湖南省兩棲動物新紀錄
2024-06-29 00:00:00程彥林劉京周鋁周軍健李仕澤
野生動物學報 2024年2期


摘 要 2023 年5 月26 日,在湖南省新寧縣崀山鎮(26°19'40″ N,110°42'54″ E;海拔441 m)進行兩棲動物調查時,采集到4號琴蛙屬(Nidirana)物種標本,經形態特征比較,與桂北琴蛙(Nidirana guibeiensis)相似;基于線粒體COI基因構建的琴蛙屬部分物種系統發育樹顯示,此次采集的琴蛙標本與桂北琴蛙聚為一支,且具有較高的支持率(1. 00/99);基于Kimura雙參數模型估算本次采集的琴蛙標本與模式產地(廣西興安)標本間的遺傳距離為0,遠小于琴蛙屬物種間的遺傳距離(2. 7%~14. 4%)。綜合形態學和系統發育比較,確定此次采集的琴蛙標本為無尾目(Anura)蛙科(Ranidae)琴蛙屬的桂北琴蛙,系湖南省兩棲動物分布新紀錄種。
關鍵詞:琴蛙屬;桂北琴蛙;新紀錄;湖南省
中圖分類號:Q958. 1 文獻標識碼:A 文章編號:2310 - 1490(2024)- 02 - 0448 - 08
DOI:10.12375/ysdwxb.20240227
琴蛙屬(Nidirana)廣泛分布于東亞和東南亞的亞熱帶地區,目前該屬物種已報道20種[1],中國記錄分布有19種,其中,湖南省記錄分布有4種,分別為彈琴蛙(N. adenopleura)、粵琴蛙(N. guangdongen?sis)、雷山琴蛙(N. leishanensis)和湘琴蛙(N. xian?gica)[2?6]。彈琴蛙曾被記錄在湖南廣泛分布[5],近年來基于整合分類學研究,琴蛙屬內有多個隱存種被發現,且這些隱存種之前大多被誤定為彈琴蛙,因此,彈琴蛙在湖南是否有分布還有待進一步研究[6];粵琴蛙分布在湖南東南部八面山等地[2];雷山琴蛙分布在湖南西部懷化、通道等地[3];湘琴蛙分布在湖南湘江流域、湖南瀏陽大圍山和雙牌陽明山等地[2]。
