深型水庫溶解氧分層規律及其機制探究
2024-05-23 08:32:36陳侃王飛飛阮琪禎吳旭春曹文志
水生態學雜志 2024年2期
陳侃 王飛飛 阮琪禎 吳旭春 曹文志
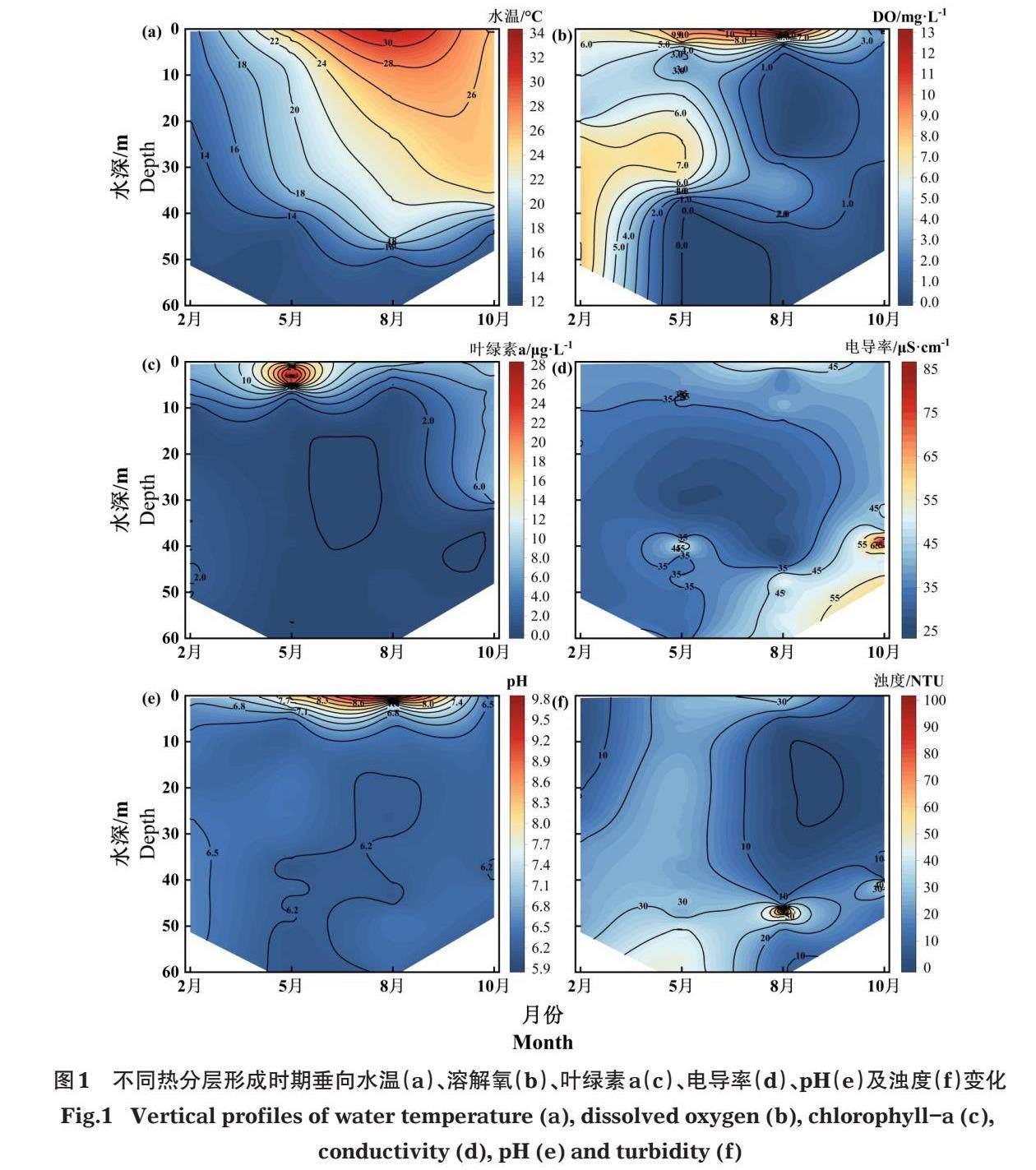
摘要:溶解氧(DO)是衡量生態狀況的重要指標,探討深型水庫DO分層規律與營養元素生物地球化學循環間的關系,可為維護庫區水生態健康提供科學依據。2022年監測了九龍江萬安水庫的理化指標及營養元素的垂向分布,并分析了庫區的DO分層規律及影響因素。結果表明:2月屬萬安水庫混合期,呈貧營養狀態,DO垂向平均濃度高達6.70 mg/L,DO變化受浮游植物分布及“翻庫”影響;5月屬熱分層形成期,處中營養狀態,水體在內源產氧減少和原位DO消耗驅動作用下出現溫躍層溶解氧極小值,36 m以下水體處于缺氧狀態;8月為熱分層穩定期,處中度富營養化,表層DO最大濃度達12.00 mg/L以上,在35 m深度出現DO極大值,水體熱分層阻礙DO補充、內源產氧減弱是造成熱分層穩定期水體上層大范圍缺氧的主要原因;10月為熱分層消退期,呈中營養狀態,DO垂向平均濃度僅為0.77 mg/L,水深2.79 m以下水層皆處于缺氧狀態。熱分層時期DO垂向變化受到水體熱分層、營養水平、污染狀態及浮游植物生長分布等因素的影響,熱分層從物理角度影響DO分層,營養水平及污染狀態可以影響DO的產耗機制,而庫區水體濁度通過影響浮游植物的垂向分布進而對DO濃度產生影響。
關鍵詞:溶解氧;水體熱分層;缺氧狀態;萬安水庫
中圖分類號:X142? ? ? ? 文獻標志碼:A? ? ? ? 文章編號:1674-3075(2024)02-0020-11
溶解氧(Dissolved oxygen, DO)是指O2以分子態溶解于水中的形態(李艷紅等,2013)。水體DO的主要來源是大氣根據氧分壓的大小向DO未飽和的水體中充氧,以及水生生物進行光合作用釋放到水中的氧(呂琳莉等,2018)。水體中的DO濃度能夠反應水體污染程度,是水質的重要參數,也是湖庫水質管理的重要指標之一。水中有機物的生物降解過程消耗了大量的O2,DO耗盡促使有機物厭氧分解,產生一些有毒、有難聞氣味的氣體有機物,使水環境進一步惡化。因此,DO濃度影響到湖庫用水質量,并直接關系居民用水安全(陳東等,2008;Harke et al,2016;Torres et al,2016)。DO不僅影響營養物質循環及溫室氣體排放,充足的DO還是水生生物生存的必要條件,許多水生生物需要DO充足的棲息地和適宜的水溫才可生存(Fernandez et al,2014;Schindler,2017;McClure et al,2018)。當DO低于5.0 mg/L時,存在威脅魚類與浮游動物生存的可能,而水體處于低氧或厭氧狀態時(一般低于2.0 mg/L)會影響水生生物的生存繁衍。此外,湖庫底部的低DO濃度還會通過降低氧化還原電位(Eh),促進沉積物中有毒物質、還原性污染物(如氨氮)等釋放,從而威脅到魚類、底棲生物等生存,并危害水生態系統健康(Singleton and Little,2006;Matzinger et al,2010;North et al,2014)。
在全球氣候變暖的背景下,庫區的深層水溫穩定,但表層水溫上升顯著,庫區水體分層結構更加穩定、生物異養呼吸速率增加、沉積物耗氧及氧氣溶解度下降等因素驅動了DO的減少(Muller et al,2012;Jane et al,2021)。但大壩的修建、水資源過度開發利用等人類干擾行為改變了流域生態狀況,工農業、生活排污等向湖庫輸入營養物質與污染物,營養水平高的庫區藻類暴發頻繁,其強烈的光合作用常使得表層水體DO超飽和(Kosten et al,2012;Schindler,2017)。DO存在很高的生化反應參與度,這使得DO濃度垂向變化規律特別是缺氧現象很難預測(Yvon-Durocher et al,2010;Woolway & Merchant,2019)。