丹參LBD基因家族的特性鑒定與抗熱性分析
2024-05-17 06:59:34李萬(wàn)樊昕華
商洛學(xué)院學(xué)報(bào) 2024年2期
李萬(wàn) 樊昕華
摘 要:利用生物信息學(xué)方法,對(duì)丹參LBD(later organ boundaries domain)家族成員(SmLBDs)進(jìn)行了系統(tǒng)的篩選和鑒定,并分析了其在高溫脅迫(37 °C)下的表達(dá)模式。結(jié)果表明,丹參中共有52個(gè)SmLBDs,可分為groupI、groupII、groupIII、groupIV、groupV、groupVI和groupVII 共7組。52個(gè)SmLBDs編碼蛋白的氨基酸殘基數(shù)為103~298,相對(duì)分子質(zhì)量為11.29~31.72 kDa,等電點(diǎn)分布于4.44~10.71,均為親水性蛋白,且大多定位在細(xì)胞核中。此外,分析52個(gè)SmLBDs在高溫脅迫(37 °C)下的表達(dá)模式發(fā)現(xiàn),7個(gè)基因的表達(dá)水平下降,37個(gè)基因的表達(dá)水平上升,尤其是SmLBD8、SmLBD40和SmLBD44的表達(dá)水平顯著提高,表明這些基因可能與丹參的抗熱性相關(guān)。
關(guān)鍵詞:丹參;LBD家族;亞細(xì)胞定位;高溫脅迫
中圖分類號(hào):S567.23? ? ?文獻(xiàn)標(biāo)識(shí)碼:A? ? ?文章編號(hào):1674-0033(2024)02-0066-09
引用格式:李萬(wàn),樊昕華.丹參LBD基因家族的特性鑒定與抗熱性分析[J].商洛學(xué)院學(xué)報(bào),2024,38(2):66-74.
Genome-wide Identification and Analysis of LBD
Gene Family in Salvia miltiorrhiza
LI Wan, FAN Xin-hua
(School of Biopharmaceutical and Food Engineering, Shangluo University, Shangluo? 726000, Shaanxi)
Abstract: The members of the later organ boundaries domain (LBD) family of Salvia miltiorrhia LBD (SmLBDs) were systematically screened and identified using bioinformatics methods, and their expression patterns were analyzed under high temperature stress (37 °C). The results showed that there were 52 SmLBDs in Salvia miltiorrhiza, which could be divided into 7 groups: groupI, groupII, groupIII, groupIV, groupV, groupVI and groupVII. The amino acid residues of the 52 proteins encoded by SmLBDs ranged from 103 to 298, the relative molecular