婺源藍冠噪鹛種群生存力分析
2024-09-11 00:00:00肖暢程彬彬石金澤黎敏靈劉潔楊軍張微微
野生動物學報 2024年3期
摘 要:為了解藍冠噪鹛(Pterorhinus courtoisi)的滅絕風險,預測未來種群變化趨勢,采用Vortex 10. 5. 0模型對藍冠噪鹛進行種群生存力分析,并模擬不同參數對藍冠噪鹛種群數量的影響,探索制約藍冠噪鹛種群增長的主要因子,為該物種的保護管理提供科學依據。結果顯示:在標準狀態下(考慮近交、災難因素),藍冠噪鹛滅絕概率為73%,內稟增長率(R)為0. 148 0,周期增長率(λ)為1. 159 5,凈生殖率(R0)為1. 504 6,雌性和雄性的平均世代時間(T)為2. 76 a。在9個參數敏感度分析中,性別比例、1歲后雄性死亡率和天敵災害發生頻率3個參數是影響種群數量變化的主要參數。結果表明,在復雜的環境條件下藍冠噪鹛仍有滅絕風險,其保護工作仍面臨較大壓力。控制天敵捕食壓力、提高藍冠噪鹛雄鳥成活率,將有益于藍冠噪鹛種群增長。
關鍵詞:藍冠噪鹛;種群生存力分析;Vortex模型;婺源
中圖分類號:Q958. 1
文獻標識碼:A
文章編號:2310 - 1490(2024)- 03 - 0531 - 11
DOI:10.12375/ysdwxb.20240309
小種群保護是保護生物學中的一個重要問題,因小種群更容易受環境、數量的影響而隨機波動,更易受到遺傳漂變和近交衰退等威脅,最終趨向滅絕[1?2]。對瀕危物種種群動態預測可以加深對其瀕危程度的了解,為該物種的保護和種群管理提供科學依據,常使用的方法包括指導數據收集、分析致危因素、評估人為活動的影響和評估不同管理對策對于物種恢復的作用等[3]。目前,種群生存力分析(population viability analysis,PVA)在小種群保護上已經形成了一套比較完整的理論和方法,是研究瀕危物種保護的最重要的方法之一[4]。通過持續監測研究種群的動態,并運用模型分析收集的數據,可以為PVA研究者提供堅實的基礎,以制定更為科學的保護策略和規劃[5]。Vortex模型是目前使用較廣泛的PVA模型之一,尤其適用于鳥類和獸類,如黃腹角雉(Tragopan caboti)[6]、朱鹮(Nipponia nippon)[7]、獼猴(Macaca mulatta)[8]、中華斑羚(Naemorhedus gri?seus)[9]以及長江江豚(Neophocaena asiaeorientalisasiaeorientalis)[10]等。
藍冠噪鹛(Pterorhinus courtoisi)屬雀形目(Passeriformes)噪鹛科(Leiothrichidae)Pterorhinus 屬[11],IUCN 將其列為極危物種(CR)[12],為國家一級重點保護野生動物[13]。目前,藍冠噪鹛野生種群僅分布在江西婺源及其周邊地區,種群數量在300 只左右[14]。其繁殖地為多個斑塊化闊葉林或孤立古樹,生存狀況岌岌可危[15?16]。對于藍冠噪鹛的研究主要集中在分類[17?19]、繁殖生態[20]、生境選擇及利用[21]、籠養個體的食性[22]、行為節律[23]、疾病[24]、種群遺傳多樣性[25]以及人類活動干擾[26]等方面,尚缺乏對其種群動態的預測研究,這不利于掌握其瀕危狀況及采取相應保護措施。基于此,本研究擬通過Vortex模型對藍冠噪鹛進行種群生存力分析,以期為該物種的保護計劃制定提供科學依據。
1 研究方法
1. 1 研究區概況