庫區水體常被視作“半封閉”水體,其水力停留時間長,故季節變化所引起的水體熱分層通常較穩定,在此驅動下DO也呈現出較穩定的垂向分層規律(Wagner & Adrian,2011;Jones et al,2011)。現有研究將水深30 m以上的水庫界定為深型水庫,易形成由混合層、變溫層及滯溫層組成的熱分層結構(李秋華等,2013;鄭瑩瑩等,2021)。變溫層中溫度變化梯度最大的薄水層稱為溫躍層,湖庫水體熱分層的穩定性與其水溫、水體透明度等物理化學因素及湖泊形態學參數密切相關(Demers & Kalff,1993;Houser,2006;Huang,2016)。熱分層出現的根本原因是季節變化導致水體的上下層間出現溫度差異從而產生水層密度差,熱分層不僅阻礙水層交換,并對生物的活性和生化反應速率產生影響,進而影響DO分布規律(Saito et al,2001;Caliskan & Elci,2009;Borics et al,2015;Zhang et al,2015)。深型庫區滯溫層在一年中長時間存在大范圍缺氧水層,DO補充受阻、浮游植物及有機質下沉分解與沉積物耗氧等是缺氧現象出現的主要原因。此外,流域內人類活動影響,水質進一步惡化,對缺氧層的形成存在一定的促進作用(Diaz & Rosenberg,1995;Muller et al,2012)。
國內外研究雖已對湖庫DO垂向分布規律及機制進行了廣泛探討,但卻缺乏對深型湖庫中水體營養狀態及營養元素生物地球化學循環與DO分層規律相互影響關系的研究。本研究以我國東南地區萬安水庫為研究對象,分析DO在不同時期的垂向變化特征,探究水層缺氧及變溫層DO極小值等現象出現的內在原因,DO垂向分層規律與水層營養狀況間的響應關系,以及DO變化規律與營養元素生物地球化學循環間的關系,以期為維護庫區的水生態健康提供科學依據。
1? ?材料與方法
1.1? ?研究區域概況
萬安水庫(25°20'58"~25°24'36" N,116°58'30"~117°03'36" E)位于福建省南部的九龍江北溪上游,壩址位于龍巖市萬安鎮境內,1994年底開始蓄水。壩址以上流域面積667 km2,多年平均流量23.3 m3/s,水庫正常蓄水位365 m,總庫容約2.29×108 m3,調節庫容1.68×104 m3,為多年調節水庫。最大壩高93.8 m,屬于混合式電站,設計水頭84.7 m,多年平均發電量約為1.36×108 kW·h。水流入庫匯流處存在一定規模的漁業養殖活動(林炳明,2006)。
1.2? ?樣品采集與測定
選取萬安水庫深水區(年平均水深大于55 m),分別于2022年2月、5月、8月及10月進行4個季節的采樣。現場使用Hondex便攜式數字聲納深度計測量水深,并采用YSI-EXO3便攜式多參數水質測試儀垂向連續測定DO、水溫、pH、葉綠素a、濁度及電導率,以葉綠素a濃度表征浮游植物生物量。并使用5 L有機玻璃采水器分別在水深0.5、10、25、35、45及60 m采集水樣并將其低溫保存于聚乙烯水樣瓶中帶回實驗室測定總氮(TN)、總磷(TP)、營養鹽五參數(NO2–、NO3–、NH4+、PO43-、SiO32-)及高錳酸鹽指數(CODMn)等相關指標。
TN和TP分別采用堿性過硫酸鉀消解紫外分光光度法(HJ636-2012)和鉬酸銨分光光度法(GB 11893-89)測定,CODMn采用高錳酸鹽指數測定標準(GB 11892-89)測定,營養鹽五參數采用連續流動化學分析儀(AA3,德國Bran+Luebbe公司)進行測定。
1.3? ?數據處理
根據《湖泊(水庫)富營養化狀況評價方法及分級技術規定》,富營養化狀態評價采用綜合營養狀態指數法(金相燦,1995)。評價中以0~3 m水深的葉綠素a平均濃度代表表層水體葉綠素a水平,以水深0.5 m的TN、TP及CODMn濃度代表三者在表層水體中的水平。
本文將垂直方向上溫度梯度大于0.2°C/m的水層定義為溫躍層,其是變溫層中水溫垂直梯度較大的一個水層;DO濃度梯度大于0.2 mg/(L·m)的水層定義為氧躍層,溫躍層及氧躍層的深度分別定義為溫躍層和氧躍層上部邊界的深度,溫躍層底部和氧躍層底部分別定義為溫躍層和氧躍層下界的深度,溫躍層及氧躍層厚度則為上下邊界的差值(Wilhelm & Adrian,2008;Zhang et al,2015)。將水體DO濃度小于2.0 mg/L定義為缺氧狀態。
