蘆筍超雄株花粉母細(xì)胞減數(shù)分裂觀察
2024-08-24 00:00:00李霞牛曉雪包艷存路遠(yuǎn)李付軍牟萌雷霆田洪平李保華楊林
中國瓜菜 2024年7期
摘" " 要:減數(shù)分裂是有性生殖的關(guān)鍵過程。為了明確蘆筍超雄株花粉母細(xì)胞減數(shù)分裂的具體情況及異常行為對配子生長發(fā)育的影響,采用改良苯酚品紅壓片法對58株蘆筍超雄株的986個(gè)花粉母細(xì)胞減數(shù)分裂過程進(jìn)行了系統(tǒng)觀察。結(jié)果表明,(1)超雄株花粉母細(xì)胞減數(shù)分裂為胞質(zhì)分裂同時(shí)型,染色體構(gòu)型為10個(gè)二價(jià)體,四分體為十字交叉型。(2)中期Ⅰ小型棒狀二價(jià)體的出現(xiàn)比例為96.55%,四價(jià)體為8.62%,染色體粘連為22.41%。(3)后期Ⅰ落后染色體的出現(xiàn)比例為12.07%,染色體橋?yàn)?3.79%;分裂期出現(xiàn)微核的比例為6.59%。(4)在中期Ⅱ~末期Ⅱ中分裂不同步的出現(xiàn)比例為34.45%。超雄株花粉母細(xì)胞在整個(gè)減數(shù)分裂進(jìn)程中,前期Ⅰ所需時(shí)間最長。減數(shù)分裂配對成10個(gè)二價(jià)體,在倍性水平上屬于二倍體。減數(shù)分裂異常行為主要出現(xiàn)在中期Ⅰ和后期Ⅰ。對超雄株花粉母細(xì)胞減數(shù)分裂過程的觀察,不僅能夠豐富蘆筍的細(xì)胞遺傳學(xué)內(nèi)容,還可以為蘆筍發(fā)育生物學(xué)、雜交育種、基因定位等方面提供必要的細(xì)胞遺傳學(xué)信息。
關(guān)鍵詞:蘆筍;超雄株;花粉母細(xì)胞;減數(shù)分裂
中圖分類號:S644.6 文獻(xiàn)標(biāo)志碼:A 文章編號:1673-2871(2024)07-095-05
Observation on meiosis of pollen mother cells of super-male plants in Asparagus officinalis L.
LI Xia, NIU Xiaoxue, BAO Yancun, LU Yuan, LI Fujun, MU Meng, LEI Ting, TIAN Hongping, LI Baohua, YANG Lin
(Weifang Academy of Agricultural Sciences, Weifang 261071, Shandong, China)
Abstract: Meiosis is a key process in sexual reproduction. In order to clarify the specific situation and the effect of abnormal meiotic behavior of pollen mother cells(PMCs) on the growth and development of gametes in super -male asparagus plants, the improved phenol fuchsin method was used to systematically observe the meiosis process of 986 PMCs of 58 super-male asparagus plants. The results showed that:(1)The PMCs of super-male plants carried meiosis and cytokinesis simultaneously, with a chromosome configuration of 10 bivalents and the tetrad of a cross shaped structure.(2)At metaphase I, the proportion of small rod bivalents was 96.55%, tetravalent was 8.62%, and chromosome sticking was 22.41%.(3)At anaphase I, the proportion of lagging chromosomes was 12.07%, chromosome bridges was 13.79%, and the proportion of micronucleus was 6.59% in the division phase.(4)At metaphase II to telophase II, the proportion of asynchronization of chromosomes segregation was 34.45%. In the whole process of meiosis, the prophase Ⅰ is the longest. There was paired as 10 bivalents at meiosis metaphase Ⅰ,thus it is a diploid species.The abnormal behavior of meiosis mainly occurs in metaphase I and anaphase I. Based on the observation of the meiosis process of PMCs of the super-male, it can not only enrich the cytogenetic content of asparagus, but also provide the necessary cytogenetic information for asparagus of developmental biology, cross breeding, gene localization, and so on.