weights ranged from 11.29 to 31.72 kDa, and the isoelectric points were distributed from 4.44 to 10.71. All of 52 SmLBDs were hydrophilic proteins, and most of them were located in the nucleus. In addition, the expression patterns of 52 SmLBDs under high temperature stress (37 °C) were analyzed, and the expression levels of 7 genes were decreased, and 37 genes were increased, especially the expression level of SmLBD8, SmLBD40 and SmLBD44 was significantly increased, indicating that these genes may be related to the heat resistance of Salvia miltiorrhiza.
Key words: Salvia miltiorrhiza; LBD family; subcellular localization; high temperature stress
丹參(Salvia miltiorrhiza)為唇形科鼠尾草屬多年生草本植物,是一種傳統(tǒng)中藥材,擁有悠久的歷史,并被廣泛應(yīng)用。目前,由于丹參的市場(chǎng)需求量越來(lái)越大、野生丹參生境破壞加劇,野生丹參的產(chǎn)量已無(wú)法滿足市場(chǎng)需要。同時(shí),人工種植丹參存在抗性差、品質(zhì)退化等問(wèn)題,對(duì)丹參的產(chǎn)量和品質(zhì)造成了極為嚴(yán)重的影響。因此,培育具有較強(qiáng)抗性的丹參已成為近年來(lái)的研究熱點(diǎn)[1]。研究發(fā)現(xiàn),側(cè)生器官邊界結(jié)構(gòu)域(later organ boundaries domain,LBD)也被稱為the asymmetric leaves2-like(ASL)基因家族,根據(jù)蛋白序列的不同可將LBD家族成員分為I類和II類[2-3],I類包含一個(gè)類似鋅指結(jié)構(gòu)域CX2CX6CX3C的基序[4],一個(gè)類似亮氨酸拉鏈結(jié)構(gòu)域的甘氨酸-丙氨酸-絲氨酸(GAS-block)區(qū)域,以及一個(gè)蛋白質(zhì)二聚體LX6LX3LX6L螺旋卷曲結(jié)構(gòu);II類只包含鋅指結(jié)構(gòu)CX2CX6CX3C[4]。研究表明,CX2CX6CX3C在LBD蛋白與DNA的結(jié)合中起重要作用,GAS-block和LX6LX3LX6L參與了LBD蛋白與其他蛋白的相互作用。例如,LBD與bHLH蛋白的相互作用可以降低CX2CX6CX3C對(duì)DNA的親和力[2,5]。自從在擬南芥(Arabidopsis thaliana)中發(fā)現(xiàn)43個(gè)LBD蛋白以來(lái),許多LBD蛋白已在水稻(Oryza sativa)、蘋(píng)果(Malus domestica)等植物中被鑒定[6]。