基于高通量測序的海州灣沉積物中微生物多樣性分析
2024-05-13 00:00:00顧穎伏光輝王亞東葉仁智盧璐孫苗苗
南方農(nóng)業(yè)·上旬 2024年2期
摘 要 為探究海州灣淺海域沉積物中微生物群落結(jié)構(gòu)的組成及差異性,選取6個點位的沉積物樣品,采用高通量測序技術(shù)對沉積物微生物群落結(jié)構(gòu)的組成進行分析。結(jié)果表明,海州灣6個點位沉積物樣品中微生物群落結(jié)構(gòu)組成存在差異。6個采樣點共發(fā)現(xiàn)2 851個OUTs,分屬于31門74綱116目199科331屬;在門水平上,海州灣水域沉積物的主要優(yōu)勢菌群為變形菌門(Proteobacteria)、擬桿菌門(Bacteroidetes)、酸桿菌門(Acidobacteria)、綠彎菌門(Chloroflexi)、放線菌門(Actinobacteria)、浮霉菌門(Planctomycetes)、疣微菌門(Verrucomicrobia)和硬壁菌門(Firmicutes)等。其中,6個點位沉積物樣品中微生物群落豐度最高的優(yōu)勢菌門均為變形菌門。共現(xiàn)網(wǎng)絡(luò)分析結(jié)果表明,變形菌門、綠彎菌門、酸桿菌門和放線菌門是對共現(xiàn)網(wǎng)絡(luò)結(jié)構(gòu)有重要作用的中心節(jié)點微生物,在群落結(jié)構(gòu)和多樣組成中具有重要作用。此外, FAPROTAX功能預(yù)測發(fā)現(xiàn)海州灣沉積物中微生物主要參與硫循環(huán)、碳循環(huán)和氮循環(huán)等海洋生物地球化學(xué)過程。
關(guān)鍵詞 沉積物;微生物多樣性;高通量測序;海州灣
中圖分類號:X172 文獻標志碼:A DOI:10.19415/j.cnki.1673-890x.2024.03.001
海州灣是位于江蘇省連云港市沿岸的開放型淺海海灣,其底質(zhì)類型豐富、入海河流眾多、生物餌料豐富,是我國重要的漁場之一[1-2]。然而,近年來由于資源保護不當、過度捕撈、環(huán)境污染等因素的影響,海州灣資源急劇衰退,影響了海州灣海洋生態(tài)系統(tǒng)的可持續(xù)健康發(fā)展[3]。海洋微生物是海洋生態(tài)系統(tǒng)的重要組成部分,參與海洋生物地球化學(xué)循環(huán)過程,在海洋生態(tài)系統(tǒng)的功能中具有重要作用[4-5]。相關(guān)研究表明,微生物群落特征的變化會影響海洋碳、氮和磷等營養(yǎng)物的化學(xué)循環(huán)及微生物的降解功能[6-8]。沉積物在海洋生態(tài)系統(tǒng)中通常具有較高的微生物豐度和多樣性,其中的微生物是調(diào)節(jié)各種生物元素轉(zhuǎn)化和生物修復(fù)的生物地球化學(xué)過程的主要執(zhí)行者[9-10]。因此,對沉積物中微生物多樣性的研究有助于了解海洋微生物群落的分布特征,維持海洋生態(tài)資源的可持續(xù)發(fā)展。目前,對海州灣的研究主要集中在魚類多樣性、浮游動植物多樣性和海洋環(huán)境研究等方面[1-3,11-16],對海州灣沉積物中微生物多樣性的研究較少。鑒于此,本研究以海州灣不同點位的沉積物樣品為研究對象,利用 Illumina MiSeq 高通量測序技術(shù),對海州灣水域沉積物的微生物多樣性進行了分析,以期為海州灣海域生態(tài)系統(tǒng)的健康發(fā)展提供科學(xué)依據(jù)。
1" 材料與方法
1.1" 樣品采集
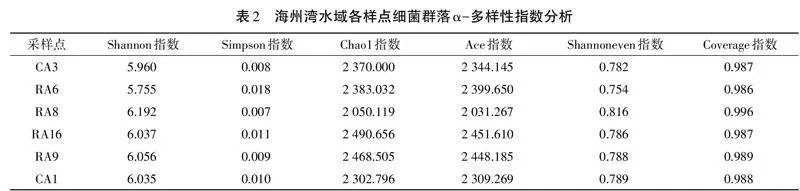
參照黃備等[17]的方法進行沉積物樣品采集。沉積物樣品采集于江蘇省連云港市海州灣,采樣點布設(shè)如表1所示。每個點位各采集3 g沉積物,3次重復(fù)。
1.2" DNA提取及高通量測序
參照張帥等[18]的方法,稱取0.5 g新鮮沉積物樣品,采用磁珠式土壤基因組DNA提取試劑盒(Omega公司,美國)提取樣品總?cè)郝浠蚪MDNA,利用Qubit 4.0(Thermo公司,美國)檢測DNA濃度后,對細菌16S rRNA基因的V3-V4區(qū)進行PCR擴增。擴增后的PCR產(chǎn)物經(jīng)2%瓊脂糖凝膠電泳進行檢測后,送至生工生物工程(上海)股份有限公司,利用Illumina MiSeq系統(tǒng)(Illumina MiSeq,USA)進行雙端測序。
