闊葉紅松林中黑啄木鳥的取食生境偏好
2024-06-29 00:00:00于美辰陳俊達司雨蕙魏爽荊媛戎可
野生動物學報 2024年2期
摘 要 2020年冬季—2022年冬季,采用樣線法和樣方法對黑龍江省涼水國家級自然保護區中黑啄木鳥(Dryocopus martius)取食生境偏好進行調查研究,結果表明:在取食微生境尺度中,黑啄木鳥取食樹樹種、取食樹狀態與對照樣方具有顯著差異(plt;0. 05),偏好在冷杉(Abies fabri,47. 71%)和半枯立木(73. 86%)的樹木主干(86. 93%)中上部(43. 14%)、上部(32. 03%)取食,且偏好粗壯、高大的樹木;在取食樣方尺度中,黑啄木鳥偏好在有林窗(92. 16%)的天然林(96. 73%)中取食,偏好郁閉度較低、優勢喬木均高較低、優勢喬木平均胸徑較大、樣方中活立木較少、枯立木、倒木較多和距人為干擾更近的生境中取食,與對照樣方呈顯著差異(plt;0. 05)。資源選擇函數結果表明,影響黑啄木鳥取食生境偏好的關鍵因子是取食樹樹種,次關鍵因子是枯立木數量、優勢喬木均高、取食樹高度和倒木數量。
關鍵詞:黑啄木鳥;取食生境偏好;闊葉紅松林;資源選擇函數
中圖分類號:Q958. 1 文獻標識碼:A 文章編號:2310 - 1490(2024)- 02 - 0328 - 10
DOI:10.12375/ysdwxb.20240212
取食行為是指動物依靠外界食物獲取供其生存所需的營養,包括搜尋食物、獲取食物和處理食物等過程[1]。行為生態學家在20世紀70年代初就用優化模型探究覓食策略,用最優覓食理論解釋上述問題[2]。鳥類的取食行為研究是進行鳥類生態學研究的重要內容之一,取食行為不僅在不同種之間存在差異,在性別甚至個體間也都有所差別,可以通過取食行為探尋鳥類與環境之間的關系,揭示鳥類生存的原理[3]。
啄木鳥作為重要的森林物種,主要以棲息在樹皮表面或亞表面的昆蟲為食,在某些情況下,啄木鳥捕食樹木害蟲可使林分尺度中的蛀木昆蟲減少98%[4]。啄木鳥被認為是生態系統工程師和傘護種,它們在樹上挖掘的巢穴,為多種鳥類、哺乳動物和無脊椎動物提供庇護所和繁殖地[5?6],森林中多種植物、鳥類和哺乳動物的豐度也與啄木鳥的存在、物種豐富度呈正相關[7?10]。黑啄木鳥(Dryocopus martius)作為大型初級鑿洞者的關鍵物種,在森林生態系統中發揮著重要的生態作用[11]。黑啄木鳥的種群數量與所在地區森林生態系統特征間具顯著聯系,森林結構對黑啄木鳥的分布也有影響[12]。較大的體型使黑啄木鳥在取食、繁殖及棲息中需要依賴一定的林木尺度,然而目前國內對于黑啄木鳥取食生境偏好的研究較少。本研究于2020年冬季—2022年冬季在涼水國家級自然保護區的闊葉紅松林中開展,利用樣線法、樣方法對黑啄木鳥取食生境進行野外調查,探究其取食樹微生境偏好及取食樣方特征,為黑啄木鳥的保護和管理提供科學依據,對于理解森林生態系統的演替規律和制定森林生態系統的保護對策具有重要意義。
1 研究區概況