Key words: Asparagus officinal L.; Super-male plants; Pollen mother cells; Meiosis
減數(shù)分裂是DNA復(fù)制一次,而細(xì)胞連續(xù)分裂兩次,形成單倍體的精子或卵子,減數(shù)分裂過程中非同源染色體及非姊妹染色單體間發(fā)生交換,使配子的遺傳多樣化,增加了后代的適應(yīng)性。減數(shù)分裂是維持物種染色體數(shù)目穩(wěn)定,使生物物種得以保存延續(xù)的重要生物過程,在植物生命周期中維持遺傳穩(wěn)定性和豐富變異性方面起重要作用[1]。在減數(shù)分裂過程中會出現(xiàn)正常二價(jià)體(指在減數(shù)分裂偶線期同源染色體聯(lián)會時(shí),正常是每對同源染色體形成一個(gè)二價(jià)體,此時(shí),每條染色體由兩條姊妹染色單體組成。有n對染色體的細(xì)胞中將形成n個(gè)二價(jià)體)、四分體(指花粉母細(xì)胞在孢子母細(xì)胞的分化方向中,在花粉形成前,四個(gè)花粉由胼胝質(zhì)相連形成的結(jié)構(gòu))。當(dāng)然也會出現(xiàn)異常分裂行為:(1)單價(jià)體:減數(shù)分裂過程中不能配對的染色體。一般在花粉母細(xì)胞減數(shù)分裂前期Ⅰ和中期Ⅰ出現(xiàn)。(2)四價(jià)體:在減數(shù)分裂偶線期發(fā)生聯(lián)會的四條同源染色體配成一組的多價(jià)體。(3)染色體橋:由兩條染色體分別發(fā)生斷裂后,具有著絲粒的殘臂相接而形成的雙著絲粒染色體所特有的結(jié)構(gòu),在兩極之間被拉緊而形成橋。(4)落后染色體:在減數(shù)分裂的后期I、末期I、后期Ⅱ、末期Ⅱ,大多數(shù)染色體移向兩極之后,少數(shù)染色體不移向兩極,仍停滯在赤道板上或者在移向兩極過程中與大多數(shù)染色體不同步,形成落后的染色體或斷片。(5)微核:隨著落后染色體或斷片的出現(xiàn)及分裂的進(jìn)行,這些滯后的染色體或斷片丟失或游離于主核外,形成微核[1]。
蘆筍(Asparagus officinalis L.)為百合科天門冬屬多年生草本植物,具有2n =2x = 20條染色體,二倍體基因組大小為1323 Mb [2-4],幼莖質(zhì)嫩味美,營養(yǎng)豐富,風(fēng)味獨(dú)特,是一種具有較高藥用、營養(yǎng)和保健價(jià)值的高檔蔬菜,暢銷全球,享有“蔬菜之王”的美譽(yù)[5-6]。蘆筍為典型的雌雄異株植物,具有XY型性別決定系統(tǒng),蘆筍的自然群體中性別表現(xiàn)型有4種,分別為雌性、雄性、超雄性和兩性株,性染色體組成分別為XX,XY,YY和XY[7]。超雄株(YY)可以正常生長發(fā)育,形態(tài)及生理等各方面特征均與正常雄株(XY)相同,但其與雌株雜交的后代全為雄性,因此超雄株對蘆筍育種具有重要意義[8-10]。目前生產(chǎn)上獲得超雄株的方法主要有兩種,一是花粉培養(yǎng)誘導(dǎo)蘆筍雄核形成雙單倍體,二是雄性兩性花自花或異花授粉[11-12]。國內(nèi)對蘆筍超雄株的研究主要涉及超雄株的獲得、田間鑒定篩選、分子鑒定篩選等方面[13-16],還未涉及蘆筍超雄株染色體的研究,因此,筆者采用常規(guī)的細(xì)胞學(xué)分析方法對蘆筍超雄株花粉母細(xì)胞進(jìn)行減數(shù)分裂觀察,旨在為蘆筍超雄株的研究增加細(xì)胞學(xué)資料,為進(jìn)一步揭示超雄株生長發(fā)育及全雄育種提供理論基礎(chǔ)。