擬南芥AtLOB(AtASL4)基因主要表達(dá)在外側(cè)組織的近端,可與SHOOTMERISTEMLESS(STM)蛋白和BREVIPEDICELLUS(BP)蛋白結(jié)合,調(diào)控幼葉的發(fā)育[4]。AtLBD41(AtASL38)在甜瓜(Cucumis melo)中的過(guò)表達(dá)可導(dǎo)致葉皺畸形,并具有明顯的初始化現(xiàn)象。因此,推測(cè)它可能參與調(diào)控葉片的近軸極性和遠(yuǎn)軸極性[7]。AtLBD6(AtAS2)是水稻OsAS2的同源基因,其表達(dá)產(chǎn)物能夠抑制葉片近軸區(qū)的細(xì)胞增殖和近遠(yuǎn)軸對(duì)稱,與AS1和JAG協(xié)同形成扁平葉片,同時(shí)可參與水稻芽分化的調(diào)控和不同輪的邊界表達(dá),表明該基因可能調(diào)控花發(fā)育[8-10]。LBD10蛋白在擬南芥花粉發(fā)育過(guò)程中起關(guān)鍵作用[11]。在水稻中,OsIG1參與水稻的花器官數(shù)量和配子體發(fā)生[12]。OsLBD37和OsLBD38的過(guò)表達(dá)可以延緩水稻抽穗過(guò)程和速度,提高水稻產(chǎn)量[13]。蘋(píng)果MdLBD13蛋白能抑制蘋(píng)果中花青素的合成和氮的吸收[14]。玉米(Zea mays)ZmIG1調(diào)控雌配子的發(fā)育和葉片軸向分化[15]。桉樹(shù)(Eucalyptus)EgLBD37基因的過(guò)表達(dá)使桉樹(shù)的節(jié)間長(zhǎng)度增加,植株較高,木質(zhì)素木質(zhì)化組分增加。EgLBD29的過(guò)表達(dá)使纖維的長(zhǎng)度縮短,縮短了植物的循環(huán)生長(zhǎng)。EgLBD22的過(guò)表達(dá)可增加韌皮部的纖維含量[16]。用10% PEG處理24 h后,葡萄(Vitis vinifera)VvLBD19表達(dá)上調(diào),是對(duì)照的26倍[17]。研究表明,丹參易受不良環(huán)境影響,導(dǎo)致產(chǎn)量下降和品質(zhì)降低,尤其是高溫對(duì)丹參有效成分的積累十分不利[18-19]。因此,本文擬利用生物信息學(xué)和分子生物研究方法,從丹參中鑒定LBD基因家族成員(SmLBDs),通過(guò)分析其性質(zhì)、基序(motif)組成、系統(tǒng)發(fā)育、亞細(xì)胞定位等,以及SmLBDs在高溫脅迫下(37 ℃)的表達(dá)模式,篩選出對(duì)高溫有明顯響應(yīng)的候選基因資源,為改善丹參對(duì)高溫脅迫的抗性提供參考和理論依據(jù),從而為提高丹參的產(chǎn)量和品質(zhì)提供實(shí)驗(yàn)基礎(chǔ)和數(shù)據(jù)支撐。
1? 材料與方法
1.1 植物材料及生長(zhǎng)條件
本研究使用的丹參植株由陜西天士力植物藥業(yè)有限責(zé)任公司提供。將丹參植株移栽至10 cm×10 cm的方盆中(含有營(yíng)養(yǎng)土和蛭石),置于人工氣候箱中培養(yǎng)(20~22 °C,4 000 lx,16 h光照/8 h黑暗,濕度70%)。兩周后,將長(zhǎng)勢(shì)相近的幼苗分為兩組,第一組在37 °C下培養(yǎng)24 h(試驗(yàn)組),第二組在正常環(huán)境下培養(yǎng)24 h(對(duì)照組),隨后采集相類部位的根、莖和葉組織各200 mg,混合后置于液氮中快速冷凍,并在-80 °C保存,待用。