采用Excel、SPSS 27.0進行數據處理及統計分析,并使用Origin 2018繪制垂向數值圖。
2? ?結果與分析
2.1? ?DO及環境因子分層規律
2.1.1? ?DO及基礎理化指標? ?萬安庫區不同季節呈現出混合期(2月)、熱分層形成期(5月)、熱分層穩定期(8月)及熱分層消退期(10月)的變化特征(圖1-a)。混合期水體無顯著熱分層,垂向水溫變化范圍12.59~15.24°C。進入熱分層形成期,滯溫層以上水溫顯著上升,熱分層結構形成(第1溫躍層+混合層+第2溫躍層+滯溫層),雙溫躍層分別出現在0.21 m及33.08 m處,厚度為7.24 m和7.86 m,水溫梯度達0.67°C/m和0.63°C/m。熱分層穩定期的水體分層結構更加穩固(第1溫躍層+混合層+第2溫躍層+滯溫層),垂向平均水溫升高至21.78°C,雙溫躍層分別出現在水深0.65 m及44.57 m處,厚度為8.23 m和5.30 m,水溫梯度達0.77℃/m和1.26℃/m,滯溫層深度較熱分層形成期增大。熱分層消退期水體分層強度減弱(混合層+變溫層+滯溫層),溫躍層出現在水深34.64 m處,厚度為9.51 m,水溫梯度達到了1.08℃/m,滯溫層深度減小。
在水體熱分層驅動下,DO垂向分層在不同季節也呈現出明顯變化特征(圖1-b)。在混合期,庫區水體DO垂向變化(4.12~8.04 mg/L)較小,DO垂向濃度平均值(6.70 mg/L)較高,僅在8.66 m水深處出現氧躍層,其厚度為1.65 m,濃度梯度為0.27 mg/(L·m),未出現缺氧狀態水層。垂向DO分布規律在混合期呈“下高上低”,熱分層形成期、穩定期及消退期則呈“上高下低”。進入熱分層形成期,滯溫層DO濃度下降,水體在第1個溫躍層中出現了典型的DO極小值現象;2個氧躍層分別出現在1.37 m和31.33 m處,厚度分別為4.07 m和5.90 m,濃度梯度為1.53 mg/(L·m)和1.10 mg/(L·m),且低于36.68 m水深時處于缺氧狀態。在熱分層穩定期中,表層水體DO最大濃度達12.00 mg/L以上,2個氧躍層分別出現在1.01 m及38.74 m處,厚度分別為13.19 m和6.71 m,濃度梯度為0.90 mg/(L·m)和0.26 mg/(L·m);并在水深5.09 m處出現了厚度23.22 m的缺氧水層,在35 m深度出現DO極大值現象,水深低于40.39 m再次進入缺氧狀態,滯溫層中的缺氧層出現深度較熱分層形成期時有所增大。隨著庫區進入熱分層消退期,垂向DO濃度顯著下降,垂向平均濃度僅為0.77 mg/L,2個氧躍層分別出現在0.85 m及29.80 m處,厚度分別為2.04 m和3.10 m,濃度梯度為0.93 mg/(L·m)和0.22 mg/(L·m)。缺氧現象較熱分層穩定期進一步加劇,深度2.79 m以下的水層皆處于缺氧狀態。
萬安庫區各時期葉綠素a整體呈“上高下低”分布趨勢(圖1-c)。在混合期、熱分層形成期及熱分層穩定期,葉綠素a濃度在10 m以上水層顯著高于10 m以下水層,且在10 m水深處降至接近0。熱分層形成期溫躍層DO濃度最小的水深中葉綠素a濃度達27 μg/L以上。熱分層消退期中水深30 m以上葉綠素a濃度穩定,至第2個氧躍層及溫躍層范圍內,葉綠素a濃度顯著下降至接近0。
庫區水體電導率在混合期變化較小(30.1~37.7 [μ]S/cm),在其他時期變化范圍大(圖1-d)。在熱分層形成期及熱分層穩定期,水體電導率在第1個溫躍層及氧躍層位置顯著下降,在第2個溫躍層及氧躍層位置上升。熱分層消退期中的水體電導率始終高于40 [μ]S/cm,在第2個溫躍層位置由41 [μ]S/cm增至80 [μ]S/cm。