婺源位于江西東北部,地處贛、浙、皖三省交界處(29°1′—29°35′ N,117°22′—118°11′ E),地貌以中低山、丘陵為主,全縣森林覆蓋率達82%[27],四季分明,是典型的中亞熱帶東南季風氣候,年平均降水量1 882. 76 mm,其中1—6月降水呈遞增趨勢,7—12月呈遞減趨勢。藍冠噪鹛繁殖期4—7月的總降雨量約占全年的57%[28]。婺源地區鳥類資源十分豐富,共有鳥類63科302種[29],除藍冠噪鹛外,瀕危鳥類還有白腿小隼(Microhierax melanoleucos)、中華秋沙鴨(Mergus squamatus)和小太平鳥(Bombycilla ja?ponica)等[30]。本研究所使用的藍冠噪鹛野外種群數量、繁殖參數等均來源于多年對婺源及其周邊地區分布的藍冠噪鹛繁殖種群的野外調查。
1. 2 種群動態預測
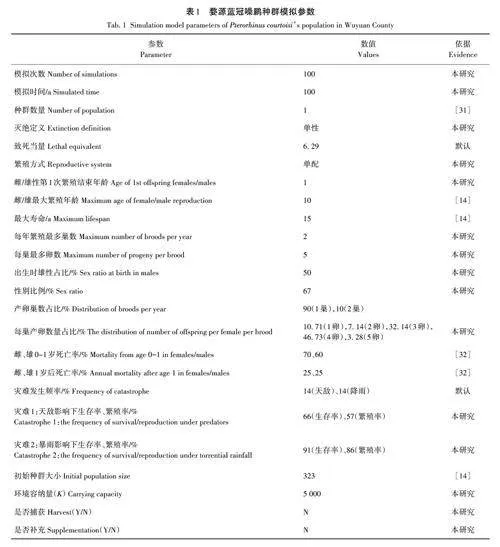
選擇Vortex 10. 5. 0對婺源藍冠噪鹛的種群動態進行預測,模擬期限為100 a,模擬重復次數為100次。繁殖參數來源于2016—2021年藍冠噪鹛野外調查數據,并以相關研究[14,31?32]報道作為模型參數設置的依據(表1)。
1. 2. 1 基本參數
藍冠噪鹛繁殖種群只在婺源地區活動[33],各繁殖地直線距離較近,冬季游蕩但有集群現象[31],該鳥目前在我國境內除江西以外沒有其他發現記錄,所以將種群數設置為1。滅絕定義為當種群中只剩同一性別個體。近親繁殖在小種群中很難避免,近交衰退對于種群數量、繁殖成功率和存活率都有一定影響[34]。Vortex 10. 5. 0軟件能模擬近親繁殖對種群生存力的影響,存在雜種優勢、隱形致死兩個模型,本研究選擇雜種優勢模型,由于尚未見藍冠噪鹛及近緣種近交衰退的研究,因此致死當量選擇系統默認參數6. 29。
1. 2. 2 繁殖參數
藍冠噪鹛雌雄個體間無明顯差異,而單色型鳥種中一夫多妻制較少出現[35],根據野外觀察認為該物種應為單配制。盡管何芬奇等[14]曾記錄到1只雄性藍冠噪鹛與3只雌性相繼交配,但考慮到其不具有普遍性,在種群中應屬偶然性發生,本研究中藍冠噪鹛的婚配制度仍然設置為單配制。
小型鳥類的性成熟年齡為1齡,目前并沒有藍冠噪鹛的最大繁殖年齡研究,藍冠噪鹛野外種群壽命可達10 a以上,圈養種群壽命最高可達25 a[14],設定婺源藍冠噪鹛雌雄個體最大繁殖年齡為10齡,最長壽命為15齡;每年產卵最多巢數為2巢,每巢最多卵數為5枚;初始性比中雄性占比50%。
由于藍冠噪鹛是合作繁殖社會系統,假設在種群非常接近承載能力前,成年繁殖的比例幾乎不會受到密度依賴的影響,因此使用Vortex中的方程式
P(N)={P(0)-[(P(0)-P(K))(N/K)B]}N(/ N+A)。
式中:P(N)是種群數量為N 時繁殖的雌性的百分比,P(K)是種群達到環境承載力(K)時繁殖的百分比,P(0)是不存在Allee效應時低密度繁殖的成年雌性百分比,A 為Allee效應,種群數量極低時尋找配偶難度造成的配對率降低,在藍冠噪鹛種群無此現象時,故取值為0,B 表示P(N)隨著N 的變化強度,設定為2,因為當P(N)為N 的二次函數時,可以更好地模擬密度制約型種群的增長。
1. 2. 3 死亡率
藍冠噪鹛的死亡率未見報道,在野外也很難目擊死亡。因此,本研究中參考Lu 等[32]對斑尾臻雞(Tetrastes sewerzowi)研究中的參數,并結合一處繁殖地育雛期間觀察得到的數據將幼鳥死亡率設為60%,成鳥死亡率設定為25%。
藍冠噪鹛成鳥性比為67%[36],明顯偏雄。Desbiez等[37]提出:成年性別比的偏倚更有可能是死亡率的差異,而不是出生時性別比的偏倚。因此本研究將初始性比設定為50%,并將可配對繁殖雄性占雄性個體的比例作為性比參數。Székely 等[38]的研究結果發現雌性相較于雄性更容易因為性染色體上的有害突變而導致死亡率升高。依此推斷,藍冠噪鹛成鳥性比偏雄可能與雌性相對于雄性更容易遭受疾病影響等導致雌性死亡率更高有關;同時考慮到雛鳥出巢至成鳥的這段時期其死亡率更高,因此將0~1歲雌性死亡率相對于雄性提高了10%,而1歲后的死亡率設定保持相同。