黑龍江省涼水國家級自然保護區位于伊春市帶嶺區(47°6'—47°16' N,128°47'—128°57' E),行政區屬伊春市大箐山縣[13],保護區總面積12 133 hm2,核心區面積3 740 hm2,其中原始紅松(Pinus koraien?sis)林面積約為2 375 hm2。保護區森林覆蓋率達98%[14],近年來林型正從原始林向過熟林演替,站桿數量逐漸增多,主要以白樺(Betula platyphylla)林、椴樹(Tilia spp.)、紅松林、紅皮云杉(Picea koraien?sis)林、冷杉(Abies fabri)、云杉(Picea spp.)林構成天然森林型[15]。野生動物區系組成較為豐富,鳥類共有16目46科252種,以古北界種類居多,占保護區鳥類99%[16]。
2 研究方法
2. 1 數據收集
于2020年冬季—2022年冬季按季度采用樣線法和樣方法進行野外調查。黑啄木鳥一般存在于保護區的闊葉紅松林中,根據黑啄木鳥的求偶鳴叫聲、筑巢行為、取食行為或其他行為等線索來確定黑啄木鳥的取食位置,發現后利用GPS定位,待黑啄木鳥離開,以取食地為中心設置取食生境調查樣方(半徑為10 m的圓形區域),測量并記錄取食生境因子(表1)。在距離調查樣方500 m附近,未發現黑啄木鳥取食及其取食痕跡的區域,隨機設置同等大小的對照樣方,記錄相應生境因子。
2. 2 數據分析
共選取6 個影響黑啄木鳥取食微生境水平偏好,12 個取食樣方水平偏好,共計18 個生境因子。使用卡方檢驗對其中的非數值型生境因子進行差異性分析。由于樣本量≥50,對數值型生境因子使用單個樣本Kolmogorov-Smirnov檢驗,pgt;0. 05表示生境因子呈正態分布,采用獨立樣本t 檢驗;plt;0. 05表示生境因子不符合正態分布,采用2個獨立樣本的U 檢驗(Mann-Whitney U Test)比較差異。
對18 個生境因子使用主成分分析法選取特征值gt;1 且累計貢獻率大于80% 的數個主成分,提取各主成分中載荷絕對值較大的數個生境因子,定義為“綜合因子集合”。采用Spearman秩相關檢驗對非正態分布的生境因子進行相互關系分析,絕對值≥0. 5 的因子經生態學意義分析后進行保留或剔除[17]。將剩余的生境因子標準化后進行二元Logis?tic 回歸建模,根據資源選擇函數模型分析不同生境因子對黑啄木鳥取食行為的影響[18?19]。使用ROC曲線評價資源選擇函數分析的準確性[20],評價標準:曲線下面積的值在0. 5~0. 6,為不及格;≥0. 6~0. 7,較差;≥0. 7~0. 8,一般;≥0. 8~0. 9,良好;≥0. 9~1. 0,優秀[21]。數據統計分析均利用R 4. 2. 1軟件完成。
3 結果
3. 1 黑啄木鳥取食微生境尺度生境因子
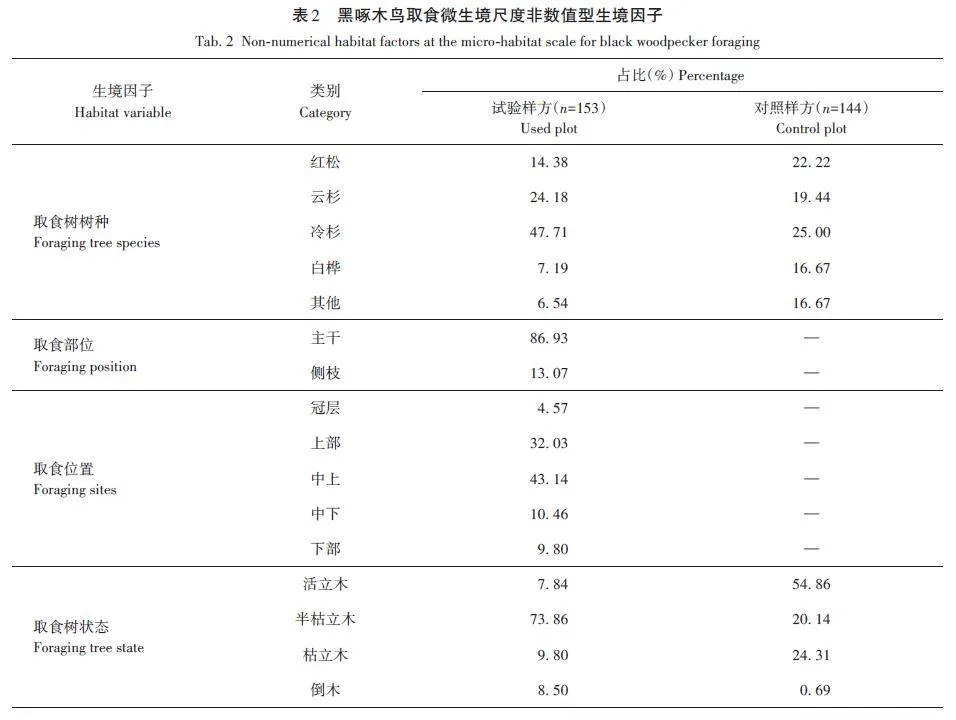