各時期pH整體呈“上高下低”狀態,大部分水體屬于弱酸性,水體混合期及熱分層消退期pH整體變化范圍小;熱分層形成及穩定期pH變化較大(6.07~8.04及6.15~9.47),在0~10 m水層的pH表現出與同深度DO變化相同的下降趨勢(圖1-e);且在熱分層形成期的第2個溫躍層及氧躍層出現位置,pH顯著下降。
水體濁度在熱分層形成期最高,各深度水體濁度均高于20 NTU,混合期及熱分層形成期的水體濁度在滯溫層呈顯著增大趨勢(圖1-f)。在熱分層穩定期,濁度在第1個溫躍層及氧躍層位置顯著下降,在第2個溫躍層位置上升。熱分層消退期中的水體濁度在第2個溫躍層范圍內顯著增加。
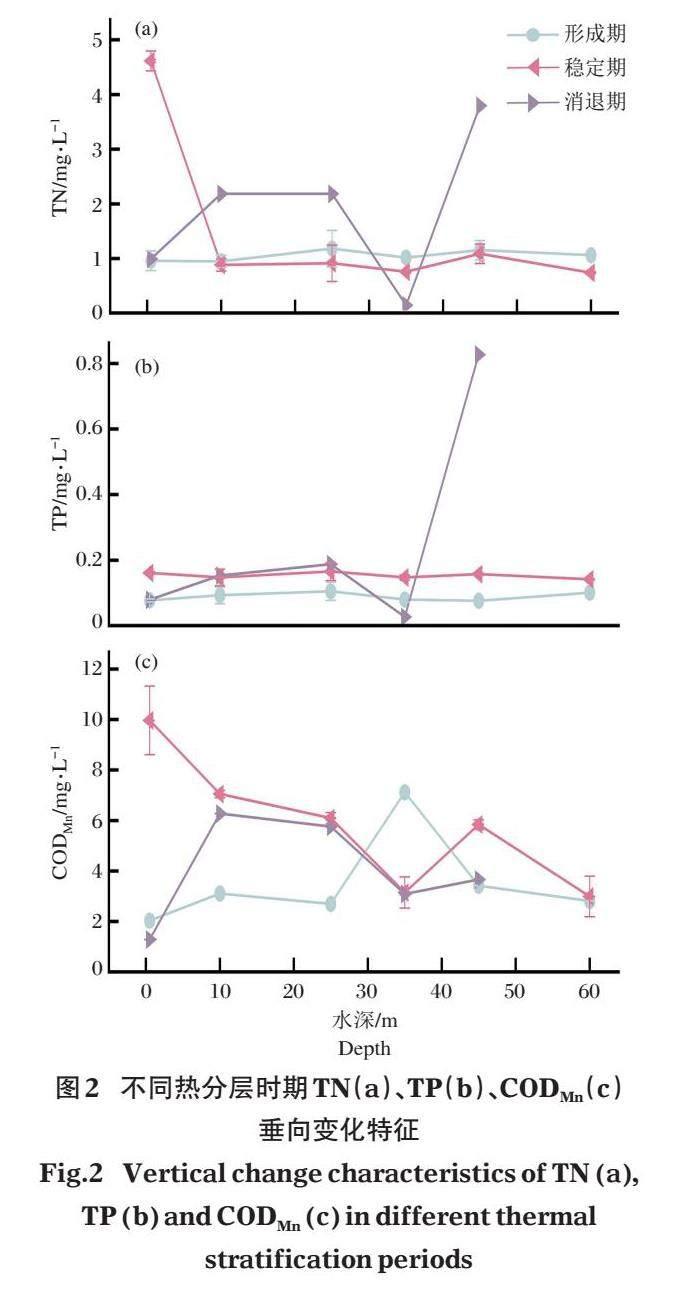
2.1.2? ?營養物質垂向變化特征與庫區營養狀態? ?庫區水體熱分層形成期的TN、TP及CODMn的變化范圍小于熱分層穩定期及消退期(圖2)。在熱分層形成期,庫區水體的TN在垂向上穩定。熱分層穩定期時,庫區表層的TN濃度超過Ⅴ類地表水環境質量標準的限值,其在第1個變溫層中以0.46 mg/(L·m)的梯度下降,在混合層、第2變溫層及滯溫層中保持穩定。在熱分層消退期,TN在10 m以上水層呈上升趨勢。在第2個溫躍層中出現極小值,滯溫層TN明顯回升,超過Ⅴ類地表水環境質量標準的限值(圖2-a)。
熱分層形成期及消退期的水體TP濃度在垂向上保持穩定。在熱分層消退期,庫區混合層的TP保持穩定,第2個溫躍層位置TP出現極小值,在滯溫層回升,超過Ⅴ類地表水環境質量標準的限值(圖2-b)。
熱分層形成期的水體CODMn在水表變溫層中呈上升趨勢,在第2個溫躍層中出現CODMn的極大值。在熱分層穩定期,水體CODMn在混合層中持續下降,在DO極大值位置出現極小值,在滯溫層中略有回升后下降。熱分層消退期的水體CODMn在混合層中呈先上升后下降趨勢(圖2-c)。
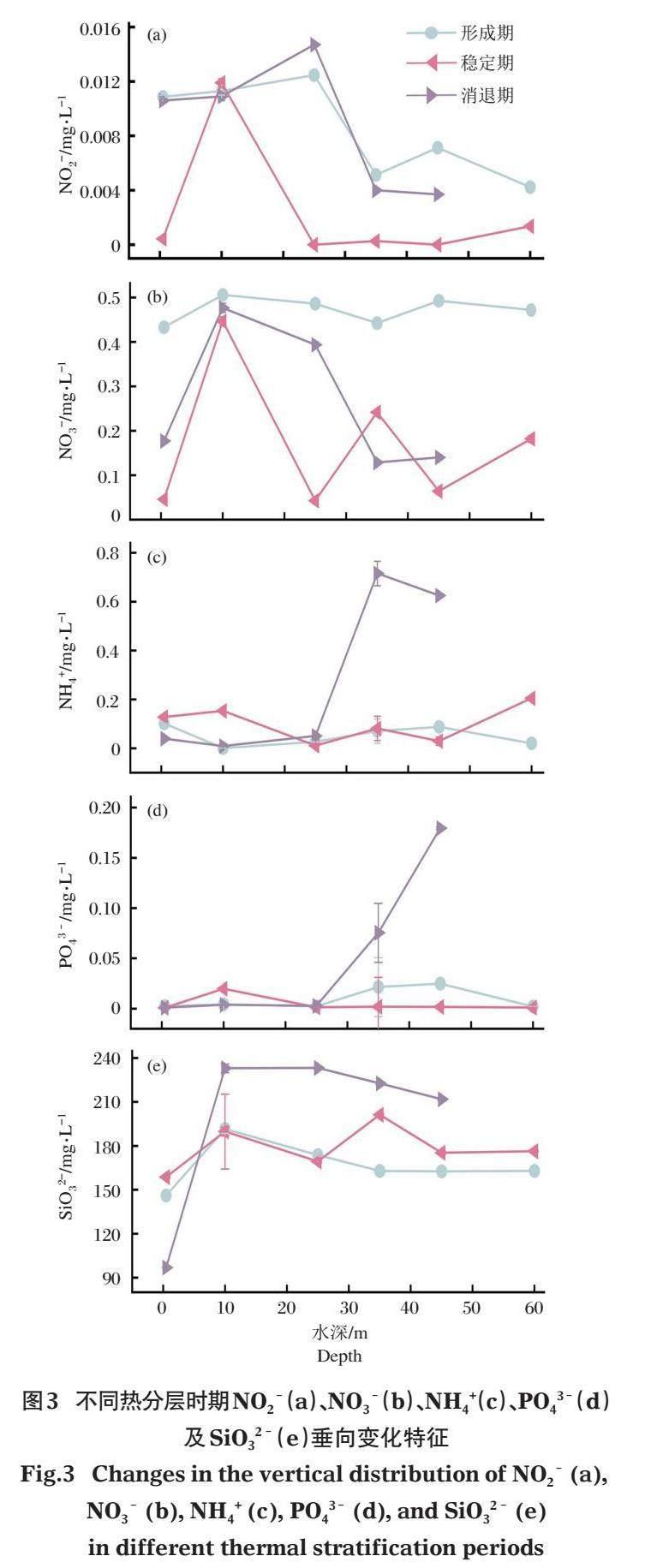
在熱分層形成期,庫區水體NO2–垂向變化較大,其余4項指標的變化較小(圖3)。在溫躍層DO極小值出現位置,NO2–、NO3–及SiO32–呈上升趨勢,NH4+呈下降趨勢。在第2個溫躍層及氧躍層出現位置,NO2–及NO3–出現極小值,NH4+及PO43–略有上升。隨著熱分層結構穩定,庫區水體的NO2–及NO3–變化范圍增大。第1個溫躍層及氧躍層范圍內的NO2–、NO3–、NH4+、PO43–及SiO32–均呈現上升趨勢。在DO極大值出現位置,NO3–及SiO32–也達到極大值,NO2–及NH4+略有增加后減少。在第2個氧躍層范圍內,NO3–、NH4+及SiO32–均呈下降趨勢。而NO3–及NH4+在滯溫層明顯增大,NO2–含量也略有上升。進入熱分層消退期,營養鹽五參數的變化范圍均增大,水體表層氧躍層中NO2–、NO3–及SiO32–呈上升趨勢,NH4+呈下降趨勢。在第2個溫躍層出現位置,NO2–、NH4+及SiO32–不同程度減少,NO3–及PO43–不同程度增加。
根據富營養化狀態評價結果,萬安庫區表層水體的營養狀態在混合期處貧營養級,在熱分層形成期及熱分層消退期皆處于中營養級,在熱分層穩定期更是達到中度富營養化(表1)。庫區水體的營養水平在熱分層形成后顯著提高,熱分層時期較高的營養狀態對DO的垂向分布規律存在很大影響(Jane et al,2021)。
2.2? ?不同熱分層時期DO分層規律變化的影響因素
將熱分層不同時期各分層水體的DO與環境因子進行相關性分析,結果見圖4。混合期中,DO與水溫、pH及濁度呈正相關,與電導率呈極顯著負相關;葉綠素a與水溫呈極顯著負相關,與pH呈極顯著正相關,但與濁度不存在顯著相關性;pH與濁度呈極顯著正相關。熱分層形成期及穩定期的表層變溫層及混合層中DO與環境因子的相關性結果類似:庫區表層變溫層的DO與水溫、葉綠素a、電導率、pH及濁度皆呈極顯著正相關,同時葉綠素a與濁度呈極顯著正相關;2個時期的混合層中DO與水溫、葉綠素a及電導率呈極顯著負相關,而與pH呈極顯著正相關;葉綠素a與濁度呈極顯著正相關。在熱分層形成期的第2個變溫層中,DO與水溫及pH呈極顯著正相關,而與電導率呈極顯著負相關;葉綠素a與pH及濁度呈極顯著負相關,而pH與濁度呈極顯著正相關。熱分層穩定期的第2個變溫層中DO與水溫及濁度呈極顯著正相關,與pH呈極顯著負相關,同時與葉綠素a呈顯著負相關。在熱分層消退期,混合層DO與水溫、葉綠素a及pH呈極顯著正相關,與電導率及濁度呈極顯著負相關;變溫層中DO與水溫及葉綠素a呈極顯著正相關,與電導率、pH及濁度呈極顯著負相關,滯溫層DO濃度已降至0 mg/L。
