CK(20 ℃/25 ℃)溫度脅迫處理下,2個PT均下調(diào)表達(dá)。其中CsCAN_00G0202190屬于葉綠素生物合成過程(GO:0015995,chlorophyll biosynthetic process)。
大麻二酚合成酶(cannabidiolic acid synthase,CBDAS)催化CBGA單萜部分氧化環(huán)化,分別轉(zhuǎn)化為大麻二酸(cannabidiolicacid,CBDA)。在所有的顯著差異表達(dá)基因中,有29個基因被注釋為CBDAS。其中21個CBDAS響應(yīng)T1-vs-CK(0 ℃/25 ℃)組溫度脅迫處理,10個CBDAS響應(yīng)T2-vs-CK(5 ℃/25 ℃)組溫度脅迫,14個CBDAS響應(yīng)T3-vs-CK(15 ℃/25 ℃)組溫度脅迫,13個CBDAS響應(yīng)T4-vs-CK(20 ℃/25 ℃)組溫度脅迫。除CsCAN_00G0100790之外,其他28個基因GO富集為BP條目氧化還原過程(GO:0055114//oxidation-reduction process)。
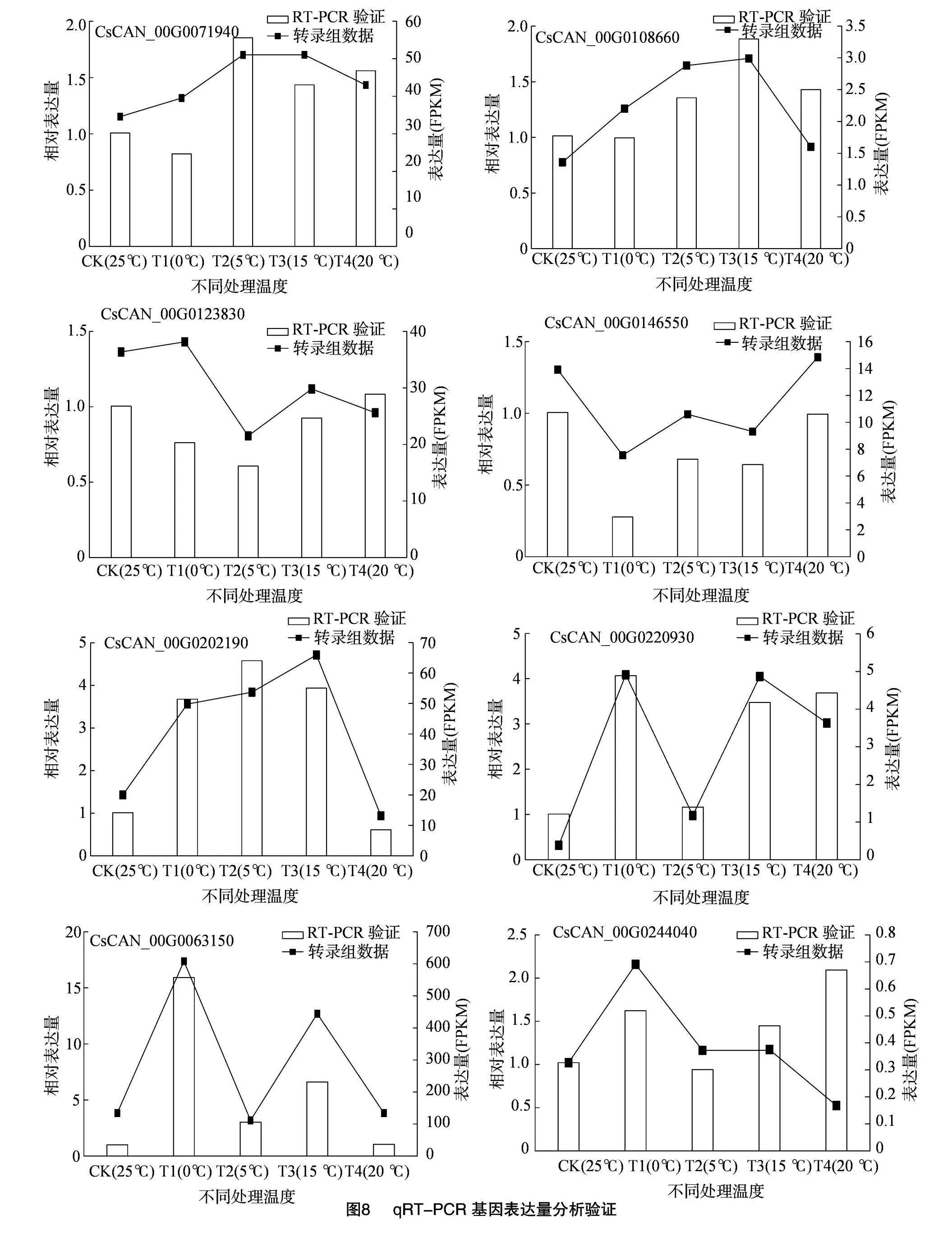
2.2.7 差異基因的qRT-PCR驗證
為了驗證由RNA-seq測序獲得的基因表達(dá)模式的準(zhǔn)確性,通過qRT-PCR檢測了CBD生物合成途徑中10個DEG的表達(dá)模式(圖8)。比較了用qRT-PCR檢測的基因的表達(dá)量在T1(0 ℃/25 ℃)、T2(5 ℃/25 ℃)、T3(15 ℃/25 ℃)和T4(20 ℃/25 ℃)4個比較組之間的相對表達(dá)情況。所選基因的qRT-PCR檢測的相對表達(dá)趨勢與RNA-seq檢測的表達(dá)趨勢一致,說明RNA-seq數(shù)據(jù)是可靠的,重復(fù)性好。
3 討論
首先,本研究探討了不同溫度處理下工業(yè)大麻植株CBD積累的規(guī)律,結(jié)果表明:與對照(25 ℃)相比,T1組(0 ℃)處理后CBD含量最高,表明0 ℃對大麻CBD含量有一定的影響。T2組(5 ℃)、T3組(15 ℃)和T4組(20 ℃)與對照常溫組(25 ℃)相比CBD含量均有所下降,其中T2組(5 ℃)CBD含量最低。隨著處理溫度的升高,從5 ℃到20 ℃,CBD含量緩慢上升,但始終低于常溫對照組。由此可見,0 ℃短時處理后會提高大麻CBD含量,但短時 5~20 ℃之間,短時處理會減低大麻CBD含量。可能與植物體對溫度的適應(yīng)機制有關(guān),在5~20 ℃這個溫度范圍內(nèi),持續(xù)較低溫度使得工業(yè)大麻生長變慢,低溫抑制地上部生長,使得干物質(zhì)積累量減少[10-11]。
其次,試驗研究了低溫脅迫對工業(yè)大麻CBD含量的調(diào)控作用,通過不同溫度處理后對大麻葉片進(jìn)行轉(zhuǎn)錄組分析,發(fā)現(xiàn)在T1-vs-CK組(0 ℃/25 ℃)上調(diào)基因數(shù)量最多,且差異表達(dá)基因上調(diào)數(shù)遠(yuǎn)多于下調(diào)數(shù)。另外對于差異共表達(dá)基因進(jìn)行GO分析發(fā)現(xiàn),三大功能類別中膜(GO:0016020,membrane)、氧化還原酶活性(GO:0016491,oxidoreductase activity)和氧化還原過程(GO:0055114,oxidation-reduction process)富集最多。由于植物受到低溫脅迫時會促進(jìn)細(xì)胞內(nèi)活性氧的產(chǎn)生,推測溫度處理可能影響了大麻細(xì)胞內(nèi)的氧化還原反應(yīng)。植物次生代謝物的合成不僅是一類有用的天然產(chǎn)物,而且是植物抵抗環(huán)境脅迫防御系統(tǒng)的重要組成。