1 材料與方法
1.1 材料
2018年在濰坊市農(nóng)業(yè)科學(xué)院現(xiàn)代農(nóng)業(yè)科技示范園定植968株蘆筍雄性兩性株,從中鑒定出了58株超雄株[15],筆者以58株超雄株單株為試驗(yàn)材料,2023年5—6月在超雄株的盛花期不間斷地取長度0.5 cm左右的花蕾進(jìn)行觀察。
1.2 方法
采用改良的苯酚品紅染色法,取超雄株0.5 cm左右的花蕾,在卡諾氏固定液Ⅰ中固定24 h,然后轉(zhuǎn)移到70%酒精中于4 ℃冰箱保存?zhèn)溆谩2捎脡浩ㄖ破⌒迈r花蕾或從70%酒精中取出保存的花蕾,用鑷子取出花藥,放置于載玻片中央,滴加1滴苯酚品紅染液,用鑷子和解剖針剖開花藥,將其中的小孢子母細(xì)胞擠出,去除雜質(zhì),蓋上蓋玻片并壓片,用解剖針輕輕敲擊并排除氣泡,在酒精燈外焰烤片3~4 s或者手背感覺微熱時(shí),在N-300M生物顯微鏡下100倍觀察并拍照。
2 結(jié)果與分析
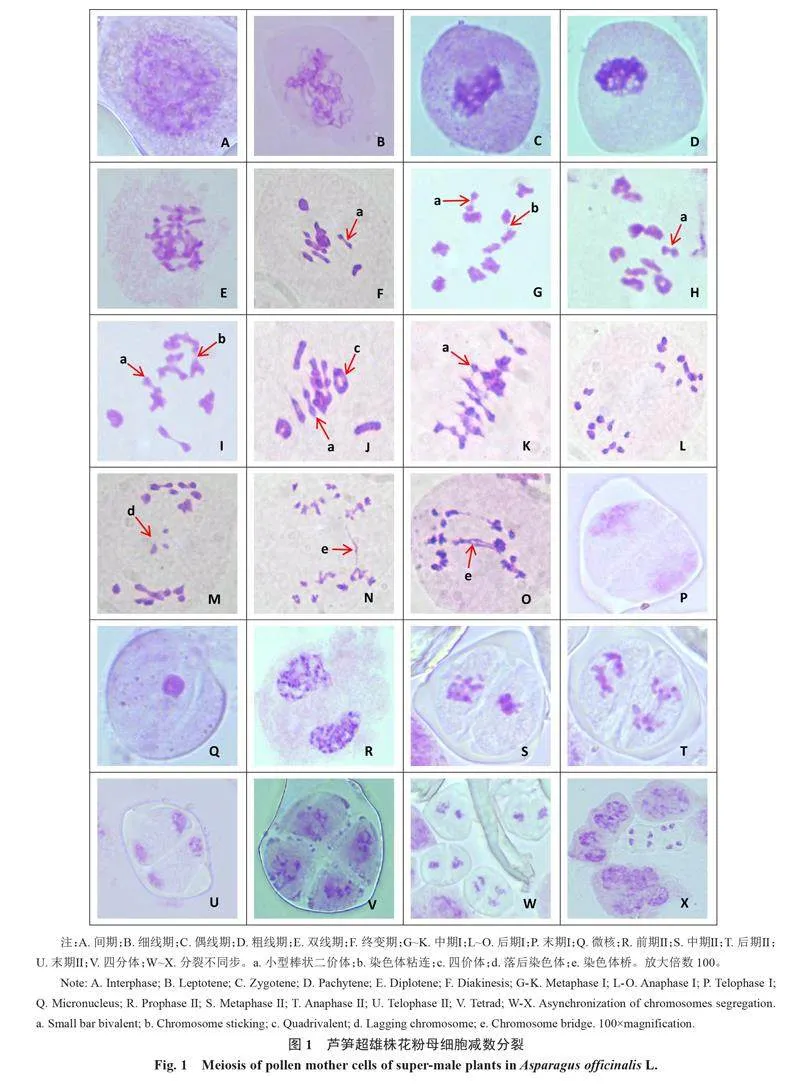
筆者對58株超雄株的986個(gè)花粉細(xì)胞減數(shù)分裂進(jìn)行觀察,發(fā)現(xiàn)減數(shù)分裂過程包括2次連續(xù)的核分離,第一次減數(shù)是同源染色體的分離,第二次減數(shù)是姊妹染色單體的分離。細(xì)胞胞質(zhì)分裂方式為同時(shí)型,詳細(xì)分裂過程如圖1所示。
2.1 間期
染色體呈松散的染色質(zhì)狀態(tài),核內(nèi)染色較為均勻,核質(zhì)分界明顯(圖1-A)。
2.2 前期Ⅰ-細(xì)線期
花粉母細(xì)胞的核仁開始解體,著色能力越來越弱。染色質(zhì)濃縮為幾條細(xì)而長的線,首尾不分地繞成一團(tuán),每一染色體已復(fù)制為2個(gè)單體(圖1-B)。
2.3 前期Ⅰ-偶線期
染色體形態(tài)與細(xì)線期沒有太大的變化,同源染色體開始配對,出現(xiàn)聯(lián)會現(xiàn)象,染色體松散,增粗,但無法辨認(rèn)染色單體(圖1-C)。
2.4 前期Ⅰ-粗線期
同源染色體配對完畢,出現(xiàn)二價(jià)體,每個(gè)二價(jià)體含有4個(gè)染色單體,但僅含有2個(gè)著絲粒,染色體繼續(xù)縮短變粗、螺旋化,向四周發(fā)散(圖1-D)。
2.5 前期Ⅰ-雙線期
染色體進(jìn)一步縮短變粗,同源染色體分離,此時(shí)可以看到染色體的交叉現(xiàn)象,因?yàn)榻徊媸沟脝误w上的若干點(diǎn)仍然連在一起,出現(xiàn)U形、8字形等多種形狀(圖1-E)。
2.6 前期Ⅰ-終變期
交叉向染色體端部移動,交叉數(shù)減少,染色體變得更粗。到終變晚期,染色體與中期染色體大小基本相同,核仁著色變淺,此時(shí)可見10對同源染色體構(gòu)成10個(gè)二價(jià)體,呈環(huán)狀、棒狀(圖1-F)。
2.7 中期Ⅰ