水體熱分層、浮游植物生物量及分布與濁度是影響DO變化的關鍵環境因子(圖4)。為探究水體熱分層對DO分層規律的影響,對萬安庫區氧躍層與溫躍層的出現深度、厚度以及變化梯度進行線性擬合,混合期未出現溫躍層故不納入分析(圖5)。氧躍層與溫躍層在深度上存在極顯著的正線性關系(P<0.001),在梯度上存在顯著負線性關系(P<0.05),但在厚度上卻不存在顯著的線性關系。
依據庫區水體濁度的垂向變化特征及浮游植物分布特征,將0~5 m和5~10 m水層葉綠素a濃度變化梯度與濁度進行線性擬合,發現兩者存在極顯著的正線性關系(P<0.001)(圖6)。其中,熱分層形成期的水體平均濁度與葉綠素a濃度變化梯度的擬合關系較差,兩者在混合期、熱分層穩定期及消退期的擬合關系較好。
3? ?討論
3.1? ?混合期DO分層變化機制
在混合期影響水體DO變化的主要因素是浮游植物生物量。庫區在混合期未出現熱分層,盡管DO與水溫呈極顯著正相關,但水溫不是影響DO變化的最主要因素。水體中浮游植物的光合作用消耗CO2,釋放O2,氫離子濃度隨之降低,從而導致pH上升。根據CO2水解方程式:
當水體中的呼吸作用強于光合作用時,生物消耗O2分解有機物質,產生CO2,氫離子濃度上升,pH和DO隨之降低(趙紫涵等,2020;郭詩君等,2021)。水體上層DO下降的主要原因是浮游植物生物量的減少,pH的下降也證明了這一點。但在混合期貧營養及低水溫的條件下,浮游植物生長受限制,故浮游植物生物量變化引起的DO下降梯度較小。庫區在混合期未出現缺氧現象,并且DO濃度及濁度分布呈“上低下高”,同時滯溫層在熱分層消退期存在高營養水平及濁度,極可能是由于水庫在上年的秋冬交際時(熱分層消退期與混合期的過渡時期)發生“翻庫現象”(吳豐昌等,2010)。
3.2? ?不同熱分層時期DO分層變化機制
在熱分層時期,水體熱分層是直接影響DO分層規律的最主要因素,浮游植物、營養物質及有機物也是影響DO分層的重要因素,水體濁度間接影響DO變化。水體熱分層造成的密度分層是驅動DO分層的物理原動力,根據相關性分析結果及氧躍層與溫躍層深度、厚度及變化梯度的線性擬合結果,庫區水體熱分層對DO垂向分層規律產生了顯著影響,但同時也受到其他環境因素的影響,這與在密蘇里州水庫及中國千島湖所發現的規律類似(Jones et al,2011;Zhang et al,2015)。
分層期間適宜的水溫及高營養負荷使得浮游植物在表層大量繁殖,強烈的光合作用抵消了O2溶解度因溫度升高而降低的作用,表層的DO達到超飽和。但水體熱分層阻礙DO向下補充,且浮游植物生物量下降削弱光合作用,加以好氧條件下強烈的硝化反應及有機物質堆積分解耗氧驅動導致庫區表層水體在熱分層期間出現了梯度變化大的溫躍層與氧躍層(Yoshimizu et al,2010;Kreling et al,2017)。水體中的浮游植物趨向分布于光合有效輻射強的水層,而水體濁度影響光合有效輻射強度的衰減,進而影響浮游植物的分布及光合作用,并對DO垂向分層規律產生影響(唐詩等,2013;谷嬌等,2016)。熱分層形成期、穩定期及消退期間的水體濁度影響著浮游植物分布(Gallegos,2001;Obrador & Pretus,2008)。加之水體表層溫躍層具有“兼上顧下”特殊的物化條件,浮游生物將自身定位在密度梯度大的空間中以便獲得上下層水體的優異條件,因此表層溫躍層因內、外源O2補充豐富,DO水平較高(Boehrer & Schultze,2008;Yang et al,2018)。