1. 2. 4 災難
藍冠噪鹛繁殖地靠近居民區,受人類活動影響較大,根據多年野外觀察,天敵捕食和強降雨是導致藍冠噪鹛繁殖失敗及死亡的主要因素。藍冠噪鹛繁殖地天敵眾多,包括猛禽、蛇類、松鴉(Garrulusglandarius)、赤腹松鼠(Callosciurus erythraeus)和野貓等,它們會捕食幼鳥甚至成鳥,捕食卵或毀巢等,威脅藍冠噪鹛生存。藍冠噪鹛繁殖期為4—7月,此時婺源正處于汛期,雨水較多,連續降雨容易造成繁殖失敗。本研究選擇了一處易于觀察的繁殖點進行災難調查(包括天敵捕食和暴雨),根據調查結果進行災難參數的設置。考慮到調查時間較短,且2016年至今,藍冠噪鹛每年均因天敵捕食而導致較低的繁殖率和生存率,因此參考Reed等[39]對脊椎動物的研究,將災難發生頻率設為14%。
1. 2. 5 初始種群規模
何芬奇等[14]統計野外藍冠噪鹛種群數量為323只,因此以323只作為種群初始數量,并以成年雄性占比67%平均分配雌雄數量,同時模擬占比為60%和70%時種群的變化。
1. 2. 6 環境容納量
將婺源地區各個藍冠噪鹛繁殖地面積除以歷史上該區域最大的種群數量,取最小值設定為藍冠噪鹛最小家域面積,將藍冠噪鹛繁殖地與最小家域面積相除即得到環境容納量(K),經過計算將環境容納量設為5 000。
1. 2. 7 收獲和補充
Vortex中的“Harvest(收獲)”可以模擬狩獵、撲殺等場景,而“Supplementation(補充)”用以模擬外源個體的遷入,婺源藍冠噪鹛種群孤立,目前沒有人工繁育個體的重引入和其他地區個體遷入情況,所以不考慮“Supplementation”情況。
1. 2. 8 遺傳多樣性
根據程彬彬[40]對藍冠噪鹛遺傳多樣性的研究結果,設定期望雜合度平均值為0. 268。其他參數設置取軟件默認值。
1. 2. 9 敏感度分析
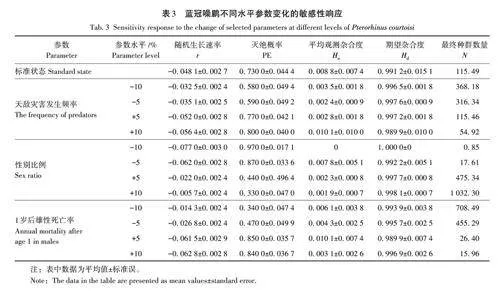
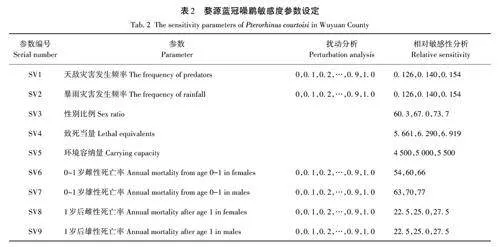
通過擾動分析(perturbation analysis)和相對敏感性分析(relative sensitivity)兩種方法探究藍冠噪鹛種群動態變化對不同死亡率變化的敏感性[41]。其中,擾動分析可以探究最接近種群崩潰的參數,為此在保證其他參數不變的情況下,在0~1范圍內分別將天敵災害發生頻率、暴雨發生頻率、幼鳥死亡率、成鳥死亡率和種群性別比例進行改變,以探究該物種種群大小持久性急劇下降的閾值。不同場景的模擬期限為100 a,模擬重復次數為100,敏感性測試(sensitivity tests,ST)是分析相對敏感性分析的方法之一,可以在保證其他模擬參數一致的情況下,對待測參數進行一定取值范圍內的運行計算,通過不同參數間的對比,來確定哪些參數的變化對模擬物種最為敏感。以標準狀態(考慮近交衰退和災難因素)種群參數作為基礎(表1)進行單因素取樣(singlefactor),在其他參數保持一致的情況下,分別對天敵發生率、暴雨發生率和性別比例等9個參數分別上下波動10%進行模擬,具體參數設置見表2。
同時采用敏感性指數(SX)的計算公式,對死亡率、災難、環境容納量以及性比參數進行獨立模擬分析。具體計算公式為
SX=(λ+-λ-)/0. 2λ0。
式中:λ+和λ-為參數改變后的輸出值;λ0為標準模型的輸出值;0. 2為參數的總變化值(±10%)[41]。
2 結果
2. 1 未來100 年生存動態
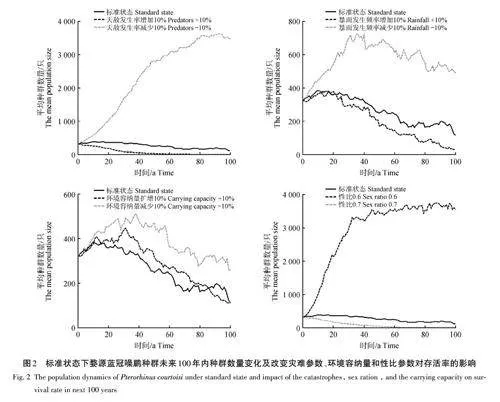