側(cè)面觀察染色體整齊地排列在赤道板上,極面觀察染色體散布在細(xì)胞中,兩兩配對分散在細(xì)胞質(zhì)中。在58株超雄株的所有花粉母細(xì)胞的減數(shù)分裂過程中,單株之間不存在明顯的差異,染色體構(gòu)型均為10個(gè)二價(jià)體,即10Ⅱ,二價(jià)體構(gòu)型的形狀有棒狀、環(huán)狀,以環(huán)狀為主,棒狀二價(jià)體1~5個(gè)不等(圖1-G~K);在分裂過程中,小型棒狀二價(jià)體的出現(xiàn)比例為96.55%、四價(jià)體為8.62%、染色體粘連為22.41%。
2.8 后期Ⅰ
同源染色體彼此分開,移向細(xì)胞兩極。由于此時(shí)著絲點(diǎn)未分開,所以細(xì)胞兩極的染色體數(shù)是母細(xì)胞的一半,但每條染色體中依然含有兩條單體(圖1-L~O)。在分裂過程中落后染色體的出現(xiàn)比例為12.07%;染色體橋?yàn)?3.79%。
2.9 末期Ⅰ
同源染色體分別到達(dá)兩極,染色體變成了染色質(zhì)狀,核膜、核仁重新出現(xiàn),形成2個(gè)子核,此時(shí)每個(gè)子核染色體數(shù)目減半為10條,同時(shí)細(xì)胞質(zhì)分開形成2個(gè)子細(xì)胞(二分體),可確定超雄株的細(xì)胞質(zhì)分裂方式為同時(shí)型(圖1-P)。在分裂期微核的出現(xiàn)比例為6.59%(圖1-Q)。
2.10 前期Ⅱ
染色體縮短,前期快結(jié)束時(shí)核膜消失,此時(shí)的每條染色體都有2條單體,細(xì)胞有10條染色體(圖1-R)。
2.11 中期Ⅱ
染色體濃縮變短,2組染色體分別排列在各自的赤道板上,紡錘體出現(xiàn),每條染色體有2個(gè)染色單體和1個(gè)著絲粒(圖 1-S)。
2.12 后期Ⅱ
著絲點(diǎn)完成復(fù)制,彼此分開。姊妹染色單體從著絲粒處分裂為二,在紡錘絲的作用下分別移向兩極(圖 1-T)。
2.13 末期Ⅱ
移到兩極的染色體解螺旋,濃縮成團(tuán),出現(xiàn)核仁、核膜,形成單倍的子核(圖 1-U),胞質(zhì)分裂,1個(gè)母細(xì)胞分裂成 4個(gè)子細(xì)胞,至此減數(shù)分裂結(jié)束,進(jìn)入四分體時(shí)期(圖 1-V)。在中期Ⅱ~末期Ⅱ中分裂不同步的出現(xiàn)比例為34.45%(圖 1-W~X)。
3 討論與結(jié)論
國內(nèi)外鮮見對蘆筍超雄株花粉母細(xì)胞減數(shù)分裂的研究。筆者在本研究中發(fā)現(xiàn),超雄株花粉母細(xì)胞的減數(shù)分裂過程符合二倍體減數(shù)分裂的規(guī)律。減數(shù)分裂的染色體行為,在不同植株或相同植株的花蕾上表現(xiàn)基本一致;超雄株花粉母細(xì)胞是胞質(zhì)同時(shí)型;染色體構(gòu)型為10個(gè)二價(jià)體,有棒狀和環(huán)狀,四分體為十字交叉型。絕大多數(shù)花粉母細(xì)胞減數(shù)分裂中染色體的行為正常,但也有不同比例的四價(jià)體、染色體粘連、落后染色體、染色體橋、微核、分裂不同步等現(xiàn)象發(fā)生。在自然條件下,外界因素或個(gè)體細(xì)胞內(nèi)某些生理、生化過程可誘發(fā)染色體結(jié)構(gòu)變異。