1.3" 數(shù)據(jù)分析
對高通量測序得到的樣本數(shù)據(jù)進行質(zhì)控過濾以得到有效數(shù)據(jù)[19-21]。利用Usearch 11.0.667軟件將相似性>97%的序列聚類為一個OUT[22-23]。通過Mothur 1.43.0軟件計算序列的α-多樣性指數(shù)[24],其中,Chao1和Ace指數(shù)用于評估群落分布豐度,Shannon和Simpson指數(shù)用于評估群落分布多樣性,Shannoneven指數(shù)用于評估群落分布均勻度,Coverage指數(shù)能夠反映測序結(jié)果是否代表樣本的真實情況。
2" 結(jié)果與分析
2.1" "微生物α-多樣性分析
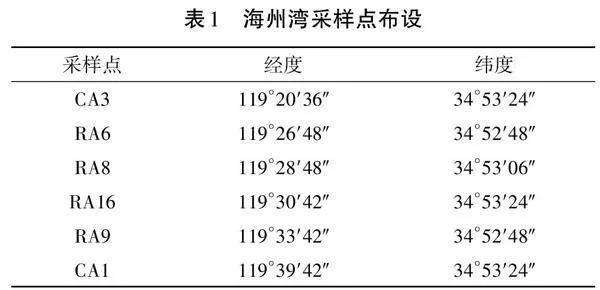
對海州灣水域各樣點采集樣品進行α-多樣性分析,多樣性指數(shù)統(tǒng)計結(jié)果見表2。Shannon指數(shù)越大,Simpson指數(shù)越小,說明微生物群落多樣性越高。RA8點位沉積物樣品的Shannon指數(shù)最大(6.192),且Simpson指數(shù)最小(0.007),表明RA8點位沉積物樣品的微生物群落多樣性最高。各采樣點沉積物樣品的Chao1指數(shù)和Ace指數(shù)不同,表明不同沉積物樣品的微生物群落豐度不同。此外,各樣品的Coverage指數(shù)均接近1(p<0.01),表明樣本測序結(jié)果能夠代表樣本微生物的真實情況。
2.2" "微生物PCoA分析
基于UniFrac距離算法,在OUT水平上,對海州灣沉積物樣品進行主坐標分析(Principal Co-ordinates Analysis,PCoA)。結(jié)果如圖1(見封三)所示,坐標軸 PCo1 能解釋45.67%的微生物群落組成差異,第二坐標軸 PCo2 能解釋25.67%的微生物群落變化。在PCo1維度上,RA8點位的沉積物樣品與其他點位的沉積物樣品距離均較遠,表明RA8點位與其他點位的沉積物樣品的微生物群落結(jié)構(gòu)差異較大。此外,CA3、RA6點位沉積物樣品的微生物群落結(jié)構(gòu)差異較小,CA1、RA9、RA16點位沉積物樣品的微生物群落結(jié)構(gòu)差異較小。
2.3" 微生物群落組成分析
通過blastn將序列與RDP、GTDB、NCBI、Silva和UNITE等數(shù)據(jù)庫進行比對,進行物種注釋,海州灣沉積物樣品中共發(fā)現(xiàn)2 851個OUTs,分屬于31門74綱116目199科331屬。門水平上(圖2,見封三),海州灣水域主要優(yōu)勢菌群為變形菌門(Proteobacteria)、擬桿菌門(Bacteroidetes)、酸桿菌門(Acidobacteria)、綠彎菌門(Chloroflexi)、放線菌門(Actinobacteria)、浮霉菌門(Planctomycetes)、 疣微菌門(Verrucomicrobia)和硬壁菌門(Firmicutes)等8類。其中,各采樣點的第一優(yōu)勢門類均為變形菌門,分別占各樣品細菌群落豐度的62.39%、53.15%、54.30%、57.97%、56.09%、58.93%。除RA8、CA3點位外,擬桿菌門在其他點位沉積物中的含量僅次于變形菌門,平均相對豐度為7.76%。
選取在屬水平上相對豐度排名前22位的細菌優(yōu)勢菌屬做熱圖分析,如圖3(見封三)所示,顏色越紅表示物種相對豐度越高,越綠表示相對豐度越低。不同點位的沉積物樣品中的優(yōu)勢菌屬存在一定差異。RA8點位沉積物樣品中相對豐度最高的菌屬為Gp10,相對豐度為3.38%,Sulfurovum(2.23%)、Thioprofundum(3.17%)和Gp23(2.28%)也是RA8點位沉積物樣品中的優(yōu)勢菌屬,平均相對豐度均超過2%;Sulfurovum是RA6、RA16點位沉積物樣品中的最優(yōu)勢菌屬,相對豐度分別為10.43%、6.02%;Thioprofundum是RA9、CA1點位沉積物樣品中的最優(yōu)勢菌屬,相對豐度分別為3.40%、3.94%;此外,CA3點位沉積物樣品中相對豐度最高的菌屬為Gp10,相對豐度為3.36%。