2023年5月26日,在新寧縣崀山鎮進行兩棲動物調查時,于坪地村(26°19'40″ N,110°42'54″ E;海拔441 m)采集到4號(4 )琴蛙屬成體標本。經形態學比較和系統發育分析,鑒定為桂北琴蛙(N. guibei?ensis),為湖南省兩棲動物新紀錄種。本文對桂北琴蛙的形態特征及生態習性進行描述,并初步探討其形態特征的地理變異,為琴蛙屬物種的系統學和譜系地理學研究提供新信息。
1 材料與方法
1. 1 材料
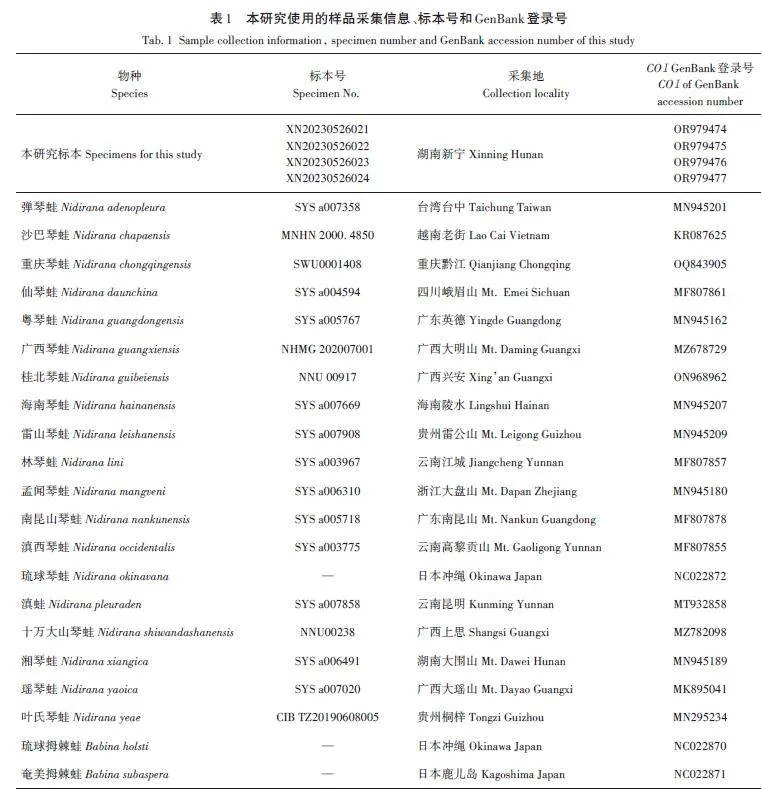
對采集的標本取少量肌肉組織存于95%的乙醇中用于提取DNA。標本浸泡于10%福爾馬林溶液中,保存于茅臺學院生物學標本室。本研究采集的4號標本及用于系統發育分析的部分琴蛙屬物種信息見表1。
1. 2 形態鑒定與測量
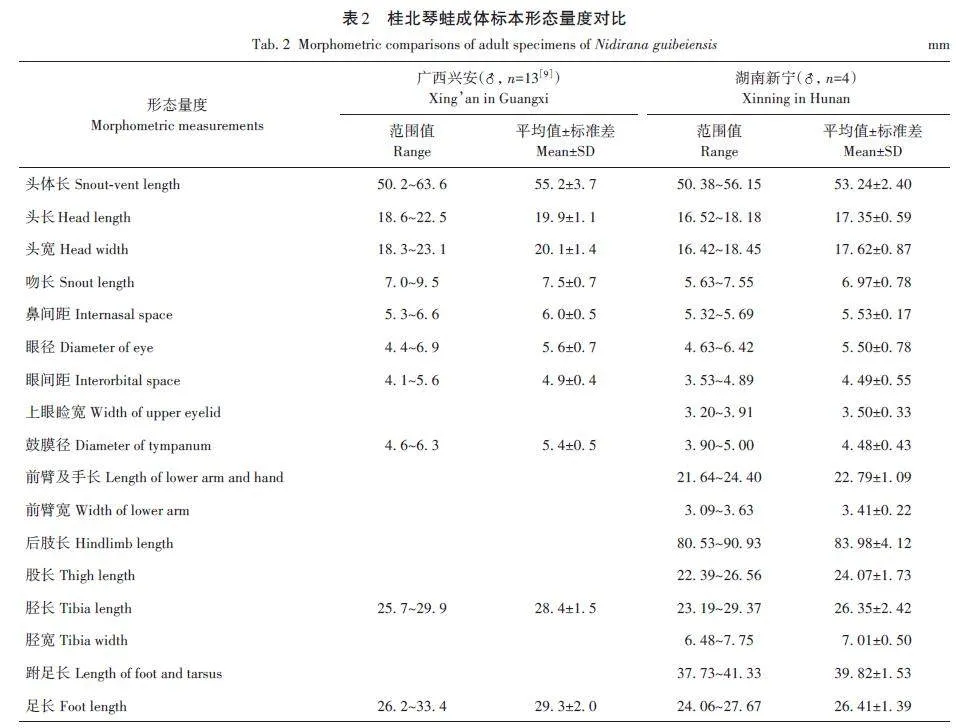
使用游標卡尺(無錫錫工量具有限公司,量程200 mm,精度0. 01 mm)測量4號標本的頭體長、頭長和頭寬等17項形態指標,并參照《中國兩棲動物檢索及圖解》[7]、《中國兩棲動物及其分布彩色圖鑒》[8]和桂北琴蛙形態描述[9]進行形態鑒定。
1. 3 分子鑒定
以高鹽法提取保存于乙醇中的肌肉組織DNA。參照Che等[10]引物序列擴增COI基因部分片段并測序,選用引物Chmf4:5'-TYTCWACWAAYCAYAAAGAYATCGG-3'和Chmr4:5'-ACYTCRGGRT-GRCCRAARAATCA-3'。PCR反應擴增體系為2. 50 μL 10×Buffer,2. 00 μL dNTP,0. 25 μL Taq 酶,上、下游引物各1. 00 μL,DNA 模板1. 00 μL,最后加入17. 25 μL滅菌超純水補足至終體積25. 00 μL。PCR擴增反應程序:95 ℃預變性4 min;94 ℃變性40 s,48 ℃退火40 s,72 ℃延伸60 s,共35 個循環;最后72 ℃延伸10 min。擴增產物用1%瓊脂糖凝膠電泳檢測大小和純度,大小符合要求的產物交生工生物工程(上海)股份有限公司測序。所得序列上傳到GenBank,登錄號為OR979474~OR979477。