中期Ⅰ四價(jià)體的出現(xiàn)可能導(dǎo)致一部分細(xì)胞中的同源染色體不能準(zhǔn)確地分配到兩極,從而造成染色體的重復(fù)或缺失;落后染色體很可能是四價(jià)體在后期分離產(chǎn)生的,最終形成微核或消失在細(xì)胞質(zhì)中從而導(dǎo)致配子染色體數(shù)不平衡,從而影響超雄株的育性[17]。同源染色體之間出現(xiàn)的一定程度異質(zhì)性導(dǎo)致染色體橋的出現(xiàn),這可以作為鑒定是否出現(xiàn)染色體倒位的證據(jù)之一[18-19]。在第二次減數(shù)分裂過程中發(fā)現(xiàn)1~3個(gè)時(shí)期分裂同時(shí)出現(xiàn)在1個(gè)花蕾中,這種不同步性可以擴(kuò)展花期,在發(fā)育過程中增加兩性功能的有效作用時(shí)間,對種群繁殖后代有利,是一種適應(yīng)當(dāng)?shù)丨h(huán)境條件的進(jìn)化表現(xiàn)[20-21]。
筆者首次對蘆筍超雄株的減數(shù)分裂過程進(jìn)行了較為系統(tǒng)的觀察研究,結(jié)果表明,超雄株花粉母細(xì)胞經(jīng)過2次分裂,最終形成4個(gè)子細(xì)胞。超雄株花粉母細(xì)胞在整個(gè)減數(shù)分裂過程中,前期Ⅰ所需時(shí)間最長。減數(shù)分裂配對成10個(gè)二價(jià)體,在倍性水平上屬于二倍體。減數(shù)分裂異常行為主要出現(xiàn)在中期Ⅰ和后期Ⅰ。在減數(shù)分裂中期Ⅰ有單價(jià)體、四價(jià)體的產(chǎn)生,后期Ⅰ、后期Ⅱ出現(xiàn)落后染色體、染色體橋、微核等,從而導(dǎo)致小孢子發(fā)育成無育性的配子,進(jìn)而產(chǎn)生無活力的花粉,降低了花粉的育性。在花粉母細(xì)胞第二次分裂過程中出現(xiàn)的染色體不同步現(xiàn)象,在一定程度上延長了花期,可降低外界極端環(huán)境對散粉或減數(shù)分裂的影響而增加授粉概率,提高了種群繁殖能力,是一種生物適應(yīng)環(huán)境的進(jìn)化機(jī)制。通過對超雄株花粉母細(xì)胞減數(shù)分裂過程的觀察,有助于了解其花粉形成的過程和超雄性的起源與進(jìn)化。研究結(jié)果不僅能夠豐富蘆筍的細(xì)胞遺傳學(xué)內(nèi)容,還可以為蘆筍發(fā)育生物學(xué)、雜交育種、基因定位等方面提供必要的細(xì)胞遺傳學(xué)信息,為全雄育種和遺傳改良奠定基礎(chǔ)。隨著雙色熒光原位雜交技術(shù)、高通量測序技術(shù)、DAPI分帶技術(shù)等分子細(xì)胞遺傳學(xué)各種研究手段的不斷改進(jìn),染色體研究的準(zhǔn)確率有了較大提高,染色體研究將在物種的染色體組分析、物種的起源與演化分析、外源染色體導(dǎo)入片段的鑒定、染色體精細(xì)結(jié)構(gòu)分析等方面有更廣闊的應(yīng)用前景。
參考文獻(xiàn)
[1] 劉大鈞.細(xì)胞遺傳學(xué)[M].北京:中國農(nóng)業(yè)出版社,1999.
[2] 任青.蘆筍簡介[J].現(xiàn)代農(nóng)業(yè),1988(5):9-10.
[3] 李可峰.石刁柏(Asparagus officinalis L.)種質(zhì)資源遺傳多樣性分析[D].濟(jì)南:山東師范大學(xué),2004.