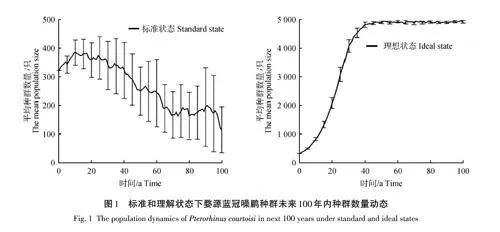
在理想狀態(不考慮近交衰退和災難因素)下,藍冠噪鹛種群數量在40年內迅速增長,從323只達到5 000只左右,其內稟增長率(R)為0. 249 4,基因雜合率(GD)為0. 999 6,種群滅絕概率為0(圖1)。在包含了近交衰退和災難因素的標準狀態時,種群數量在20年內緩慢增長,從323只達到380只左右,之后種群數量呈波動下降狀態,到第100年時,種群數量為115只左右,其內稟增長率為0. 148 0,凈生殖率(R0)為1. 504 6,周期增長率(λ)為1. 159 5,種群滅絕概率為73%,雌性和雄性的平均世代時間(T)為2. 76 a。對近交衰退、災難因素分別與理想狀態進行種群數量上的差異顯著性檢驗(t 檢驗),結果顯示近交衰退對藍冠噪鹛種群數量的影響不顯著 (p=0. 998),而災難對藍冠噪鹛的影響表現為極顯著(plt;0. 001)。
對災難、環境容納量和性比參數的單獨模擬分析結果(圖2)顯示,當天敵災害發生頻率減少10%時,滅絕概率為0,當天敵災害發生頻率增加10%時,滅絕概率為96%;當暴雨災害發生頻率減少10%時,滅絕概率為55%,當暴雨災害發生頻率增加10%時,滅絕概率為75%;當環境容納量擴增10%時,滅絕概率為65%,當環境容納量減少10%時,滅絕概率為81%;當性比為0. 7時,滅絕概率為97%,當性比為0. 6時,滅絕概率為0。說明災害發生頻率越低,環境容納量越大,性比越接近0. 5,藍冠噪鹛種群滅絕概率越小。災難和性比參數變動對藍冠噪鹛種群數量變化影響較大,環境容納量參數變動對藍冠噪鹛種群數量變化影響較小。
2. 2 敏感度分析
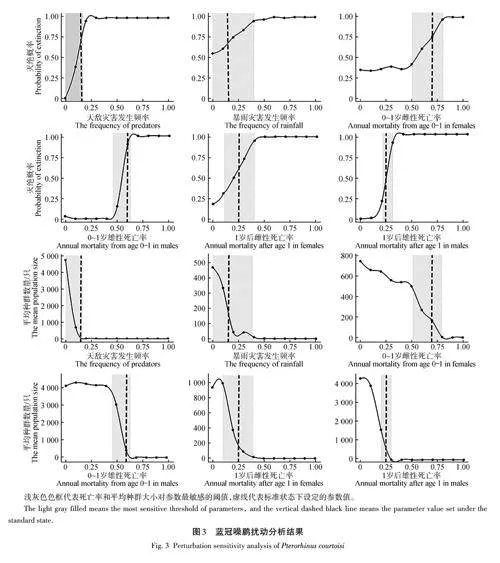
擾動分析結果如圖3所示,滅絕概率和平均種群數量大小對天敵災害發生頻率、0~1歲雄性死亡率和1歲后雄性死亡率更敏感。當天敵災害發生頻率在0~20%時,藍冠噪鹛滅絕概率迅速上升,同時種群數量迅速下降;當天敵災害發生概率在25%左右時,未來100年內藍冠噪鹛的滅絕概率便達到100%左右,平均種群數量下降到0只左右。當0~1歲雄性死亡率在45%~65% 時,藍冠噪鹛滅絕概率迅速上升,同時種群數量迅速下降;當0~1歲雄性死亡率在65%左右時,藍冠噪鹛的滅絕概率達到100%左右,未來100年內平均種群數量下降到0只左右。當1歲后雄性死亡率在15%~35%時,藍冠噪鹛滅絕概率迅速上升,同時種群數量迅速下降;當1歲后雄性死亡率在35%左右時,未來100年內藍冠噪鹛的滅絕概率達到100%左右,平均種群數量下降到0只左右。
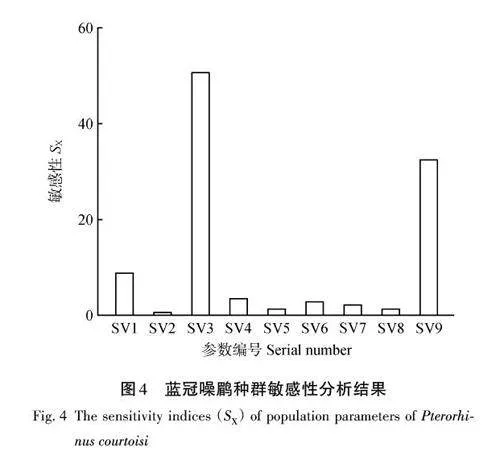
利用SX的絕對值對相對敏感性進行分析(圖4),結果顯示藍冠噪鹛對性別比例參數(SV3)最為敏感,其次為1歲后雄性死亡率(SV9)和天敵災害發生頻率(SV1)。此外,本研究還將最敏感的3種參數在不同水平上進行設置,以進一步探究該物種對不同參數的靈敏程度,結果顯示在此波動范圍內,藍冠噪鹛的隨機生長速率始終是負數,且始終具有較高的滅絕概率(表3)。
2. 3 綜合分析
綜合來看,將影響藍冠噪鹛種群動態變化的因素分為外部因素和內部因素,在外部因素中,由天敵和暴雨構成的災難對種群動態變化占主導地位,其中天敵影響最強,暴雨影響較弱;在內部因素中,性別比例參數最為敏感,其次為1 歲后雄鳥死亡率參數。
3 討論
藍冠噪鹛作為我國特有珍稀鳥類,多年來種群數量增長緩慢。PVA的分析結果表明,性別比例、1歲后雄性死亡率和天敵災害發生頻率是限制種群增長的主要因素。相較于理想狀態,目前標準狀態下藍冠噪鹛種群隨機增長速率均為負數,始終屬于種群數量不穩定的瀕危物種,具有非常小的種群數量,容易受到威脅和隨機事件的影響,因此本研究數據對該瀕危物種的保護具有借鑒意義。