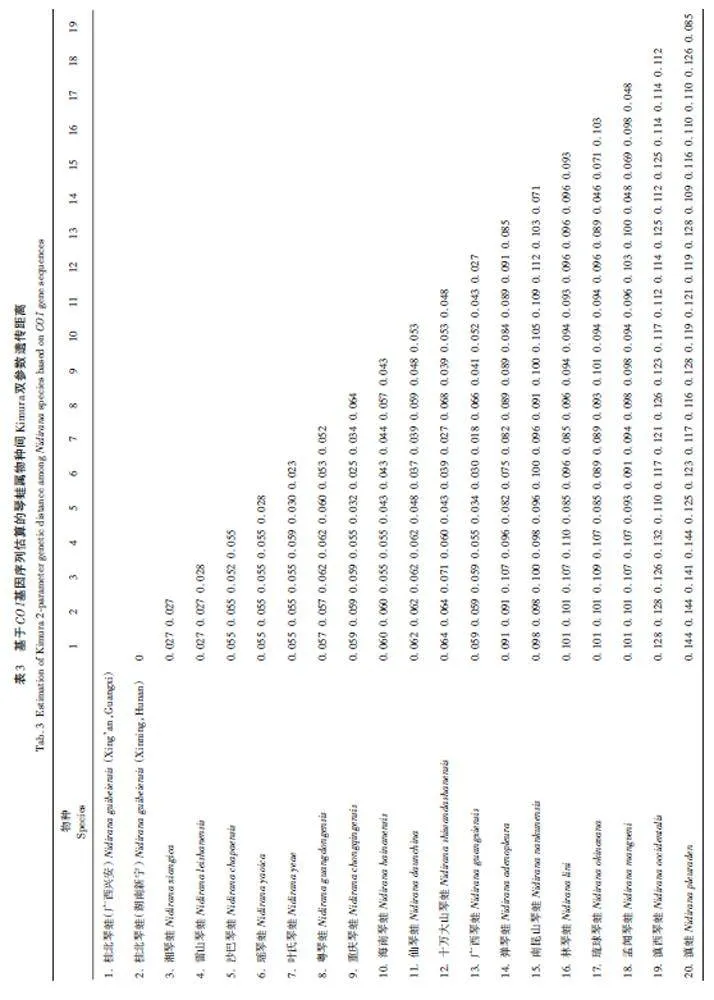
在GenBank中下載琴蛙屬部分物種和近緣類群拇棘蛙屬(Babina)物種的COI基因,結合本次采集標本所測序列進行系統發育分析。以琉球拇棘蛙(Babina holsti)和奄美拇棘蛙(Babina subaspera)為外群[9]。所有序列在軟件Clustal X v1. 8中比對[11],采用貝葉斯法(Bayesian inference of phylogeny,BI)和最大似然法(maximum likelihood,ML)重建系統發育關系。建樹前,利用軟件jModelTest v2 基于AIC 標準選擇最佳核苷酸替代模型[12]。最大似然樹(ML)在軟件PhyML v 3. 0 中完成[13],并進行1 000 次boot?strap 重抽樣分析,檢驗各節點置信度。采用MrBayes 3. 2. 4軟件進行貝葉斯分析[14],以隨機樹為起始樹,在默認熱值條件下同時運行4條馬爾可夫鏈,運行1×107代。每隔1 000代抽樣一次,舍棄25%的老化樹。使用Figtree軟件查看結果并獲得系統樹的支系結構和各支系的后檢驗概率。最后用MEGAv6. 0中的Kimura雙參數模型[15]計算表1中所列琴蛙屬物種間的遺傳距離。
2 結果
2. 1 形態描述
本次采集的4號標本均為雄性個體(XN20230526021~XN20230526024),頭體長50. 38~56. 15 mm,其他部位量度見表2。標本吻端鈍圓,頭長和頭寬幾乎相等,瞳孔橫置,鼓膜明顯,大而圓,邊緣略突出,舌后端有明顯缺刻,犁骨齒發達;前臂較粗,手指纖細,指端略微膨大,指長序Ⅲgt;Ⅰgt;Ⅳgt;Ⅱ,掌突2個,呈橢圓形,指關節下瘤明顯;左右跟部重疊明顯,脛跗關節貼體前伸達眼鼻之間,趾長序Ⅳgt;Ⅲgt;Vgt;Ⅱgt;Ⅰ,趾端有腹側溝,趾間具2/3蹼,蹼式為I2-2Ⅱ1?