研究共記錄黑啄木鳥取食生境樣方153個,對照樣方144個。對黑啄木鳥取食微生境的4個非數值型生境因子進行卡方檢驗(表2),得出黑啄木鳥取食所在樹種最多為冷杉(47. 71%),其后依次為云杉(Picea asperata,24. 18%)、紅松(14. 38%)、白樺(7. 19%)及其他樹種(6. 53%),與對照樣方樹種差異極顯著(χ2=26. 002,df=4,plt;0. 001);多在主干(86. 93%)部位取食;偏好在取食樹的中上(43. 14%)及上部(32. 03%)位置取食;多選擇樹狀態為半枯立木(73. 86%)作為取食樹,與對照樣方差異極顯著(χ2=117. 140,df=3,plt;0. 001)。
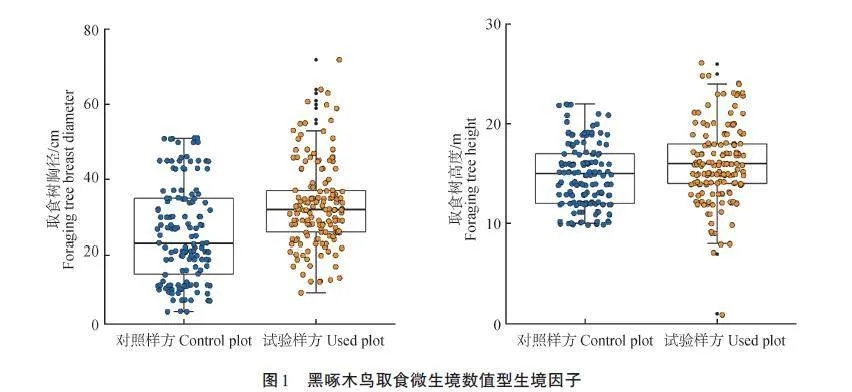
對黑啄木鳥取食微生境尺度中的2個數值型生境因子與對照樣方進行比較發現,黑啄木鳥偏好胸徑較大[(33. 19±12. 09)cm,Z=-3. 590,plt;0. 001]、高度較高[(16. 03±4. 08)m,Z=-2. 944,plt;0. 01]的樹木進行取食(圖1)。
3. 2 黑啄木鳥取食樣方生境因子
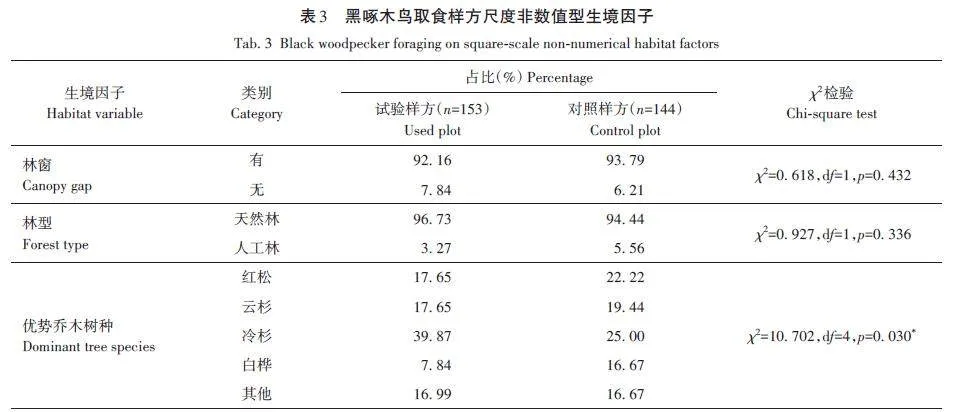
對黑啄木鳥取食樣方中3個非數值型生境因子進行卡方檢驗得出(表3),黑啄木鳥多偏好在有林窗(92. 16%)的天然林(96. 73%)取食,樣方中優勢喬木樹種多為冷杉(39. 87%),樣方優勢喬木與對照樣方存在顯著差異(p=0. 030)。
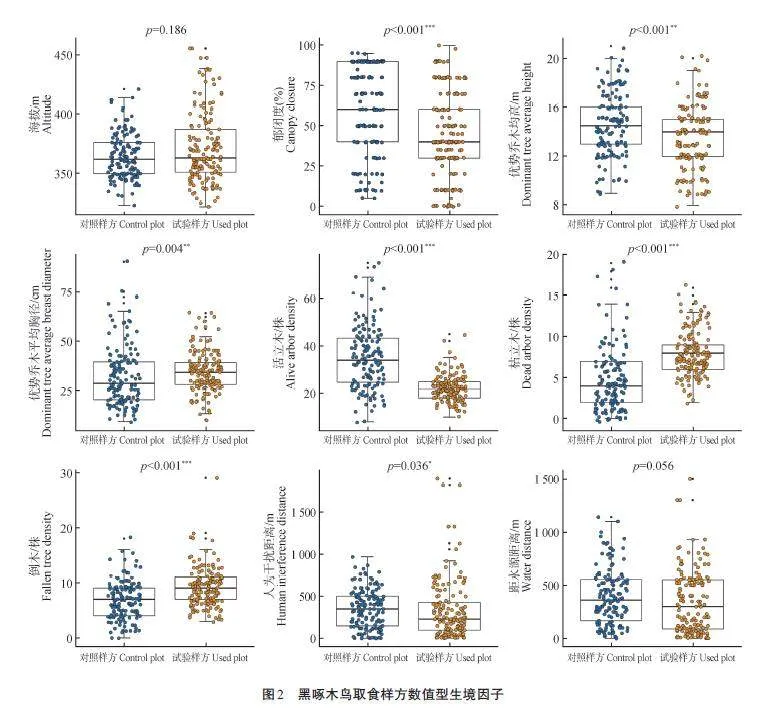
對黑啄木鳥取食樣方尺度中的9個數值型生境因子與對照樣方進行比較發現,黑啄木鳥所取食樣方中對郁閉度、優勢喬木均高、優勢喬木平均胸徑、活立木數量、枯立木數量、倒木數量和人為干擾距離的偏好與對照樣方差異顯著(plt;0. 05)。與對照樣方相比,其偏好在郁閉度較低[(43. 82±25. 68)%]、活立木較少[(22. 00±5. 59)株]、枯立木較多[(8. 00±2. 75)株]、倒木較多[(9. 10±3. 58)株]、距人為干擾距離更近[(334. 09±346. 86)m]的生境取食。其次,對于樣方中優勢喬木的高度及胸徑與對照樣方相比呈現出較大差異,黑啄木鳥取食樣方生境中優勢喬木較低[(13. 51±2. 61)m]且平均胸徑較大[(34. 22±10. 27)cm]。取食生境中的海拔和距水源距離與對照樣方差異均不明顯(圖2)。
3. 3 影響黑啄木鳥取食生境偏好的生境因子
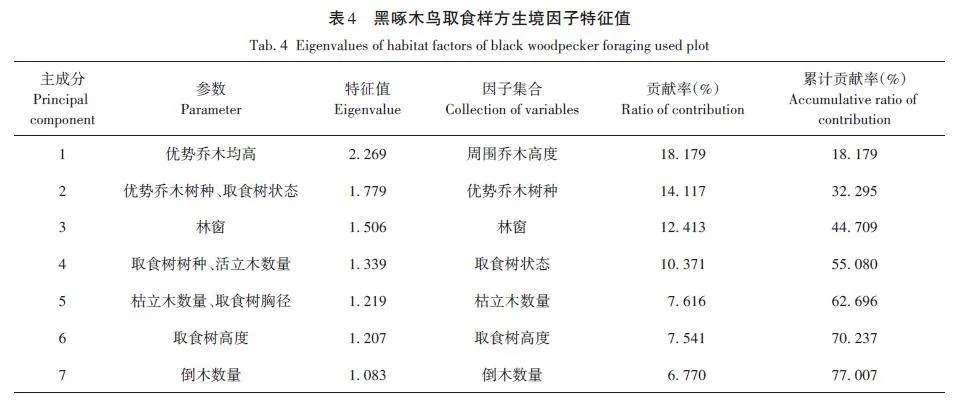
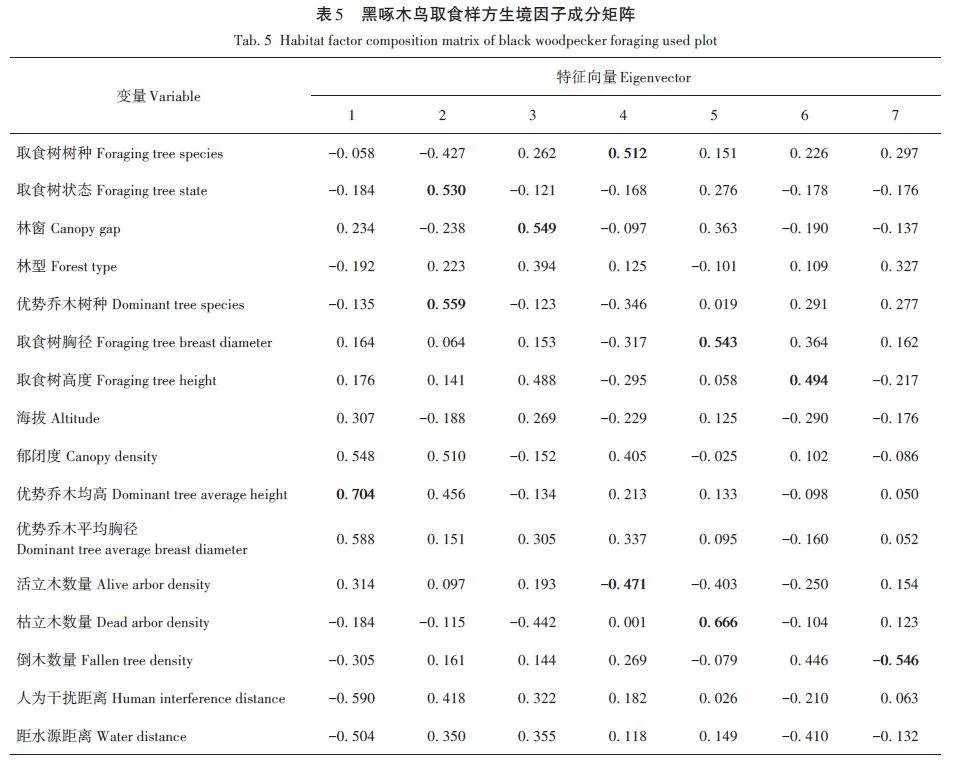
對黑啄木鳥取食生境偏好的18個因子除取食部位和取食位置的16個生境因子進行主成分分析,前7個主成分特征值累計貢獻率達77. 007%(表4),能較好地反映黑啄木鳥取食生境偏好的特征,因此選擇前7個主成分進行分析(表5)。