1.2 丹參中LBD基因的鑒定及蛋白序列分析
為鑒定SmLBDs,通過(guò)本地Blast程序,利用TAIR數(shù)據(jù)庫(kù)下載的46個(gè)AtLBDs蛋白序列,對(duì)丹參蛋白數(shù)據(jù)庫(kù)中的SmLBDs(E<1×10-10)進(jìn)行匹配和篩選。對(duì)獲得的候選SmLBDs進(jìn)行CDD(conserved domain database,https://www.ncbi.nlm.nih.gov/cdd/)分析,去除重復(fù)序列和錯(cuò)誤序列,其余為丹參SmLBDs。利用MEGA7軟件構(gòu)建了丹參和擬南芥LBD蛋白的系統(tǒng)發(fā)育樹(shù)(NJ、Jones- Taylor-Thornton JTT模型、1 000個(gè)重復(fù)和根值分析)。用MEME(http://meme-suite.org/)分析LBD蛋白的保守基序,基序數(shù)目設(shè)置為15個(gè)。
1.3 跨膜結(jié)構(gòu)域和信號(hào)肽分析及亞細(xì)胞定位
利用ProtParam網(wǎng)站(http://us.expasy.org/tools/protparam.html)預(yù)測(cè)蛋白質(zhì)性質(zhì),通過(guò)TOPCONS網(wǎng)站(https://topcons.cbr.su.se/)預(yù)測(cè)跨膜結(jié)構(gòu)域和信號(hào)肽,使用CELLO網(wǎng)站(http://cello.life.nctu.edu.tw/)預(yù)測(cè)亞細(xì)胞定位。
1.4 RNA提取及基因表達(dá)的實(shí)時(shí)RT-PCR分析
使用天根生化科技(北京)有限公司總RNA提取試劑盒(DP419)提取總RNA;應(yīng)用寶日醫(yī)生物技術(shù)(北京)有限公司(takara中國(guó))的反轉(zhuǎn)錄試劑盒PrimeScript RT reagent Kit(RR047A)合成cDNA;利用北京百泰克生物技術(shù)有限公司的熒光定量試劑2×Plus SYBR real-time PCR mixture(PR7702)實(shí)行熒光定量PCR(qPCR)分析。數(shù)據(jù)分析采用2-ΔΔct法進(jìn)行分析[20]。
2? 結(jié)果與分析
2.1 丹參LBD基因家族成員的鑒定
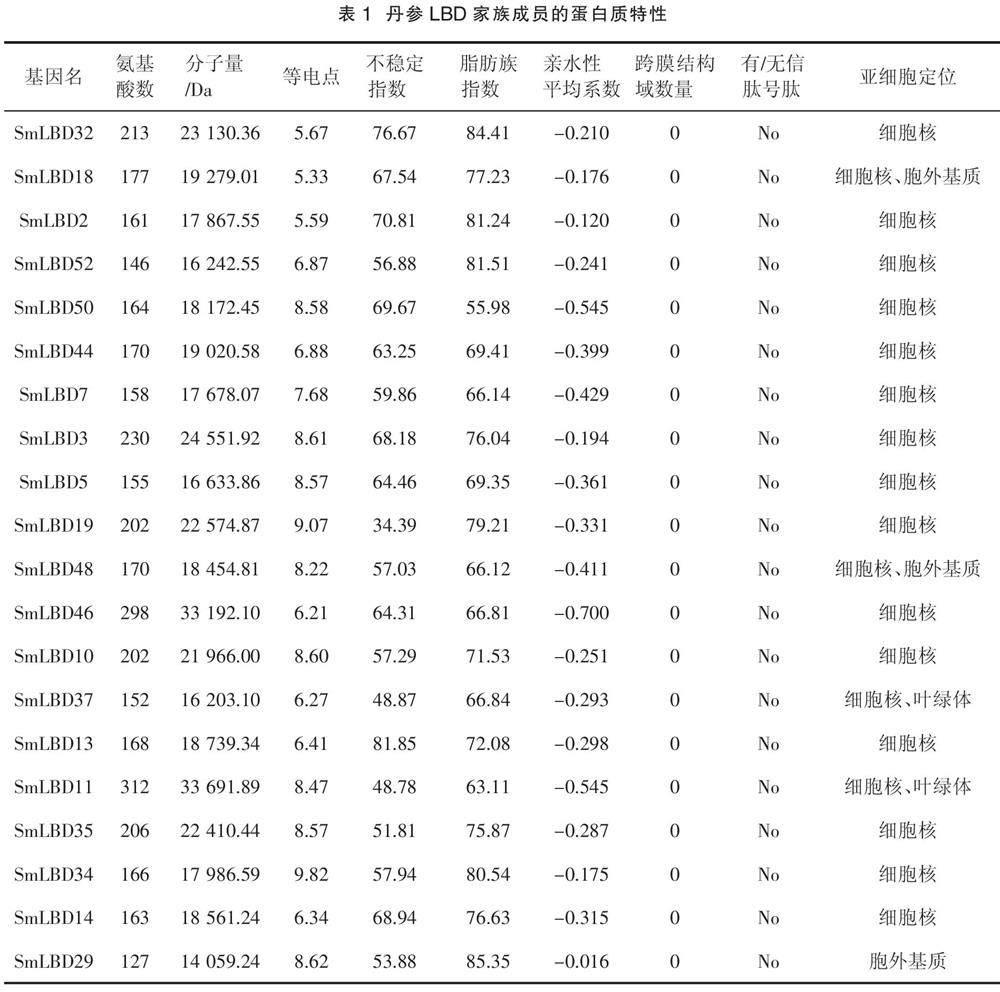
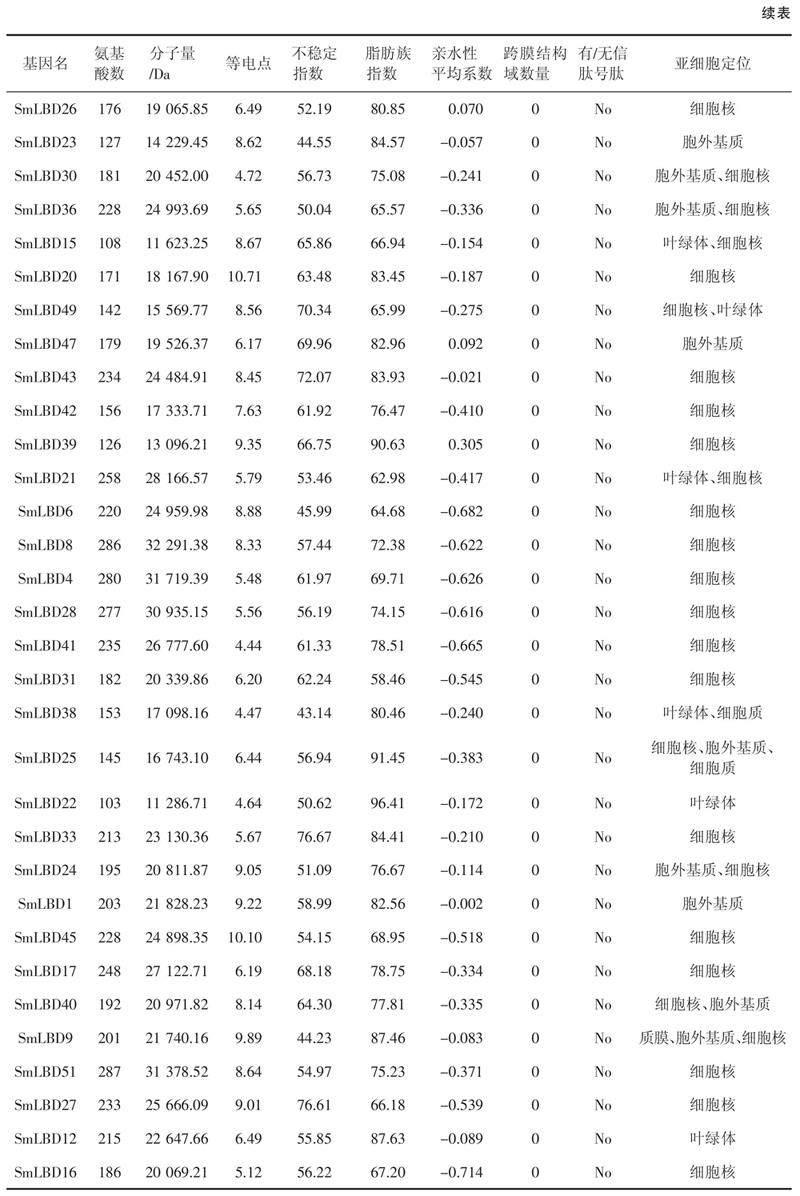
本研究鑒定了52個(gè)SmLBDs蛋白,如表1所示,SmLBD11是最大的蛋白,含有312個(gè)氨基酸殘基,分子量為33.7 kDa。最小的蛋白是SmLBD22,只有103個(gè)氨基酸殘基,分子量為11.3 kDa。52個(gè)SmLBDs蛋白的跨膜結(jié)構(gòu)域數(shù)均為0,且沒(méi)有信號(hào)肽,等電點(diǎn)范圍為4.44(SmLBD41)~10.71(SmLBD20),親水性平均系數(shù)(Grand average of hydropathicity,GRAVY)均小于0,范圍為- 0.714(SmLBD16)~ - 0.002(SmLBD1),不穩(wěn)定指數(shù)在34.39(SmLBD19)~81.85(SmLBD13),脂肪族指數(shù)為55.98(SmLBD50)~96.41(SmLBD22)。