PVA的分析結果表明,藍冠噪鹛生存率對種群存活率影響較大,其中天敵災害發生頻率是影響種群增長的主要因素之一。相較于理想狀態,藍冠噪鹛種群存活率在復雜的環境條件下仍具有滅絕風險。t 檢驗結果顯示災難因素與近交衰退相比,更能顯著影響藍冠噪鹛的種群數量,該結果與捕食、氣候變化是影響鳥類種群數量的主要威脅因素的觀點[42?43]相符。對災難因素的單獨模擬也顯示,降低災難程度對種群存活率影響巨大,說明這是影響藍冠噪鹛種群增長的主要因素之一。
災難相對于其他因素具有更高的不確定性,降雨量的增加會影響鳥類取食食物的質量及數量,降雨導致的溫度變化也會影響其生存率[44]。每年4—7月為藍冠噪鹛的主要繁殖期,這一時期也是婺源的雨季,經常暴發洪水災害,稱為汛期,平均月降雨量在200~400 mm,占全年降水量的57%[28],藍冠噪鹛常因幾天連續降雨而棄巢,孵化較早的雛鳥也會因失溫或親鳥無法喂食而死亡,降低了繁殖成功率。天敵的存在一方面會增加直接死亡率,另一方面會使藍冠噪鹛產生警戒等反捕食行為,增加了繁殖期的能量投入進而影響繁殖。從敏感性分析來看,控制天敵捕食率可以使種群數量隨機生長速率增加。此外,性比也是種群的基本參數之一,是影響種群動態的重要因素,尤其是種群水平的成年性比對種群配偶關系及繁殖潛力有很大影響,它決定有效種群的大小,是種群走向繁榮還是沒落的一個重要指標[45],對于一雄一雌制的動物來說,性比1∶1對種群增長最有利[46],性比偏倚會使有效種群降低,種群存活率下降。本研究中藍冠噪鹛屬于單配制性比偏雄物種,敏感性分析顯示性別偏倚對種群存活率影響最大,這與Lu等[32]對斑尾臻雞研究的情況一致。由此可見,對藍冠噪鹛而言,偏雄的性比對其種群十分不利,其性比偏雄的機制尚需進一步研究。
相對敏感性分析結果顯示,性別比例與種群增長率間存在正反饋現象,性別比例和1歲后雄性死亡率對藍冠噪鹛種群數量的影響最大。結合擾動分析結果可知,在性別偏倚影響下的生存率是影響藍冠噪鹛種群數量變化的主要參數,其次是1歲后雄性死亡率和天敵災害發生頻率。由此可以看出,平衡藍冠噪鹛的性別比例、降低1歲后雄鳥死亡率及提高天敵災害影響下的存活率,將有益于藍冠噪鹛種群數量的提升。朱鹮為單配制鳥類,其配對方式與藍冠噪鹛相同,李海洋[7]模擬不同性比情況下朱鹮群體的增長率,證實性比失衡將會導致種群數量下降,但該研究認為環境容納量是所研究對象的重要致危因素之一,這與本研究中環境容納量對藍冠噪鹛種群存活率變化影響不敏感的結果相反,但與Fantle-lepczyk 等[47]對瀕危鳥類小考島鶇(Myadestespalmeri)的分析結果一致,這可能與鳥類對生境要求的程度及鳥體大小有關。藍冠噪鹛體型小,活動范圍小且對生境沒有特殊要求[21],因此環境容納量沒有成為限制其自身增長的因素。
為了更好地保護藍冠噪鹛這一瀕危程度高的物種,需要針對上述結果制定相應保護管理措施。例如:加強繁殖期內藍冠噪鹛種群動態監測,控制天敵,降低因天敵捕食導致的死亡率;加強繁殖后期的監測和管理,明確其越冬地并提供適當的保護措施以降低非繁殖期成鳥的死亡率。近年來,婺源積極開發旅游業,旅游旺季也恰逢藍冠噪鹛繁殖期,繁殖點人為活動頻繁,觀鳥和拍鳥活動、鄉村基礎設施和旅游設施等工程建設對藍冠噪鹛的干擾日趨嚴重[48?49],部分繁殖點藍冠噪鹛已經暫時性或永久性消失,其種群的生存狀態不容樂觀。由于該物種短期滅絕風險依然很高,亟待開展遷地保護及人工繁育研究,為以后的種群復壯和重引入計劃做好準備。
致謝:感謝江西農業大學黃慧琴、劉濤、謝金生和蘇英鈺等在野外調查數據中提供的幫助!