第一主成分特征值為2. 269,貢獻率為18. 179%,其中載荷絕對值最大的生境因子為優勢喬木均高。第二主成分中載荷絕對值最大的生境因子為優勢喬木樹種、取食樹狀態。第三主成分中載荷絕對值最大的生境因子為林窗。第四主成分中載荷絕對值最大的生境因子為取食樹樹種、活立木數量。第五主成分中載荷絕對值最大的生境因子為枯立木數量、取食樹胸徑。第六主成分中載荷絕對值最大的生境因子為取食樹高度。第七主成分中載荷絕對值最大的生境因子為倒木數量(表4)。
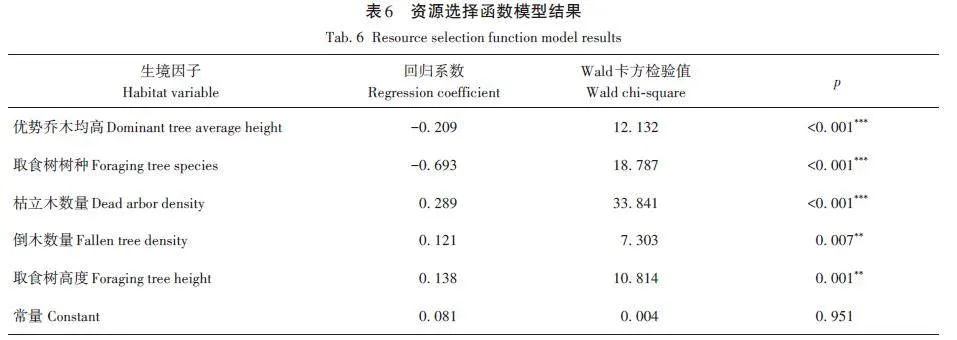
將優勢喬木均高、優勢喬木樹種、取食樹狀態、林窗、取食樹樹種、活立木數量、枯立木數量、取食樹胸徑、取食樹高度和倒木數量10個生境因子經標準化后進行Logistic回歸建模。優化后的Logistic回歸模型顯示,由優勢喬木均高、取食樹樹種、枯立木數量、倒木數量和取食樹高度5個生境因子組成的模型擬合效果最佳,即z=0. 081-0. 209 優勢喬木均高-0. 693取食樹樹種+0. 289枯立木數量+0. 121倒木數量+0. 138取食樹高度(表6),模型預測正確率達78. 1%。
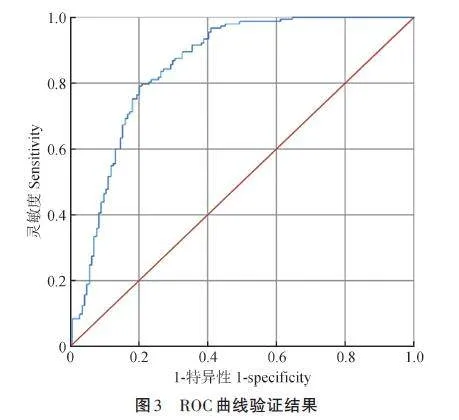
應用ROC曲線法驗證模型穩定性,將Logistic回歸模型中對取食樣方的預測值作為待檢驗變量,將黑啄木鳥是否在試驗樣方取食設置為變量,導出ROC曲線(圖3),曲線下面積為0. 852,高于假設面積0. 5,證實該Logistic 回歸模型具有較好的擬合效果。
4 討論
4. 1 黑啄木鳥取食生境偏好
涼水自然保護區中林木物種豐富,與對照樣方相比,黑啄木鳥更多地在冷杉、云杉和紅松上取食,且多在半枯立木上取食。這說明黑啄木鳥對取食樹樹種有所偏好,在本研究中更多地選擇杉樹取食,Pirovano等[22]在黑啄木鳥取食樹樹種的研究中得出,銀杉(Cathaya argyrophylla)和云杉比歐洲赤松(Pinus sylvestris)和石松(Lycopodium japonicum)斑塊更受歡迎,黑啄木鳥明顯回避后者。Nappi等[23]通過對啄木鳥的取食行為觀察,發現大多數啄木鳥在取食時都會使用枯木基質,更偏好選擇瀕死樹木或站桿。相關研究也得出同樣結論,半枯立木和枯立木是啄木鳥洞穴挖掘的主要資源,為啄木鳥提供覓食、棲息和筑巢等場所[24]。黑啄木鳥多取食于樹木主干的上部和中上部,這可能是因為多種啄木鳥之間發生了生態位分離。戎可等[25]對涼水自然保護區中同域分布的3種啄木鳥進行取食研究,發現黑啄木鳥基本只在主干上鑿洞,三趾啄木鳥(Picoides tridacty?lus)和大斑啄木鳥取食位置多在側枝上,因此啄木鳥取食位置和部位可能會受體型大小及同域分布的其他鳥類的存在而產生差異。同時,黑啄木鳥偏好選擇樹干較粗(胸徑gt;30 cm)且更為高大的樹木取食,這與Gunn 等[26]在緬甸北部和Bocca 等[27]在阿爾卑斯山對黑啄木鳥的取食樹特征研究得出的結果一致,這可能是因為該尺度樹木上昆蟲的密度更高,且胸徑越大的樹木表面積越大,更易獲取食物。同時高大的樹木更容易觀察到天敵的存在,有利于黑啄木鳥在取食時觀察天敵避免被捕食。