亞細(xì)胞定位分析表明,大部分基因位于細(xì)胞核中,而SmLBD18、SmLBD48、SmLBD24和SmLBD40存在于細(xì)胞核和胞外基質(zhì)中,SmLBD37、SmLBD11、SmLBD15、SmLBD49和SmLBD21出現(xiàn)在細(xì)胞核和葉綠體中,SmLBD29、SmLBD23、SmLBD47和SmLBD1出現(xiàn)在胞外基質(zhì)中,SmLBD9出現(xiàn)在細(xì)胞核、質(zhì)膜和胞外基質(zhì)中。
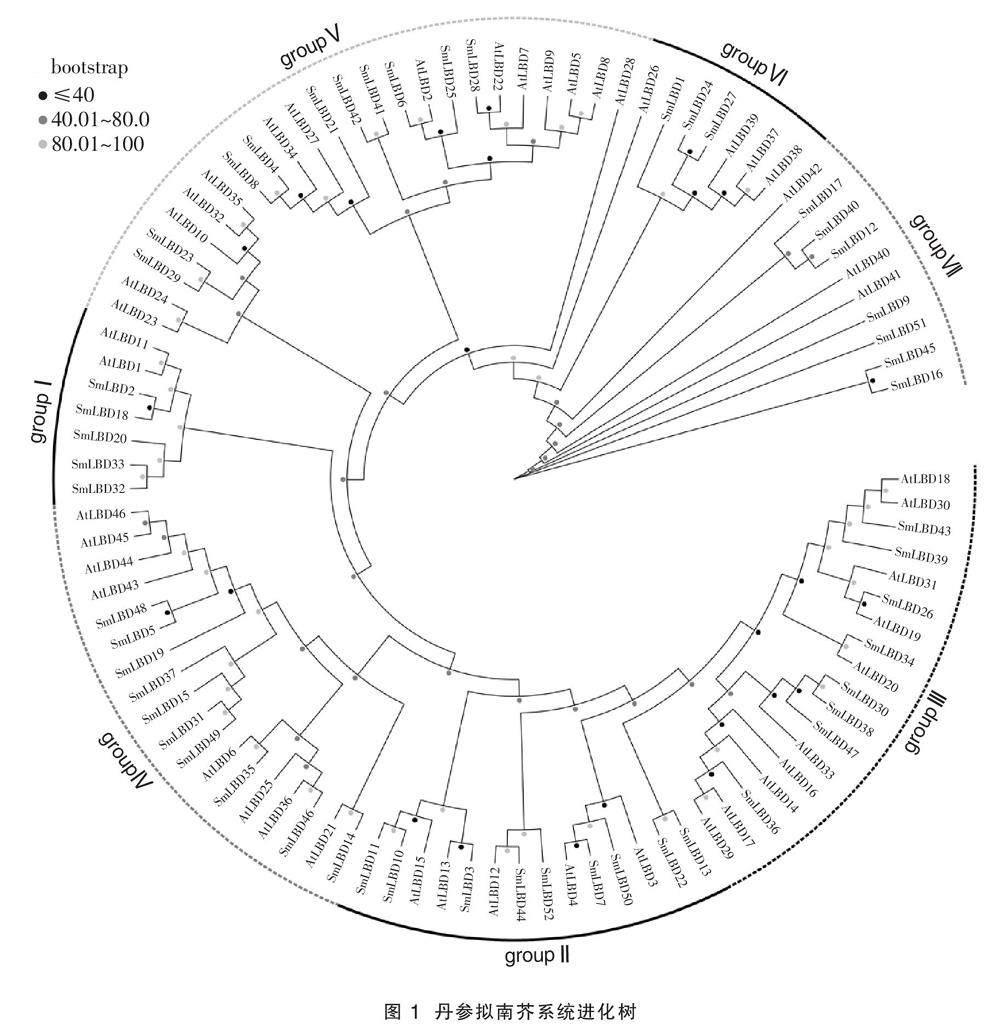
2.2 丹參LBD基因家族的系統(tǒng)進(jìn)化分析
構(gòu)建SmLBDs和AtLBDs的進(jìn)化樹(shù),如圖1所示,52個(gè)SmLBDs基因被分為groupI(包含5個(gè)成員)、groupII(包含9個(gè)成員)、groupIII(包含8個(gè)成員)、groupIV(包含10個(gè)成員)、groupV(包含10個(gè)成員)、groupVI(包含3個(gè)成員)和groupVII(包含7個(gè)成員),32個(gè)SmLBDs和AtLBDs的進(jìn)化樹(shù)組成16對(duì)同源基因,其中12對(duì)同源基因的bootstrap值高于80,分別為SmLBD52/SmLBD44、
SmLBD50/SmLBD7、SmLBD10/SmLBD11、SmLBD13/SmLBD22、SmLBD30/SmLBD38、SmLBD46/SmLBD35、SmLBD5/SmLBD48、SmLBD49/SmLBD31、SmLBD42/SmLBD41、SmLBD8/SmLBD4、SmLBD29/SmLBD23、SmLBD24/SmLBD27。
2.3 SmLBDs的保守基序分析
使用MEME(http://meme-suite.org/)在線工具鑒定了15個(gè)保守基序,如圖2所示,groupII、gruopIII、groupIV和groupV的大部分SmLBDs都包含基序1,2,3和4。含有基序數(shù)量最多的7個(gè)基因分別是SmLBD4、SmLBD8、SmLBD10、SmLBD11、SmLBD32、SmLBD33和SmLBD45,分布在groupI、groupII、groupV和groupVII,共有11個(gè)基序,分別為基序1,2,3,4,5,7,9,10,11,14和15。含有基序數(shù)量最少的基因是groupVI的SmLBD27,只有2個(gè)基序,基序1和5。此外,groupVI只包含3個(gè)基因:SmLBD1,SmLBD24和SmLBD27,均包含基序1,2,5和11;除了SmLBD16僅含有基序7和11之外,其它51個(gè)基因都含有基序1;基序10只存在于SmLBD32和SmLBD33。
2.4 SmLBDs在高溫條件下的表達(dá)分析
圖3是通過(guò)檢測(cè)SmLBDs在高溫脅迫下的表達(dá)模式圖,從圖3中發(fā)現(xiàn),SmLBD5、SmLBD6、SmLBD18、SmLBD19、SmLBD23、SmLBD28和
SmLBD36的基因表達(dá)量呈下降趨勢(shì);SmLBD1、SmLBD3、SmLBD9、SmLBD13、SmLBD26、SmLBD33、SmLBD35和SmLBD37這8個(gè)基因的表達(dá)量均無(wú)顯著變化;其它37個(gè)基因的表達(dá)量呈上升趨勢(shì),尤其是SmLBD8、SmLBD40和SmLBD44的表達(dá)量升高了20倍以上,表明這三個(gè)基因?qū)Ω邷乇容^敏感,可能是丹參抵御高溫脅迫的關(guān)鍵基因。