-3-Ⅲ2-3+Ⅳ3+-2-V,趾關節下瘤明顯,第Ⅳ趾上有3個,第Ⅲ、V趾上有2個,第Ⅰ、Ⅱ趾上有1個;內蹠突呈長條狀,外蹠突橢圓;背部淺棕色,體側淡黃色;背部及體側有黑色斑;背中線明顯,呈棕色,四肢棕黃色;鼓膜棕褐色;頜腺和肩腺均為乳白色;腹部光滑,四肢及咽部肉色,腹部白色;具1對內聲囊,第Ⅰ指具婚墊(圖1)。
2. 2 系統發育關系和遺傳距離
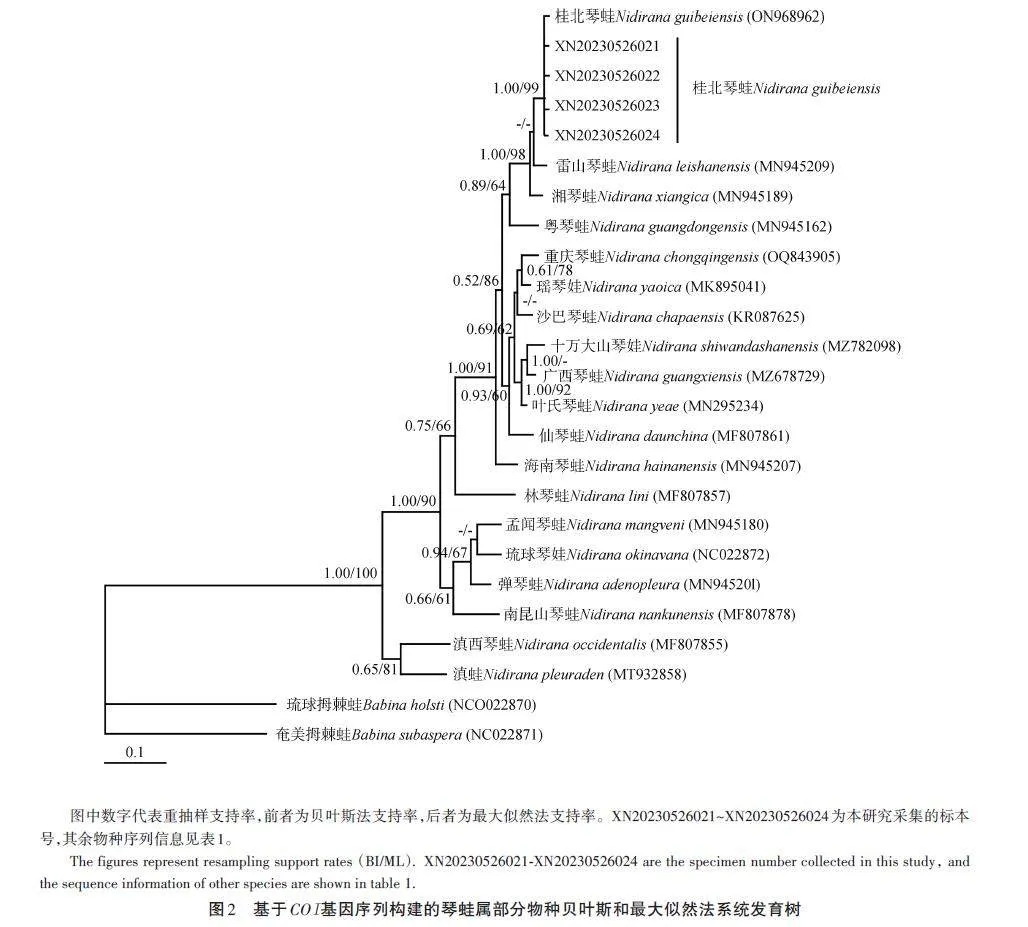
PCR擴增獲得的琴蛙線粒體COI基因片段長度為672 bp。在jModelTest v2軟件中基于AIC標準計算了最佳核苷酸替代模型,本文用于系統發育分析的最佳核苷酸替代模型為GTR+G+I。
基于貝葉斯法和最大似然法構建系統發育樹(圖2)顯示,采自湖南新寧的琴蛙標本與桂北琴蛙模式產地(廣西興安)的標本聚為一支,且有較高的支持率(1. 00/99)。基于Kimura雙參數模型估算本研究所采琴蛙屬部分物種的遺傳距離為2. 7%~14. 4%(表3),平均遺傳距離為7. 5%。本研究所采集的琴蛙標本與桂北琴蛙模式產地(廣西興安)標本間的遺傳距離為0,說明兩個種群間共享同一單倍型,未出現遺傳分化。通過形態比較,在新寧采集的琴蛙標本與桂北琴蛙原始文獻[8]的描述吻合,系統發育分析也支持采自湖南新寧的琴蛙標本與桂北琴蛙模式產地標本聚為一支。
因此,綜合形態比較與分子系統學分析,鑒定采集于湖南新寧崀山鎮的琴蛙標本為桂北琴蛙,為湖南省兩棲動物分布新紀錄。
2. 3 生境描述
本次桂北琴蛙標本的采集地位于湖南省新寧縣崀山鎮坪地村海拔441 m的農田附近,發現時雄蛙正在鳴叫(圖3)。與其同域分布的兩棲類還有中華湍蛙(Amolops sinensis)、闊褶水蛙(Hylaranalatouchii)、澤陸蛙(Fejervarya multistriata)、三港雨蛙(Hyla sanchiangensis)、斑腿泛樹蛙(Polypedatesmegacephalus)和飾紋姬蛙(Microhyla fissipes)等。
3 討論