黑啄木鳥取食樣方尺度的數值型生境因子與對照樣方相比,海拔和水源距離差異不明顯,這是因為涼水國家級自然保護區內海拔分布均勻,沒有極大高度差,且區內面積較小,隨機樣方與水源距離相差較小。Karimi等[28]在對伊朗北部森林中的黑啄木鳥棲息地利用的調查中得出了不同結果,認為海拔對黑啄木鳥的覓食和棲息地選擇有所影響,不同季節間黑啄木鳥偏好的海拔有所差異。
相比對照樣方,黑啄木鳥顯著偏好在郁閉度更低、周圍優勢喬木較少的區域尋找食物,加之其偏好在高度較高的取食樹取食,進一步說明其在取食時會選擇足夠安全、視線相對較好的環境,這更有利于發現天敵及時做出躲避,以及更快地發現下一處取食區域,提高取食效率。經調查發現,在保護區內單位面積中活立木數量與枯立木、倒木數量呈負相關,這與黑啄木鳥在樣方尺度上取食的選擇結果相同,黑啄木鳥偏好在枯立木和倒木較多而活立木較少的區域尋找食物。有研究認為,黑啄木鳥的存在與胸徑小于10 cm的灌木密度和胸徑為10~29 cm的喬木數量呈負相關[29],進一步證明黑啄木鳥不喜歡植被稀疏的未成熟林,不喜歡無冠層的林木、濃密的灌木及小直徑林木的斑塊。Pirovano等[22]認為黑啄木鳥取食地點的選擇與枯木數量呈正相關,與下層覆蓋度呈負相關,它們對森林的偏好特征是下層植被稀少,可能是因為更容易發現食物資源。但是并非所有啄木鳥都有這一偏好,Stachura-Skierczyńska等[30]在大規模的研究中發現中斑啄木鳥(Leiopicus me?dius)的出現和豐度與障礙物(枯立木和倒木)的密度之間不存在正相關關系。
4. 2 影響黑啄木鳥取食的主要生境因子
資源選擇函數結果表明,影響黑啄木鳥的5個主要生境因子為樣方優勢喬木均高、取食樹樹種、枯立木數量、倒木數量和取食樹高度,其中影響黑啄木鳥取食生境偏好的關鍵因子是取食樹樹種,次關鍵因子是枯立木數量、優勢喬木均高、取食樹高度和倒木數量。通過ROC曲線驗證得出模型正確率大于0. 8,表明模型較好地預測了黑啄木鳥的取食偏好生境。這一結果幾乎與全球大多數黑啄木鳥取食偏好生境研究結果相似。例如:Khanaposhtani等[29]在伊朗探索黑啄木鳥棲息地生境偏好中得出,黑啄木鳥更偏好森林演替的后期階段,即具有成熟林特征,這也與本研究區域相符合,即在多倒木、多枯立木斑塊中的高大粗壯的樹木上棲息、取食。Pirovano等[22]和Zahner等[31]也在阿爾卑斯山脈和德國南部對黑啄木鳥與森林結構中環境變量的相關性研究中發現,大胸徑和高樹木是黑啄木鳥選擇筑巢地點的重要環境因子,并且成熟的林木是其選擇覓食的頻繁區域,同時得出山毛櫸(Fagus longipetiolata)是黑啄木鳥在其研究地進行覓食、繁殖及筑巢的重要樹種,特別是山毛櫸和冷杉、山毛櫸和松樹的混合林,這與本研究中黑啄木鳥偏好的取食生境和樹種具有相同的特點。