3? 討論與結(jié)論
LBD蛋白家族是一種植物特異性轉(zhuǎn)錄因子,在植物生長(zhǎng)發(fā)育和逆境脅迫中發(fā)揮重要作用。本研究在丹參中發(fā)現(xiàn)了52個(gè)SmLBDs蛋白,并對(duì)其進(jìn)行了系統(tǒng)分析。等電點(diǎn)范圍為4.44~10.71,其中25個(gè)SmLBDs的等電點(diǎn)小于7,為酸性蛋白,27個(gè)SmLBDs大于7,為堿性蛋白。跨膜結(jié)構(gòu)域和信號(hào)肽是分泌蛋白的標(biāo)志[21],52個(gè)SmLBDs蛋白均不含有跨膜結(jié)構(gòu)域和信號(hào)肽,即均為非分泌蛋白。GRAVY值大于0表明蛋白疏水,小于0表明蛋白質(zhì)親水,丹參SmLBDs的GRAVY值均小于0,表明均為親水蛋白。脂肪族指數(shù)是衡量蛋白溶解性、穩(wěn)定性和抗氧化性等特性的指標(biāo),值越大,說(shuō)明該蛋白的溶解性和穩(wěn)定性越好。丹參52個(gè)SmLBDs的脂肪族系數(shù)均在55.98以上,最大為96.41,表明這些蛋白具有較好的溶解性和穩(wěn)定性。不穩(wěn)定指數(shù)用來(lái)衡量目的蛋白在體外的穩(wěn)定性,值小于40表明蛋白質(zhì)較為穩(wěn)定[22]。本研究發(fā)現(xiàn),51個(gè)SmLBDs的不穩(wěn)定指數(shù)大于40,說(shuō)明蛋白性質(zhì)不穩(wěn)定,而SmLBD19的不穩(wěn)定指數(shù)為34.39,即蛋白性質(zhì)穩(wěn)定。轉(zhuǎn)錄因子通常在細(xì)胞核中表達(dá)發(fā)揮作用,因此通過(guò)亞細(xì)胞定位分析發(fā)現(xiàn),大部分SmLBDs定位于細(xì)胞核中,這也是其不含跨膜結(jié)構(gòu)域和信號(hào)肽的原因。然而,有一部分SmLBDs定位在胞外基質(zhì)、葉綠體和質(zhì)膜等位置,表明這些蛋白可能具有其他的特殊功能,還需要進(jìn)行深入研究加以確定。
Yamasaki等[23]研究認(rèn)為保守基序在活性蛋白中發(fā)揮了功能或結(jié)構(gòu)作用。因此,分析SmLBDs的motif組成有利于推測(cè)基因功能。此外,為了了解某個(gè)基因的功能,通過(guò)對(duì)與該基因相似度或同源性較高的其他基因的功能進(jìn)行預(yù)測(cè)是一種較為常用的方法。例如,擬南芥AtLBD16、AtLBD17、AtLBD18和AtLBD29能夠誘導(dǎo)愈傷組織形成,參與植株再生,根據(jù)進(jìn)化樹(shù)分布,位于同一分支的丹參SmLBD26、SmLBD30、SmLBD34、SmLBD36、