參考文獻:
[1] FRANKHAM R. Genetics and extinction[J]. Biological Conservation,
2005, 126(2): 131-140.
[2] 覃建庸, 陳名紅, 向左甫. VORTEX模型及其在小種群保護中
的作用[J]. 安徽農業科學, 2008, 36(20): 8783-8785; 8787.
QIN J Y,CHEN M H,XIANG Z F. VORTEX model and its function
in small population protection[J]. Journal of Anhui Agricultural
Sciences, 2008, 36(20): 8783-8785;8787.
[3] BROOK B W, O’GRADY J J, CHAPMAN A P, et al. Predictive
accuracy of population viability analysis in conservation biology
[J]. Nature, 2000, 404: 385-387.
[4] CAUGHLEY G. Directions in conservation biology[J]. Journal of
Animal Ecology, 1994, 63(2): 215-244.
[5] CHAUDHARY V, OLI M K. False dichotomy in population viability
analysis quality assessment: reply to Lawson et al[J]. Conservation
Biology, 2021, 35(5): 1686-1688.
[6] 桂小杰. 黃腹角雉保護生物學研究遺傳多樣性和種群生存力
分析[D]. 長沙:中南林業科技大學, 2007.
GUI X J. The research on the conservation biology of Cabot’s
tragopan genetics diversity and population viability analysis[D].
Changsha: Central South University of Forestry amp; Technology,
2007.
[7] 李海洋. 朱鹮(Nipponia nippon)種群生存力分析初步研究
[D]. 北京:北京林業大學, 2013.
LI H Y. The population viability analysis for Nipponia nippon
[D]. Beijing: Beijing Forestry University, 2013.
[8] 楚原夢冉, 昝啟杰, 楊瓊, 等. 廣東內伶仃島獼猴種群動態及
種群生存力分析[J]. 野生動物學報, 2019, 40(2): 259-266.
CHU Y M R, ZAN Q J, YANG Q, et al. Population dynamic and
viability analysis of rhesus macaque (Macaca mulatta) in Neilingding
Nature Reserve, Guangdong Province[J]. Chinese Journal of
Wildlife, 2019, 40(2): 259-266.
[9] 唐書培, 李春華, 劉威, 等. 內蒙古賽罕烏拉國家級自然保護
區中華斑羚種群生存力分析[J]. 林業科學, 2019, 55(3):
118-124.
TANG S P, LI C H, LIU W, et al. Viability analysis of Chinese
goral population in Saihanwula National Nature Reserve, Inner
Mongolia[J]. Scientia Silvae Sinicae, 2019, 55(3): 118-124.
[10] 吳斌, 王偉萍, 賀剛, 等. 長江江豚種群生存力分析及其對
保護區規劃的啟示[J]. 江西農業學報, 2020, 32(6): 93-98.
WU B, WANG W P, HE G, et al. Analysis of population viability
of Yangtze River finless porpoise and its enlightenment to protected
area planning[J]. Acta Agriculturae Jiangxi, 2020, 32
(6): 93-98.
[11] 鄭光美. 中國鳥類分類與分布名錄[M]. 4版. 北京:科學出
版社, 2023:308.
ZHENG G M. A checklist on the classification and distribution
of the birds of China[M]. 4th ed. Beijing: Science Press,
2023: 308.
[12] BirdLife International. Garrulax courtoisi[J/OL]. The IUCN
Red List of Threatened Species, 2018: e. T22732350A13189076
4[2023-11-16]. http://dx. doi. org/10. 2305/IUCN. UK. 2018-
2. RLTS. T2273 2350A131890764. en.
[13] 國家林業和草原局, 農業農村部. 國家重點保護野生動物名
錄(2021年2月1日修訂)[J]. 野生動物學報, 2021, 42(2):
605-640.
National Forestry and Grassland Administration, Ministry of Agriculture
and Rural Affairs. List of national key protected wild
animals (reversed on February 1, 2021)[J]. Chinese Journal of
Wildlife, 2021, 42(2): 605-640.
[14] 何芬奇, 林劍聲, 聞丞, 等. 婺源靛冠噪鹛生物學初探[J].
動物學雜志, 2017, 52(1): 167-175.
HE F Q, LIN J S, WEN C, et al. Prelim of biology of the bluecrowned
laughingthrush Garrulax courtoisi in Wuyuan of NE Jiangxi,
SE China[J]. Chinese Journal of Zoology, 2017, 52(1):
167-175.
[15] LIU T, XU Y T, XIA C W, et al. Multiple lines of evidence confirm
that the critically endangered blue-crowned laughingthrush
(Garrulax courtoisi) is an independent species[J]. Avian Research,
2022, 13: 100022.