黑啄木鳥作為古北界最大的啄木鳥,探究其在森林生態系統中的繁殖和覓食偏好生境,可以作為評估原始森林生態作用的重要指標[32]。在涼水自然保護區調查中,筆者發現毛耳飛鼠(Belomys pearso?nii)、鴛鴦(Aix galericulata)等次級巢穴利用者會在黑啄木鳥取食和繁殖后的區域覓食和棲息,因此對于該地區的啄木鳥保護和管理尤為重要。一般來說,黑啄木鳥依賴于成熟高大的林分、枯立木及倒木,這些都作為黑啄木鳥主要的食物來源和潛在的繁殖地。涼水保護區正處于從原始林向過熟林演替的關鍵階段,探究黑啄木鳥的取食和繁殖生境偏好對保護區內重點物種的生物多樣性保護及棲息生境管理至關重要。建議保護區規劃不同樹齡、樹種以及不同腐爛程度的枯木,為處在不同生態位中的啄木鳥提供穩定、合適的覓食樹,合理管理森林,控制人為干擾,定期對黑啄木鳥種群動態進行監測。
參考文獻:
[1] 羅旭, 艾懷森, 韓聯憲. 高黎貢山白尾梢虹雉取食行為及春季取食地特征[J]. 西南林學院學報, 2010, 30(6): 64-67.
LUO X, AI H S, HAN L X. Feeding ecology of Lophophorus sclat?eri at Gaoligong Mountain[J]. Journal of Southwest Forestry Uni?versity, 2010, 30(6): 64-67.
[2] CARACO T. Foraging theory[J]. Bulletin of Mathematical Biol?ogy, 1987, 49(5): 632-634.
[3] 尚玉昌. 行為生態學[M]. 2 版. 北京: 北京大學出版社,2018: 162-166.
SHANG Y C. Behavioural ecology[M]. 2nd ed. Beijing: PekingUniversity Press, 2018: 162-166.
[4] JENNINGS D E, GOULD J R, VANDENBERG J D, et al. Quan?tifying the impact of woodpecker predation on population dynamicsof the emerald ash borer (Agrilus planipennis)[J]. PLoS One,2013, 8(12): e83491.
[5] TARBILL G L, MANLEY P N, WHITE A M. Drill, baby, drill:the influence of woodpeckers on post-fire vertebrate communitiesthrough cavity excavation[J]. Journal of Zoology, 2015, 296(2):95-103.
[6] TREMBLAY J A, SAVARD J P L, IBARZABAL J. Structural re?tention requirements for a key ecosystem engineer in coniferdominatedstands of a boreal managed landscape in eastern Canada[J]. Forest Ecology and Management, 2015, 357: 220-227.