SmLBD38、SmLBD39、SmLBD43和SmLBD47應(yīng)該具有類似功能[6]。AtLBD16、AtLBD18和AtLBD29主要在側(cè)根中表達(dá),可能和擬南芥根發(fā)育有關(guān),丹參以根入藥,因此,通過(guò)調(diào)控SmLBD36和SmLBD43等同源基因的表達(dá),可能能夠促進(jìn)丹參根的發(fā)育,提高產(chǎn)量和品質(zhì)[22]。AtLBD6與擬南芥的花和葉的發(fā)育有關(guān)[8-10],過(guò)表達(dá)AtLBD1和AtLBD11,可使植株長(zhǎng)高,并增加節(jié)間長(zhǎng)度[16],因此,可推測(cè)丹參中SmLBD35和SmLBD2等基因也有相似功能。其它SmLBDs的基因功能,可以依據(jù)已有的AtLBDs基因功能的研究結(jié)果進(jìn)行分析預(yù)測(cè),或者通過(guò)構(gòu)建丹參和其他物種LBD基因的系統(tǒng)進(jìn)化樹(shù)進(jìn)行預(yù)測(cè)。
LBD家族與植物對(duì)非生物脅迫的抗性有關(guān),例如,葡萄VvLBD19的表達(dá)量在PEG處理后顯著上調(diào)。在干旱脅迫下,馬鈴薯StLBD1-5和StLBD2-6的表達(dá)量顯著下調(diào),而StLBD2-6和StLBD3-5的表達(dá)量顯著上調(diào),表明這些基因可能有助于維持馬鈴薯的正常代謝,增強(qiáng)馬鈴薯的抗旱性[6,17]。然而,在丹參等藥用植物中關(guān)于LBD與耐熱性關(guān)系的研究鮮有報(bào)道。本研究通過(guò)分析高溫脅迫下丹參52個(gè)SmLBDs的表達(dá)水平發(fā)現(xiàn),44個(gè)SmLBDs可能與丹參的耐熱性相關(guān),尤其是SmLBD8、SmLBD40和SmLBD44的表達(dá)量在高溫脅迫下極顯著升高,表明這3個(gè)基因在丹參抵御高溫脅迫中可能具有關(guān)鍵作用,可作為提高丹參耐熱性的候選基因資源。
綜上所述,本研究通過(guò)對(duì)丹參LBD基因家族進(jìn)行系統(tǒng)的篩選和鑒定,并在高溫脅迫下分析SmLBDs的表達(dá)模式,篩選出了3個(gè)可能與丹參耐熱性相關(guān)的關(guān)鍵基因,為培育具有優(yōu)良抗逆性的丹參品種,提高丹參產(chǎn)量和品質(zhì)提供了試驗(yàn)基礎(chǔ)和理論依據(jù)。
參考文獻(xiàn):
[1]? 陳塵,韓立敏,化文平,等.丹參DHAR家族基因的鑒定及表達(dá)模式分析[J].園藝學(xué)報(bào),2020,47(11):2181-2193.
[2]? MATSUMURA Y, IWAKAWA H, MACHIDA Y, et al. Characterization of genes in the Asymmetric Leaves2/Lateral Organ Boundaries (AS2/LOB) family in Arabidopsis thaliana, and functional and molecular comparisons between AS2 and other family members[J].Plant J,2009,58(3):525-537.
[3]? IWAKAWA H, UENO Y, SEMIARTI E, et al. The ASYMMETRIC LEAVES2 gene of Arabidopsis thaliana, required for formation of a symmetric flat leaf lamina, encodes a member of a novel family of proteins characterized by cysteine repeats and a leucine zipper[J].Plant Cell Physiol,2002,43(5):467-478.