轉(zhuǎn)錄組測序檢測到的DEG相關(guān)功能主要集中在膜系統(tǒng)結(jié)構(gòu)、碳水化合物(蔗糖和淀粉)代謝、次生代謝產(chǎn)物合成等代謝通路。本研究對KEGG富集分析發(fā)現(xiàn),植物MAPK信號通路(MAPK signaling pathway -plant)、次級代謝物的生物合成(biosynthesis of secondary metabolites)、代謝通路(metabolic pathways)等通路顯著富集,表明溫度處理影響了次生代謝。Shan等的研究表明,結(jié)縷草在低溫脅迫下(4 ℃)能夠激活植物體內(nèi)氧化應(yīng)急反應(yīng)、MAPK信號通路、類黃酮和異黃酮生物合成和萜類主干物質(zhì)生物合成等次生代謝產(chǎn)物通路富集顯著。
大麻素的前體實際上起源于2種不同的生物合成途徑:聚酮合成(polyketide formation)途徑和非甲羥戊酸途徑(MEP pathway)[12]。橄欖醇合成酶(OLS)和橄欖酸環(huán)化酶(OAC)是聚酮合成途徑中的關(guān)鍵酶,在它們的作用下產(chǎn)生橄欖酸(olivetolic acid,OLA)。脫氧-D-木酮糖-5-磷酸合成酶(DXS)是MEP途徑中的第一個酶,在植物激素調(diào)控、逆境抗性和病原體防御等生理過程中發(fā)揮著重要作用[13],MEP途徑最終合成焦磷酸香葉酯(geranyl diphosphate,GPP)。轉(zhuǎn)錄組數(shù)據(jù)分析發(fā)現(xiàn)有3個差異表達(dá)基因被注釋為OLS基因,2個差異表達(dá)基因被注釋為OAC基因,2個差異表達(dá)基因被注釋為DXS基因,上述7個差異表達(dá)基因在T1-vs-CK組(0 ℃ /25 ℃)均上調(diào)表達(dá),且在4個處理組中表達(dá)量最高。由此可見,T1組(0 ℃)處理促進(jìn)提高OLS、OAC和DXS基因的表達(dá),進(jìn)而提高CBD含量。大麻異戊烯轉(zhuǎn)移酶4(CsPT4)和1(CsPT1)是大麻素生物合成的限速酶,催化GPP和OLA產(chǎn)生大麻二酚酸(CBGA)[14]。轉(zhuǎn)錄組數(shù)據(jù)共有8個差異表達(dá)基因被注釋為PT基因,6個差異表達(dá)基因在 T1-vs-CK組(0 ℃/25 ℃)上調(diào)表達(dá),上調(diào)表達(dá)的基因數(shù)量明顯高于其他處理組。大麻二酚合成酶(CBDAS)催化CBGA單萜部分氧化環(huán)化,轉(zhuǎn)化為大麻二酸(CBDA)。本研究數(shù)據(jù)中共有29個差異表達(dá)基因被注釋為CBDAS基因,13個差異表達(dá)基因在T1-vs-CK組(0 ℃/25 ℃)上調(diào)表達(dá),上調(diào)表達(dá)的基因數(shù)量明顯高于其他處理組。
4 結(jié)論
溫度與植物生長發(fā)育、有機物產(chǎn)量的積累密切相關(guān),不同溫度處理后對大麻CBD的含量和調(diào)控通路產(chǎn)生了不同影響。本研究中,0 ℃能顯著增加工業(yè)大麻葉中CBD含量,其機理可能是通過影響合成通路中OLS、OAC、PT、DXS和CSDAS關(guān)鍵酶基因的表達(dá)進(jìn)而提高CBD含量。
參考文獻(xiàn):
[1]王殿奎,關(guān)鳳芝. 黑龍江省大麻生產(chǎn)現(xiàn)狀及發(fā)展對策[J]. 中國麻業(yè),2005,27(2):98-101.
[2]胡學(xué)禮,郭鴻彥,劉旭云,等. 云南工業(yè)大麻品種在黑龍江大興安嶺地區(qū)的適應(yīng)性研究[J]. 西南農(nóng)業(yè)學(xué)報,2012,25(3):838-841.