[7] MARTIN K, EADIE J M. Nest webs: a community-wide ap?proach to the management and conservation of cavity-nesting forestbirds[J]. Forest Ecology and Management, 1999, 115(2/3):243-257.
[8] PECHACEK P, D’OLEIRE-OLTMANNS W. Habitat use of thethree-toed woodpecker in central Europe during the breeding pe?riod[J]. Biological Conservation, 2004, 116(3): 333-341.
[9] ROBLES H, CIUDAD C, MATTHYSEN E. Responses to experi?mental reduction and increase of cavities by a secondary cavitynestingbird community in cavity-rich Pyrenean oak forests[J].Forest Ecology and Management, 2012, 277: 46-53.
[10] MIKUSI?SKI G, GROMADZKI M, CHYLARECKI P. Wood?peckers as indicators of forest bird diversity[J]. Conservation Bi?ology, 2001, 15(1): 208-217.
[11] AITKEN K E H, MARTIN K. The importance of excavators inhole-nesting communities: availability and use of natural treeholes in old mixed forests of western Canada[J]. Journal of Orni?thology, 2007, 148(Suppl. 2): S425-S434.
[12] FERNANDEZ C, AZKONA P. Influence of forest structure onthe density and distribution of the white-backed woodpecker Den?drocopos leucotos and black woodpecker Dryocopus martius inQuinto Real (Spanish western Pyrenees)[J]. Bird Study, 1996,43(3): 305-313.
[13] 孫美歐, 關大鵬, 孫虎. 涼水自然保護區科研現狀分析及未來發展建議[J]. 林業科技情報, 2021, 53(3): 31-34.