[4] HARKESS A,ZHOU J S,XU C Y,et al.The asparagus genome sheds light on the origin and evolution of a young Y chromosome[J].Nature Communications,2017,8:1279.
[5] 高武軍,張宏杰,王孝娟,等.石刁柏EST序列中微衛(wèi)星分布特征分析[J].生物技術(shù),2011,21(6):27-31.
[6] 郭鑫,趙傳志,趙術(shù)珍,等.蘆筍的雜交育種與生物技術(shù)研究進(jìn)展[J].分子植物育種,2021,19(23):7910-7916.
[7] 周勁松,湯泳萍,盛文濤,等.蘆筍性別決定與性別分化研究進(jìn)展[J].植物遺傳資源學(xué)報(bào),2010,11(5):600-604.
[8] L?PTIEN H.Identification of the sex chromosome pair in asparagus(Asparagus officinalis L.)[J].Zeitschrift Für Pflanzenziichtung,1979,82(2):162-173.
[9] 黃玲,賴佳,韋樹谷,等.石刁柏兩性花與雄花的抗氧化酶活性及其相關(guān)基因可變剪接的差異分析[J].植物生理學(xué)報(bào),2019,55(8):1231-1238.
[10] 李霞,包艷存,李保華,等.蘆筍兩性株染色體的細(xì)胞學(xué)研究[J].種子,2022,41(7):88-91.
[11] 周維燕.蘆筍組織培養(yǎng)及育種中的應(yīng)用[M].北京:高等教育出版社,1989.
[12] 劉奇頎,瞿華香,余格輝,等.蘆筍種質(zhì)資源與育種技術(shù)研究進(jìn)展[J].中國農(nóng)學(xué)通報(bào),2021,37(1):55-60.
[13] 李樂斌,張景麗,馮光,等.蘆筍性別基因型的分子鑒定新技術(shù)與超雄株篩選[J].中國蔬菜,2022(8):78-84.
[14] 周勁松,湯泳萍,羅紹春,等.蘆筍超雄株的DNA分子標(biāo)記輔助篩選[J].園藝學(xué)報(bào),2012,39(11):2182-2188.
[15] 李霞,高武軍,牛曉雪,等.蘆筍超雄株STS雙分子標(biāo)記的開發(fā)及應(yīng)用[J].植物遺傳資源學(xué)報(bào),2022,23(6):1828-1834.
[16] 李芳,張?jiān)獓G存,等.蘆筍全雄系同質(zhì)型父本“超雄株”的選育[J].山東農(nóng)業(yè)科學(xué),2006(2):40-41.
[17] 張蜀寧,萬雙粉,張偉,等.同源四倍體青花菜花粉母細(xì)胞的減數(shù)分裂[J].園藝學(xué)報(bào),2007,34(2):387-390.
[18] 陳欣,楊倩,張平冬,等.胡楊小孢子母細(xì)胞減數(shù)分裂與花粉變異研究[J].西北林學(xué)院學(xué)報(bào),2015,30(2):90-95.
[19] 何麗霞.黃牡丹花粉母細(xì)胞減數(shù)分裂過程的細(xì)胞遺傳學(xué)觀察[J].蘭州大學(xué)學(xué)報(bào),2004,40(6):78-82.
[20] WYATT R.Inflorescence architecture:How flowe rnumber,arrangement,and phenology affect pollnation and fruit-set[J].American Journal of Botany,1982,69(4):585-594.
[21] HARDER L D,THOMSON J D.Evolutionary options for maximizing pollen dispersal of animal-pollinated plants[J].American Naturalist,1989,133(3):323-344.
收稿日期:2024-01-09;修回日期:2024-03-28
基金項(xiàng)目:山東省重點(diǎn)研發(fā)計(jì)劃項(xiàng)目(2022LZGC024);濰坊市科技發(fā)展計(jì)劃項(xiàng)目(2022ZJ1084,2021ZJ1128);濰坊市現(xiàn)代種業(yè)科研創(chuàng)新團(tuán)隊(duì)項(xiàng)目
作者簡介:李" " 霞,女,研究員,研究方向?yàn)樘J筍遺傳育種及栽培。E-mail:55654687@qq.com
通信作者:李保華,男,研究員,研究方向?yàn)樘J筍育種與栽培及推廣。E-mail:shidaiyi@126.com
楊" " 林,男,高級農(nóng)藝師,研究方向?yàn)槭卟擞N與栽培。E-mail:80408239@qq.com