O2垂向梯度擴散受阻和局部原位消耗共同作用導致熱分層形成期出現典型的溫躍層DO極小值現象(MOM)。MOM位置的湍流擴散率隨著水體密度分層強度的增加而降低,DO運輸受垂向梯度約束(Kreling et al,2017)。另外,浮游植物生長繁殖需吸收NH4+,浮游植物生物量下降減少了對NH4+的吸收利用(Tuantet et al,2014;Su et al,2019)。有機物質及死亡藻類分解消耗DO,已有研究證實變溫層有機物質降解及藻類死亡分解對MOM的出現有著巨大貢獻(Zhang et al,2015;Wentzky et al,2019);根據亞硝化反應(式②)及硝化反應(式③):
好氧條件下的硝化反應增強,氧躍層中NO2–及NO3–濃度增加,NH4+向NO2–及NO3–的轉化過程消耗DO,DO的變化規律影響氮的轉化行為,MOM在水層內形成氧化還原梯度,為這些過程提供了好氧―厭氧耦合等適宜的物化條件,加強生物競爭,進一步影響種群結構分布及氮循環過程(Elci,2008;Woolway et al,2014; Su et al,2019;Wentzky et al,2019)。強烈的DO分層甚至影響水體中CO2和CH4的垂直分布以及擴散外流的強度和時間,或許湖庫成為溫室氣體排放的一個來源(McClure et al,2018)。
浮游植物分布趨勢、隨水溫升高而降低的O2溶解度、水體熱分層的阻礙等影響水體DO的內、外源補充,藻類死亡沉降分解、有機物質耗氧分解及NH4+的硝化反應進一步消耗O2,導致水庫在熱分層穩定期出現大范圍缺氧水層,缺氧水層中水體的反硝化反應顯著增強(Liljendahl-Nurminen et al,2008;Rice et al,2009)。DO極大值的出現是由于有機物質耗氧分解削弱,且極大值出現位置因季節變化響應延遲故水層溫度較低,仍保留著熱分層加劇前較高的DO水平,故經好氧硝化產生較高水平的NO3–。熱分層消退期間庫區表層因硝化反應及有機物質耗氧分解強烈,導致DO水平迅速下降(Blair et al,2013;Qu et al,2022)。庫區在熱分層消退期缺氧范圍大、水體濁度低,浮游植物生物量在第2個溫躍層及氧躍層位置突降,缺氧條件下水體反硝化作用顯著增強,硝化作用減弱,NH4+在氧躍層富集(Cole & Pace,1998)。滯溫層在熱分層形成期、穩定期及消退期都出現大范圍缺氧,溫度上升加劇熱分層,大氣向水體的充氧和浮游植物光合作用產氧因水體分層阻礙運輸及消耗而難以補充,藻類死亡沉降、有機物質堆積分解都使得滯溫層的缺氧現象加劇,這對湖庫深水層的生物及底棲生物的生存繁衍造成了巨大的風險,厭氧環境促進沉積物中的營養釋放,進一步惡化水質(Bajer et al,2007;McClure et al,2018)(圖7)。
4? ?結論
(1)萬安水庫在2月為混合期,未出現缺氧現象;2-5月為熱分層形成期,形成溫躍層+混合層+溫躍層+滯溫層的4分層結構;5-8月為熱分層穩定期,溫躍層+混合層+溫躍層+滯溫層的4分層結構進一步穩固;10月為熱分層消退期,形成混合層+變溫層+滯溫層的3分層結構。
(2)萬安水庫混合期的營養水平為貧營養,熱分層形成期及消退期為中營養,熱分層穩定期為中度富營養化。庫區混合期表層DO變化受浮游植物分布影響的同時也受“翻庫”現象影響。高光合速率能克服溶解度下降所帶來的DO損失,故較高營養水平下水體DO的垂向分布規律受多因素耦合影響,熱分層時期DO變化的主導因素包括水體熱分層、營養水平、污染狀態及浮游植物生長分布。而水體濁度通過影響浮游植物的垂向分布,進而影響DO的垂向變化規律。
(3)熱分層形成期的水體在內源產氧減少和原位DO消耗驅動作用下發生MOM。而水體熱分層阻礙DO補充、內源產氧減弱是造成熱分層穩定期水體上層大范圍缺氧的主要因素。
(4)庫區在熱分層形成期、穩定期及消退期皆出現滯溫層缺氧現象,溫度上升加劇熱分層,DO補充阻礙加大,大量水表藻類死亡沉降、有機物質堆積分解使得滯溫層的缺氧現象加劇。
參考文獻
陳東,張麗旭,劉漢奇,等,2008. 長江口海域春夏季溶解氧分布特征及其相關因素分析[J]. 海洋環境科學,108(S1):49-53.