琴蛙屬物種在中國南方廣泛分布,大多數物種間形態較為相似,因此據形態特征很難對其進行鑒定[8]。近年來,隨著分子生物學的發展,采用形態比較結合分子系統學分析對物種進行鑒定的方法已被廣泛應用。
本研究通過下載已有報道的琴蛙屬部分物種的COI序列,構建系統發育樹,計算遺傳距離,發現本研究所采標本和湖南省記錄分布的彈琴蛙、湘琴蛙、粵琴蛙和雷山琴蛙的遺傳距離分別為9. 1%、2. 7%、5. 7%和2. 7%,和模式產地(廣西興安)的桂北琴蛙遺傳距離為0,遠小于其他琴蛙屬的遺傳距離。
采自湖南新寧的琴蛙標本與湖南已記載的4種琴蛙屬物種在形態上存在一定差異[2?3,5,9]。桂北琴蛙頭長與頭寬幾乎相等,區別于湘琴蛙、粵琴蛙和雷山琴蛙的頭長大于頭寬;無雄性線,區別于彈琴蛙的有雄性線;掌突2個,區別于彈琴蛙、湘琴蛙、粵琴蛙和雷山琴蛙的3 個掌突;有弱的顳褶,區別于彈琴蛙、湘琴蛙、粵琴蛙和雷山琴蛙的無顳褶。本次采集的桂北琴蛙和模式產地(廣西興安)標本在形態上也存在一些差異,新寧桂北琴蛙后肢貼體前伸達眼中部,不同于模式產地桂北琴蛙的達鼻眼之間。但由于本次所采桂北琴蛙樣本數量有限,上述差異是否為適應不同環境而導致的形態變異,還有待進一步論證。
桂北琴蛙為2022 年新發表的兩棲動物新物種,此前在我國僅分布于廣西興安縣和貓兒山國家級自然保護區,棲息在海拔300~1 300 m的稻田、靜水塘旁[9]。此次在湖南省新寧縣崀山鎮坪上村(海拔441 m)發現桂北琴蛙,不僅豐富了其分布信息,也將桂北琴蛙的分布區域向北擴展至湖南中南部。結合已有的報道,本研究推測桂北琴蛙可能沿資江流域分布,因此,建議在該區域進行全面詳細地調查,以了解桂北琴蛙確切的分布范圍及種群狀況。
參考文獻:
[1] FROST D R. Amphibian species of the world: an online refer?ence: version 6. 0 [DB/OL]. [2023-09-20]. https://amphibian?softheworld. amnh. org/.