SUN M O, GUAN D P, SUN H. Analysis of scientific researchstatus and suggestions for future development of Liangshui Na?ture Reserve[J]. Forestry Science and Technology Information,2021, 53(3): 31-34.
[14] 侯宏宇. 涼水自然保護區地表鞘翅目成蟲集合群落格局及其構建機制[D]. 哈爾濱: 哈爾濱師范大學, 2020.
HOU H Y. Metaommunity pattern and maintenance mechanismof coleoptera adults in Liangshui Nature Reserve[D]. Harbin:Harbin Normal University, 2020.
[15] 王剛, 王波. 涼水自然保護區森林資源和生態現狀分析[J]林業科技情報, 2009, 41(2): 2-3.
WANG G, WANG B. Analysis on forest resources and ecologi?cal environment in Liangshui Nature Reserve[J]. Forestry Sci?ence and Technology Information, 2009, 41(2): 2-3.
[16] 許玲霞. 黑啄木取食生境偏好與潛在適宜生境[D]. 哈爾濱:東北林業大學, 2016.
XU L X. The feeding habitat preference and potential suitablehabitat of black woodpeckers[D]. Harbin: Northeast ForestryUniversity, 2016.
[17] BOYCE M S, MCDONALD L L. Relating populations to habitatsusing resource selection functions[J]. Trends in Ecology amp; Evo?lution, 1999, 14(7): 268-272.
[18] LENNON J J. Resource selection functions: taking space seri?ously?[J]. Trends in Ecology amp; Evolution, 1999, 14(10):399-400.
[19] MANLY B F J, MCDONALD L L, THOMAS D L, et al. Re?source selection by animals: statistical design and analysis forfield studies[J]. Journal of Animal Ecology, 1994, 63(3):745-746.
[20] MURTAUGH P A. The statistical evaluation of ecological indica?tors[J]. Ecological Applications, 1996, 6(1): 132-139.
[21] SWETS J A. Measuring the accuracy of diagnostic systems[J].Science, 1988, 240(4857): 1285-1293.
[22] PIROVANO A R, ZECCA G. Black woodpecker Dryocopus mar?tius habitat selection in the Italian Alps: implications for conser?vation in Natura 2000 network[J]. Bird Conservation Interna?tional, 2014, 24(3): 299-315.
[23] NAPPI A, DRAPEAU P, LEDUC A. How important is deadwood for woodpeckers foraging in eastern North American borealforests?[J]. Forest Ecology and Management, 2015, 346:10-21.
[24] SHORT L L. Burdens of the picid hole-excavating habit[J]. TheWilson Bulletin, 1979, 91(1): 16-28.
[25] 戎可, 司雨蕙, 潘麒嫣, 等. 同域分布3種啄木鳥冬季取食的生態位差異[J]. 生態學報, 2018, 38(23): 8314-8323.
RONG K, SI Y H, PAN Q Y, et al. Forage niche differentiationof three sympatric woodpecker species in winter[J]. Acta Eco?logica Sinica, 2018, 38(23): 8314-8323.
[26] GUNN J S, HAGAN J M Ⅲ. Woodpecker abundance and treeuse in uneven-aged managed, and unmanaged, forest in northernMaine[J]. Forest Ecology and Management, 2000, 126(1):1-12.
[27] BOCCA M, CARISIO L, ROLANDO A. Habitat use, homeranges and census techniques in the black woodpecker Dryocopusmartius in the Alps[J]. Ardea, 2007, 95(1): 17-29.
[28] KARIMI S, MORADI H V, REZAEI H Z, et al. Fine-scalehabitat use by black woodpecker Dryocopus martius: a yearroundstudy in the Hyrcanian forest, Iran[J]. North-westernJournal of Zoology, 2018, 14(1): 76-84.
[29] KHANAPOSHTANI M G, NAJAFABADI M S, KABOLI M, etal. Habitat requirements of the black woodpecker, Dryocopusmartius, in Hyrcanian forests, Iran[J]. Zoology in the MiddleEast, 2012, 55: 19-25.
[30] STACHURA-SKIERCZY?SKA K, KOSI?SKI Z. Do factors de?scribing forest naturalness predict the occurrence and abundanceof middle spotted woodpecker in different forest landscapes?[J].Ecological Indicators, 2016, 60: 832-844.
[31] ZAHNER V, SIKORA L, PASINELLI G. Heart rot as a key fac?tor for cavity tree selection in the black woodpecker[J]. ForestEcology and Management, 2012, 271: 98-103.
[32] LORENZ T J, VIERLING K T, JOHNSON T R, et al. The roleof wood hardness in limiting nest site selection in avian cavity ex?cavators[J]. Ecological Applications, 2015, 25(4): 1016-1033.
基金項目:國家自然科學基金面上項目(31970385)