[16] 廖為明, 洪元華, 俞社保, 等. 婺源黃喉噪鹛繁殖生態及其
與村落風水林關系研究[J]. 江西農業大學學報, 2007, 29
(5): 837-841; 850.
LIAO W M, HONG Y H, YU S B, et al. A study on the propagation
habitat of Garrulax galbanus courtoisi and the relationship of
the birds with village forests in Wuyuan, Jiangxi Province[J].
Acta Agriculturae Universitatis Jiangxiensis, 2007, 29(5):
837-841; 850.
[17] LI N, HUANG X L, YAN Q, et al. Save China’s blue-crowned
laughingthrush[J]. Science, 2021, 373(6551): 171.
[18] 何芬奇, 楊嵐. 黃喉噪鹛分類地位新議[J]. 動物學雜志,
2006, 41(5): 127.
HE F Q, YANG L. A new discussion on the taxonomic status of
the Garrulax galbanus courtosis[J]. Chinese Journal of Zoology,
2006, 41(5): 127.
[19] 劉濤. 藍冠噪鹛分類地位及其系統演化研究[D]. 南昌:江西
農業大學, 2019.
LIU T. Research on the taxonomy and phylogeny of the bluecrowned
laughingthrush (Garrulax courtoisi)[D]. Nanchang: Jiangxi
Agricultural University, 2019.
[20] 黃慧琴, 劉濤, 石金澤, 等. 藍冠噪鹛繁殖期生境選擇特征
分析[J]. 生態學報, 2018, 38(2): 493-501.
HUANG H Q, LIU T, SHI J Z, et al. Habitat selection of the
blue-crowned laughingthrush during the breeding season[J].
Acta Ecologica Sinica, 2018, 38(2): 493-501.
[21] LIU T, XU Y T, MO B, et al. Home range size and habitat use
of the blue-crowned laughingthrush during the breeding season
[J]. PeerJ, 2020, 8: e8785.
[22] 石金澤, 吳志勇, 劉濤, 等. 籠養藍冠噪鹛的食性研究[J].
野生動物學報, 2017, 38(4): 617-621.
SHI J Z, WU Z Y, LIU T, et al. Feeding habits of captive bluecrowned
laughingthrush (Garrulax courtoisi)[J]. Chinese Journal
of Wildlife, 2017, 38(4): 617-621.
[23] 劉濤, 吳志勇, 劉斌, 等. 籠養藍冠噪鹛行為節律和時間分
配研究[J]. 四川動物, 2017, 36(6): 674-679.
LIU T, WU Z Y, LIU B, et al. Study on the activity rhythm and
time budget of captive Garrulax courtoisi[J]. Sichuan Journal of
Zoology, 2017, 36(6): 674-679.
[24] BARBóN A R, LóPEZ J, JAMRI?KA J, et al. Clinical and
pathological aspects of systemic Isospora infection in bluecrowned
laughing thrushes (Garrulax courtoisi) at Jersey Zoo
[J]. Journal of Avian Medicine and Surgery, 2019, 33(3):
265-277.
[25] CHEN G L, ZHENG C Q, WAN N, et al. Low genetic diversity
in captive populations of the critically endangered blue-crowned
laughingthrush (Garrulax courtoisi) revealed by a panel of novel
microsatellites[J]. PeerJ, 2019, 7: e6643.
[26] ZHANG W W, SHI J Z, HUANG H Q, et al. The impact of disturbance
from photographers on the blue-crowned laughingthrush
(Garrulax courtoisi)[J]. Avian Conservation and Ecology,
2017, 12(1): 15.
[27] 汪磊,黃志強,翟健程,等. 旅游干擾對婺源風水林植物多樣
性的影響[J]. 湖北農業科學, 2023, 62(5): 100-106.
WANG L, HUANG Z Q, ZHAI J C, et al. Effects of tourism disturbances
on plant diversity of geomantic forests in Wuyuan[J].
Hubei Agricultural Sciences, 2023, 62(5): 100-106.
[28] WheatA 小麥芽:農業氣象大數據[DB/OL]. (2021-08-10)
[2023-11-16]. http://www. wheata. cn/.
WheatA: agrometeorological big data sharing platform[DB/OL].
(2021-08-10)[2023-11-16]. http://www. wheata. cn/.
[29] 何芬奇, 林劍聲, 王英永, 等. 婺源鳥類紀錄解析[J]. 動物
學雜志, 2014, 49(2): 170-184.
HE F Q, LIN J S, WANG Y Y, et al. Bird records from
Wuyuan, NE Jiangxi of SE China[J]. Chinese Journal of Zoology,
2014, 49(2): 170-184.
[30] 李言闊, 鄭忠杰, 查奇智, 等. 林鳥世界[J]. 森林與人類,
2018(11): 120-133.
LI Y K, ZHENG Z J, ZHA Q Z, et al. World of forest birds[J].
Forest amp; Humankind, 2018(11): 120-133.
[31] 黃慧琴. 藍冠噪鹛(Garrulax courtoisi)繁殖期生境選擇及其影
響因素研究[D]. 哈爾濱:東北林業大學, 2016.