[4]? SHUAI B, REYNAGA-PENA C G, SPRINGER P S. The lateral organ boundaries gene defines a novel, plant-specific gene family[J].Plant Physiol,2002,129(2):747-761.
[5]? MA H, ZHAO J. Genome-wide identification, classification, and expression analysis of the arabinogalactan protein gene family in rice (Oryza sativa L.)[J].J Exp Bot,2010,61(10):2647-2668.
[6]? LIU H, CAO M, CHEN X L, et al. Genome-wide analysis of the Lateral Organ Boundaries Domain (LBD) gene family in Solanum tuberosum[J].Int J Mol Sci,2019,20(21):5360.
[7]? MENG L S, LIU H L, CUI X, et al. ASYMMETRIC LEAVES2-LIKE38 gene, a Member of AS2/LOB family of Arabidopsis, causes leaf dorsoventral alternation in transgenic cockscomb plants[J].Acta Physiol Plant,2009,
31(6):1301-1306.
[8]? SEMIARTI E, UENO Y, TSUKAYA H, et al. The ASYMMETRIC LEAVES2 gene of Arabidopsis thaliana regulates formation of a symmetric lamina, establishment of venation and repression of meristem-related homeobox genes in leaves[J].Development,2001,128(10):1771-1783.
[9]? IWAKAWA H, IWASAKI M, KOJIMA S, et al. Expression of the ASYMMETRIC LEAVES2 gene in the adaxial domain of Arabidopsis leaves represses cell proliferation in this domain and is critical for the development of properly expanded leaves[J].Plant J,2007,
51(2):173-184.
[10] LUO J H, WENG L, LUO D. Isolation and expression patterns of LATERAL ORGAN BOUNDARIES-like genes
in Lotus japonicus[J].J Plant Physiol Mol Biol,2006,32(2):202-208.
[11] KIM M J, KIM M, KIM J. Combinatorial interactions between LBD10 and LBD27 are essential for male gametophyte development in Arabidopsis[J].Plant Signal Behav,2015,10(8):1-3.
[12] ZHANG J R, TANG W, HUANG Y L, et al. Down-regulation of a LBD-like gene, OsIG1, leads to occurrence of unusual
double ovules and developmental abnormalities of various floral organs and megagametophyte in rice[J].J Exp Bot,2015,66(1):99-112.
[13] LI C, ZHU S, ZHANG H, et al. OsLBD37 and OsLBD38, two class II type LBD proteins, are involved in the regulation of heading date by controlling the expression of Ehd1 in rice[J].Biochem Biophys Res Commun,2017,486(3):720-725.
[14]? ?LI H H, LIU X, AN J P, et al. Cloning and elucidation of the functional role of apple MdLBD13 in anthocyanin
biosynthesis and nitrate assimilation[J].Plant Cell Tissue Organ Cult,2017,130(1):47-59.
[15]? ?EVANS M M. The indeterminate gametophyte1 gene of maize encodes a LOB domain protein required for embryo Sac and leaf development[J].Plant Cell,2007,19(1):46-62.
[16]? LU Q, SHAO F J, MACMILLAN C, et al. Genomewide analysis of the lateral organ boundaries domain gene family in Eucalyptus grandis reveals members that differentially impact secondary growth[J].Plant Biotechnol J,2018,16(1):124-136.
[17]? ? HE H H, MA Z H, ZHANG Y X, et al. Identification and Expression analysis of LBD gene family in Grape[J].Sci Agric Sin,2018,51(21):4102-4118.
[18]? 方鑫,楊崢,王晨旭,等.丹參DnaJ基因的鑒定及脅迫響應(yīng)分析[J].南京農(nóng)業(yè)大學(xué)學(xué)報(bào),2022,45(1):94-102.
[19]? 賀玉林.丹參有效成分的積累及其與生態(tài)因子的關(guān)系[D].北京:中國(guó)協(xié)和醫(yī)科大學(xué),2007:59-74.
[20]? LI W, CHENG M, ZHAO Y P. Identification and characterization of potassium (K+) transporters in potato (Solanum tuberosum L.) [J].Taiwania,2022,67(3):422-434.
[21]? 李欣宇,徐啟宇,衛(wèi)宇翔,等.高粱LKR基因結(jié)構(gòu)及在種子發(fā)育中的表達(dá)特征分析[J].山西農(nóng)業(yè)科學(xué),2022(12):1591-1598.
[22]? 劉俊,李龍,陳玉龍,等.杜仲LBD基因家族全基因組鑒定與進(jìn)化表達(dá)分析[J].中草藥,2022,53(10):3142-3155.
[23] YAMASAKI K, KIGAWA T, SEKI M, et al. DNA-binding domains of plant-specific transcription factors: structure, function, and evolution[J].Trends Plant Sci,2013,
18(5):267-276.