谷嬌,寧曉雨,靳輝,等,2016. 沉積物再懸浮對淺水湖泊沉水植物光合作用及生長影響的研究進展[J]. 生態科學,35(1):200-208.
郭詩君,王小軍,韓品磊,等,2021. 丹江口水庫葉綠素a濃度的時空特征及影響因子分析[J]. 湖泊科學,33(2):366-376.
金相燦,1995. 中國湖泊環境[M]. 北京:海洋出版社.
李秋華,商立海,高廷進,等,2013. 喀斯特高原深水水庫―萬峰湖富營養化特征分析[J]. 生態科學,32(2):194-199.
李艷紅,成靜清,夏麗麗,等,2013. 鄱陽湖區水體溶解氧現狀及環境影響因素分析[J]. 中國農村水利水電,372(10):122-125.
林炳明,2006. 萬安水庫銀魚資源調查與開發[J]. 中國水產,(7):74-75.
呂琳莉,李朝霞,崔崇雨,2018. 高原河流溶解氧變化規律研究[J]. 環境科學與技術,41(7):133-140.
唐詩,孫濤,沈小梅,等,2013. 水體濁度變化影響下的河口溶解氧系統動力學模型及應用[J]. 水利學報,44(11):1286-1294.
吳豐昌,金相燦,張潤宇,等,2010. 論有機氮磷在湖泊水環境中的作用和重要性[J]. 湖泊科學,22(1):1-7.
趙紫涵,宋貴生,趙亮,2020. 秦皇島外海夏季溶解氧與pH的變化特征分析[J]. 海洋學報,42(10):144-154.
鄭瑩瑩,劉樹元,劉虹,等,2021. 亞深型水庫水體季節性分層特征研究[J]. 云南農業大學學報(自然科學),36(2):359-370.
Bajer P G, Millspaugh J J, Hayward R S, 2007. Application of discrete choice models to predict white crappie temperature selection in two Missouri impoundments[J]. Transactions of the American Fisheries Society, 136(4):889-901.
Blair J M, Ostrovsky I, Hicks B J, et al, 2013. Growth of rainbow trout (Oncorhynchus mykiss) in warm-temperate lakes: implications for environmental change[J]. Canadian Journal of Fisheries and Aquatic Sciences, 70(5):815-823.
Boehrer B, Schultze M, 2008. Stratification of lakes[J]. Reviews of Geophysics, 46(2):RG2005.
Borics G, Abonyi A, Varbiro G, et al, 2015. Lake stratification in the Carpathian basin and its interesting biological consequences[J]. Inland Waters, 5(2):173-186.
Caliskan A, Elci S, 2009. Effects of Selective Withdrawal on Hydrodynamics of a Stratified Reservoir[J]. Water Resources Management, 23(7):1257-1273.
Cole J J, Pace M L, 1998. Hydrologic variability of small, Northern Michigan lakes measured by the addition of tracers[J]. Ecosystems, 1(3):310-320.
Demers E, Kalff J, 1993. A Simple-Model for perdicting the date of spring stratification in temperate and subtropical lakes[J]. Limnology and Oceanography, 38(5):1077-1081.
Diaz R, Rosenberg R, 1995. Marine benthic hypoxia: A review of its ecological effects and the behavioural responses of benthic macrofauna[J]. Oceanogr. Mar. Biol. Annu. Rev, 33:245-303.
Elci S, 2008. Effects of thermal stratification and mixing on reservoir water quality[J]. Limnology, 9(2):135-142.
Fernandez J E, Peeters F, Hofmann H, 2014. Importance of the Autumn Overturn and Anoxic Conditions in the Hypolimnion for the Annual Methane Emissions from a Temperate Lake[J]. Environmental Science & Technology, 48(13):7297-7304.
Gallegos C L, 2001. Calculating optical water quality targets to restore and protect submersed aquatic vegetation: Overcoming problems in partitioning the diffuse attenuation coefficient for photosynthetically active radiation[J]. Estuaries, 24(3):381-397.
Harke M J, Steffen M M, Gobler C J, et al, 2016. A review of the global ecology, genomics, and biogeography of the toxic cyanobacterium, Microcystis spp.[J]. Harmful Algae, 54:4-20.
Houser J N, 2006. Water color affects the stratification, surface temperature, heat content, and mean epilimnetic irradiance of small lakes[J]. Canadian Journal of Fisheries and Aquatic Sciences, 63(11):2447-2455.
Huang T, 2016. Water Pollution and Water Quality Control of Selected Chinese Reservoir Basins[M]. Switzerland: Springer International Publishing.
Jane S F, Hansen G J A, Kraemer B M, et al, 2021. Widespread deoxygenation of temperate lakes[J]. Nature, 594:66-70.
Jones J R, Knowlton M F, Obrecht D V, et al, 2011. Temperature and oxygen in Missouri reservoirs[J]. Lake and Reservoir Management, 27(2):173-182.