HUANG H Q. Study on habitat selection and influence factors of
the blue-crowned laughingthrush (Garrulax courtoisi)[D]. Harbin:
Northeast Forestry University, 2016.
[32] LU N, SUN Y H. Population viability analysis and conservation
of Chinese grouse Bonasa sewerzowi in Lianhuashan Nature Reserve,
north-west China[J]. Bird Conservation International,
2011, 21(1): 49-58.
[33] 何芬奇, 奚志農. 婺源黃喉噪鹛(Garrulax galbanus courtoisi)
[J]. 動物學雜志, 2002, 37(5): 82.
HE F Q, XI Z N. Garrulax galbanus courtoisi in Wuyuan[J].
Chinese Journal of Zoology, 2002,37(5): 82.
[34] SACCHERI I, KUUSSAARI M, KANKARE M, et al. Inbreeding
and extinction in a butterfly metapopulation[J]. Nature,
1998, 392: 491-494.
[35] IRWIN R E. The evolution of plumage dichromatism in the new
world blackbirds: social selection on female brightness[J]. The
American Naturalist, 1994, 144(6): 890-907.
[36] 莫柏. 藍冠噪鹛微衛星開發及其在遺傳多樣性及親緣關系鑒
定中的應用[D]. 南昌:江西農業大學, 2020.
MO B. Microsatellite development and its application in genetic
diversity and genetic relationship identification for the bluecrowned
laughingthrush (Garrulax courtoisi)[D]. Nanchang: Jiangxi
Agricultural University, 2020.
[37] DESBIEZ A L J, BERTASSONI A, TRAYLOR-HOLZER K.
Population viability analysis as a tool for giant anteater conservation
[J]. Perspectives in Ecology and Conservation, 2020, 18
(2): 124-131.
[38] SZéKELY T, LIKER A, FRECKLETON R P, et al. Sex-biased
survival predicts adult sex ratio variation in wild birds[J]. Proceedings
of the Royal Society B: Biological Sciences, 2014, 281
(1788): 20140342.
[39] REED D H, O’GRADY J J, BALLOU J D, et al. The frequency
and severity of catastrophic die-offs in vertebrates[J]. Animal
Conservation, 2003, 6(2): 109-114.
[40] 程彬彬. 基于重測序技術藍冠噪鹛種內遺傳結構及其遺傳多
樣性研究[D]. 南昌:江西農業大學, 2022.
CHENG B B. Genetic structure and diversity of Garrulax cour?
toisi based on resequencing technology[D]. Nanchang: Jiangxi
Agricultural University, 2022.
[41] MORTENSEN J L, REED J M. Population viability and vital
rate sensitivity of an endangered avian cooperative breeder, the
white-breasted thrasher (Ramphocinclus brachyurus)[J]. PLoS
One, 2016, 11(2): e0148928.
[42] HASEBE M, AOTSUKA M, TERASAWA T, et al. Status and
conservation of the common murre Uria aalge breeding on Teuri
Island, Hokkaido[J]. Ornithological Science, 2012, 11(1):
29-38.
[43] FERNáNDEZ-OLALLA M, MARTíNEZ-ABRAíN A, CANUT
J, et al. Assessing different management scenarios to reverse the
declining trend of a relict capercaillie population: a modelling
approach within an adaptive management framework[J]. Biological
Conservation, 2012, 148(1): 79-87.
[44] LI C. A review of the relationship between cooperative breeding
and climate in birds[J]. Open Journal of Nature Science, 2019,
7(3): 170-174.
[45] 霍雅鵬, 萬冬梅. 鳥類性別鑒定的方法及性比研究的進展
[J]. 遼寧大學學報(自然科學版), 2008, 35(4): 358-362.
HUO Y P, WAN D M. Development of methods of sex identification
and studies on sex ratio in birds[J]. Journal of Liaoning University(
Natural Sciences Edition), 2008, 35(4): 358-362.
[46] 孫儒泳. 動物生態學原理[M]. 4版. 北京:北京師范大學出
版社,2019:84.
SUN R Y. Principles of animal ecology[M]. 4th ed. Beijing:
Beijing Normal University Press, 2019: 84.
[47] FANTLE-LEPCZYK J, TAYLOR A, DUFFY D C, et al. Using
population viability analysis to evaluate management activities for
an endangered Hawaiian endemic, the Puaiohi (Myadestes palm?
eri)[J]. PLoS One, 2018, 13(6): e0198952.
[48] 李梁平, 謝曉文. 婺源縣農戶景觀油菜種植制約因素的對策
探討[J]. 時代農機, 2015, 42(8): 109-111.
LI L P, XIE X W. Wuyuan County farmers the countermeasures
for the rape planting landscape factors[J]. Times Agricultural
Machinery, 2015, 42(8): 109-111.
[49] 石培華, 馮凌, 唐曉云, 等. 建設中國美麗鄉村 世界生態文
化公園:解讀“婺源之路”[J]. 今日國土, 2008(12): 39-42.
SHI P H, FENG L, TANG X Y, et al. Construction of China
beautiful countryside world ecological cultural park: interpretation
of“ Wuyuan Road”[J]. China Territory Today, 2008(12):
39-42.