[3]郭 麗,王明澤,王殿奎,等. 工業(yè)大麻綜合利用研究進(jìn)展與前景展望[J]. 黑龍江農(nóng)業(yè)科學(xué),2014,8(3):132-134.
[4]王東凱,宮海楠. 工業(yè)大麻的發(fā)展脈絡(luò)及在醫(yī)療領(lǐng)域的應(yīng)用動態(tài)[J]. 黑龍江農(nóng)業(yè)科學(xué),2022,13(22):124-127.
[5]Perez J,Ribera M V. Managing neuropathic pain with Sativex:a review of its pros and cons[J]. Expert Opin Pharmacothe,2008,9(7):1189-1195.
[6]MacCallum C A,Russo E B. Practical considerations in medical cannabis administration and dosing[J]. European Journal of Internal Medicine,2018,49:12-19.
[7]Franco V,Perucca E. Pharmacological and therapeutic properties of cannabidiol for epilepsy[J]. Drugs,2019,79(13):1435-1454.
[8]Seifalian A,Kenyon J,Khullar V. Dysmenorrhoea:can medicinal Cannabis bring new hope for a collective group of women suffering in Pain,Globally[J]. International Journal of Molecular Sciences,2022,23(24):16201.
[9]張 健,唐 露,冉啟凡,等. 植物響應(yīng)低溫脅迫轉(zhuǎn)錄組測序研究進(jìn)展[J]. 分子植物育種,2020,18(6):1849-1866.
[10]Chandra S,Lata H,Khan I A,et al. Temperature response of photosynthesis in different drug and fiber varieties of Cannabis sativa L.[J]. Physiology and Molecular Biology of Plants,2011,17(3):297-303.
[11]王雅妮,曾糧斌,汪洪鷹,等. 溫度對工業(yè)大麻生長及大麻二酚含量的影響[J]. 湖南農(nóng)業(yè)科學(xué),2021(10):27-31.
[12]高 萍,陳宇嫻,柯 崇,等. 新型靶向化合物:植物大麻素的生物合成途徑及研究進(jìn)展[J]. 生物工程學(xué)報,2021,37(6):1968-1985.
[13]Tian S,Wang D,Yang L,et al. A systematic review of 1-deoxy-D-xylulose-5-phosphate synthase in terpenoid biosynthesis in plants[J]. Plant Growth Regulation,2022,96:221-235.
[14]Sands L B,Haiden S R,Ma Y,et al. Hormonal control of promoter activities of Cannabis sativa prenyl transferase 1 and 4 and salicylic acid mediated regulation of cannabinoid biosynthesis[J]. Scientific Reports,2023,13(1):8620.