Kosten S, Huszar V L M, Becares E, et al, 2012. Warmer climates boost cyanobacterial dominance in shallow lakes[J]. Global Change Biology, 18(1):118-126.
Kreling J, Bravidor J, Engelhardt C, et al, 2017. The importance of physical transport and oxygen consumption for the development of a metalimnetic oxygen minimum in a lake[J]. Limnology and Oceanography, 62(1):348-363.
Liljendahl-Nurminen A, Horppila J, Lampert W, 2008. Physiological and visual refuges in a metalimnion: an experimental study of effects of clay turbidity and an oxygen minimum on fish predation[J]. Freshwater Biology, 53(5):945-951.
Matzinger A, Muller B, Niederhauser P, et al, 2010. Hypolimnetic oxygen consumption by sediment-based reduced substances in former eutrophic lakes[J]. Limnology and Oceanography, 55(5):2073-2084.
Mcclure R P, Hamre K D, Niederlehner B R, et al, 2018. Metalimnetic oxygen minima alter the vertical profiles of carbon dioxide and methane in a managed freshwater reservoir[J]. Science of the Total Environment, 636:610-620.
Muller B, Bryant L D, Matzinger A, et al, 2012. Hypolimnetic Oxygen Depletion in Eutrophic Lakes[J]. Environmental Science & Technology, 46(18):9964-9971.
North R P, North R L, Livingstone D M, et al, 2014. Long-term changes in hypoxia and soluble reactive phosphorus in the hypolimnion of a large temperate lake: consequences of a climate regime shift[J]. Global Change Biology, 20(3):811-823.
Obrador B, Pretus J L, 2008. Light regime and components of turbidity in a Mediterranean coastal lagoon[J]. Estuarine, Coastal and Shelf Science, 77(1):123-133.
Qu L Y, He C, Wu Z T, et al, 2022. Hypolimnetic deoxygenation enhanced production and export of recalcitrant dissolved organic matter in a large stratified reservoir[J]. Water Research, 219:118537.
Rice J, Sykes J, Barwick H, et al. 2009. The role of metalimnetic oxygen minimum zones in striped bass summer kills and forage fish entrainment: consequences and management implications[C]//139th Annual Meeting of the American Fisheries Society. Nashvile: 121-145.
Saito L, Johnson B M, Bartholow J, et al, 2001. Assessing ecosystem effects of reservoir operations using food web-energy transfer and water quality models[J]. Ecosystems, 4(2):105-125.
Schindler D E, 2017. Warmer climate squeezes aquatic predators out of their preferred habitat[J]. Proceedings of the National Academy of Sciences of the United States of America, 114(37):9764-9765.
Singleton V L, Little J C, 2006. Designing hypolimnetic aeration and oxygenation systems - A review[J]. Environmental Science & Technology, 40(24):7512-7520.
Su X X, He Q, Mao Y F, et al, 2019. Dissolved oxygen stratification changes nitrogen speciation and transformation in a stratified lake[J]. Environmental Science and Pollution Research, 26(3):2898-2907.
Torres E, Galvan L, Canovas C R, et al, 2016. Oxycline formation induced by Fe(II) oxidation in a water reservoir affected by acid mine drainage modeled using a 2D hydrodynamic and water quality model-CE-QUAL-W2[J]. Science of the Total Environment, 562:1-12.
Tuantet K, Temmink H, Zeeman G, et al, 2014. Nutrient removal and microalgal biomass production on urine in a short light-path photobioreactor[J]. Water Research, 55:162-174.
Wagner C, Adrian R, 2011. Consequences of changes in thermal regime for plankton diversity and trait composition in a polymictic lake: a matter of temporal scale[J]. Freshwater Biology, 56(10):1949-1961.
Wentzky V C, Frassl M A, Rinke K, et al, 2019. Metalimnetic oxygen minimum and the presence of Planktothrix rubescens in a low-nutrient drinking water reservoir[J]. Water Research, 148:208-218.
Wilhelm S, Adrian R, 2008. Impact of summer warming on the thermal characteristics of a polymictic lake and consequences for oxygen, nutrients and phytoplankton[J]. Freshwater Biology, 53(2):226-237.
Woolway R I, Maberly S C, Jones I D, et al. 2014. A novel method for estimating the onset of thermal stratification in lakes from surface water measurements[J]. Water Resources Research, 50(6):5131-5140.
Woolway R I, Merchant C J, 2019. Worldwide alteration of lake mixing regimes in response to climate change[J]. Nature Geoscience, 12(4):271-276.
Yang Y, Wang Y, Zhang Z, et al, 2018. Diurnal and Seasonal Variations of Thermal Stratification and Vertical Mixing in a Shallow Fresh Water Lake[J]. Journal of Meteorological Research, 32(2):219-232.
Yoshimizu C, Yoshiyama K, Tayasu I, et al, 2010. Vulnerability of a large monomictic lake (Lake Biwa) to warm winter event[J]. Limnology, 11(3):233-239.
Yvon-Durocher G, Jones J I, Trimmer M, et al, 2010. Warming alters the metabolic balance of ecosystems[J]. Philosophical Transactions of the Royal Society B-Biological Sciences, 365(1549):2117-2126.
Zhang Y L, Wu Z X, Liu M L, et al, 2015. Dissolved oxygen stratification and response to thermal structure and long-term climate change in a large and deep subtropical reservoir (Lake Qiandaohu, China)[J]. Water Research, 75:249-258.
(責任編輯? ?熊美華)
收稿日期:2023-08-12? ? ? 修回日期:2023-12-15
基金項目:福建省水利科技項目(MSK202216);福建省環保科技計劃項目(2021R023);福建省廈門環境監測中心站環保科技項目(2022S01)。
作者簡介:陳侃,2000年,男,碩士研究生,河流環境過程方向。E-mail:chenkan@stu.xmu.edu.cn
通信作者:曹文志,1968年,男,教授,主要從事河流與近海生物地球化學過程、水文與水資源。E-mail:wzcao@xmu.edu.cn