[2] LYU Z T, DAI K Y, LI Y, et al. Comprehensive approaches re?veal three cryptic species of genus Nidirana (Anura, Ranidae)from China[J]. ZooKeys, 2020, 914: 127-159.
[3] 劉京, 程彥林, 周鋁, 等. 湖南通道發現雷山琴蛙[J]. 動物學雜志, 2022, 57(1): 105-111.
LIU J, CHENG Y L, ZHOU L, et al. Nidirana leishanensis foundin Tongdao, Hunan[J]. Chinese Journal of Zoology, 2022, 57(1): 105-111.
[4] 中國科學院昆明動物研究所.“ 中國兩棲類”信息系統[DB/OL].[ 2023-09-20]. http://www. amphibiachina. org/.Kunming Institute of Zoology. The database of Chinese amphibian[DB/OL].[ 2023-09-20]. http://www. amphibiachina. org/.
[5] 沈猷慧, 楊道德, 莫小陽, 等. 湖南動物志: 兩棲綱[M]. 長沙: 湖南科學技術出版社, 2014: 236-240.
SHEN Y H, YANG D D, MO X Y, et al. Fauna Hunan: Am?phibia[M]. Changsha: Hunan Science and Technology Press,2014: 236-240.
[6] 高志偉, 錢天宇, 江建平, 等. 湖南省兩棲、爬行動物物種多樣性及其地理分布[J]. 生物多樣性, 2022, 30(2): 97-111.
GAO Z W, QIAN T Y, JIANG J P, et al. Species diversity anddistribution of amphibians and reptiles in Hunan Province, China[J]. Biodiversity Science, 2022, 30(2): 97-111.
[7] 費梁, 葉昌媛, 黃永昭, 等. 中國兩棲動物檢索及圖解[M]. 成都: 四川科學技術出版社, 2005: 6-13; 119-121.
FEI L, YE C Y, HUANG Y Z, et al. An illustrated key to Chi?nese amphibians[M]. Chengdu: Sichuan Publishing House of Sci?ence and Technology, 2005: 6-13; 119-121.
[8] 費梁, 葉昌媛, 江建平. 中國兩棲動物及其分布彩色圖鑒[M]. 成都: 四川科學技術出版社, 2012:344-345.
and their distributions[M]. Chengdu: Sichuan Publishing Houseof Science amp; Technology, 2012:344-345.
[9] CHEN W C, YE J P, PENG W X, et al. A new species of Nidi?rana( Anura, Ranidae) from northern Guangxi, China[J]. ZooK?eys, 2022, 1135: 119-137.
[10] CHE J, CHEN H M, YANG J X, et al. Universal COI primersfor DNA barcoding amphibians[J]. Molecular Ecology Re?sources, 2012, 12(2): 247-258.
[11] THOMPSON J D, GIBSON T J, PLEWNIAK F, et al. TheCLUSTAL_X windows interface: flexible strategies for multiplesequence alignment aided by quality analysis tools[J]. NucleicAcids Research, 1997, 25(24): 4876-4882.
[12] POSADA D. jModelTest: phylogenetic model averaging[J]. Mo?lecular Biology and Evolution, 2008, 25(7): 1253-1256.
[13] GUINDON S, DUFAYARD J F, LEFORT V, et al. New algo?rithms and methods to estimate maximum-likelihood phylog?enies: assessing the performance of PhyML 3. 0[J]. SystematicBiology, 2010, 59(3): 307-321.
[14] RONQUIST F, HUELSENBECK J P. MrBayes 3: Bayesian phy?logenetic inference under mixed models[J]. Bioinformatics,2003, 19(12): 1572-1574.
[15] TAMURA K, STECHER G, PETERSON D, et al. MEGA6: mo?lecular evolutionary genetics analysis version 6. 0[J]. MolecularBiology and Evolution, 2013, 30(12): 2725-2729.
基金項目:國家自然科學基金項目(31960099,32260136);貴州省科技計劃項目(黔科合基礎-ZK[2022]一般540);貴州省林業科研課題項目(黔林科合[2020]13號);茅臺學院高層次人才科研啟動經費項目(mygccrc[2022]055,mygccrc[2022]067)