應(yīng)用屬水平的Venn圖分析6個點位沉積物樣品的微生物群落差異(圖4,見封三)。6個點位樣品共有的物種有191個屬,特有的物種分別為CA3點位4個,RA6點位3個,RA8點位9個,RA9點位3個,RA16點位6個,CA1點位4個。表明6個點位微生物的種類組成具有較高的相似性,各點位特有的微生物種類較少。
2.4" 共表達網(wǎng)絡(luò)分析
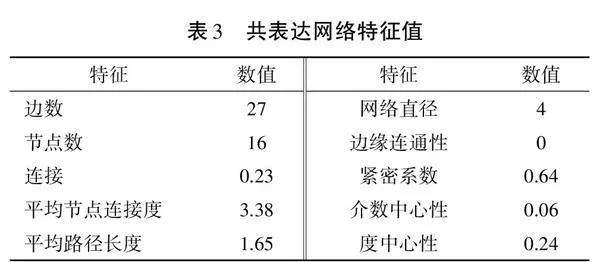
通過計算物種之間的相關(guān)性,建立物種相關(guān)性網(wǎng)絡(luò),共表達網(wǎng)絡(luò)特征值如表3所示,共表達網(wǎng)絡(luò)如圖5(見封三)所示。16 個節(jié)點間有27條表示種間強相互作用的連接(p<0.05,r>0.8),平均路徑長度是1.65,網(wǎng)絡(luò)直徑為4,緊密系數(shù)為0.64。網(wǎng)絡(luò)中的節(jié)點主要屬于變形菌門、綠彎菌門、酸桿菌門和放線菌門4個菌門,也是細菌群落的優(yōu)勢菌門。
2.5" FAPROTAX分析
通過FAPROTAX軟件分析預(yù)測微生物的功能,樣本細菌的FAPROTAX功能豐度熱圖如圖6(見封三)所示。結(jié)果表明,各采樣點沉積物樣品的功能豐度基本相似,微生物的主要功能與硫化合物呼吸(respiration of sulfur compounds)、亞硫酸鹽呼吸(sulfate respiration)、化能異養(yǎng)(chemoheterotrophy)、有氧化能異養(yǎng)(aerobic chemoheterotrophy)、發(fā)酵(fermentation)、硫酸鹽呼吸(sulfite respiration)、光合作用(chloroplasts)、光合營養(yǎng)(phototrophy)、光能自養(yǎng)(photoautotrophy)、無氧光能自養(yǎng)(anoxygenic photoautotrophy)、硝化(nitrification)、亞硝酸鹽氧化(aerobic nitrite oxidation)、尿素分解(ureolysis)、含硫化合物暗氧化(dark oxidation of sulfur compounds)、動物寄生蟲共生體(animal parasites or symbionts)、硫暗氧化(dark sulfur oxidation)、亞硫酸鹽暗氧化(dark sulfite oxidation)、硝酸鹽還原(nitrate reduction)及光能異養(yǎng)(photoheterotrophy)等有關(guān)。其中,硫化合物呼吸占5.9%~10.68%,亞硫酸鹽呼吸占5.74%~10.61%,化能異養(yǎng)占6.1%~7.38%。
3" 討論
海洋沉積物是海洋生態(tài)系統(tǒng)中主要的微生物聚集地,蘊藏著豐富的微生物資源,多種營養(yǎng)物質(zhì)轉(zhuǎn)化在此進行,是參與生物地球化學(xué)循環(huán)的重要環(huán)節(jié)[25]。海洋沉積物中微生物多樣性在維持海洋生態(tài)系統(tǒng)平衡的過程中非常重要,其相關(guān)研究已成為熱點。在門水平上,變形菌門廣泛參與海洋的生物地球化學(xué)循環(huán)過程,在有機物分解和循環(huán)中有重要作用[26]。白潔等研究發(fā)現(xiàn)黃海西北部沉積物樣品中的變形細菌門在所有樣品中均為優(yōu)勢菌門[27];劉欣等研究發(fā)現(xiàn)膠州灣海域表層沉積物樣品中絕對優(yōu)勢菌群為變形菌門[28];黃備等研究發(fā)現(xiàn)椒江口海域沉積物樣品中的最主要優(yōu)勢門為變形菌門[29];有小娟等研究發(fā)現(xiàn)變形菌門為象山港內(nèi)西滬港海域沉積物樣品中的最主要優(yōu)勢門[30]。本研究中,變形菌門在海州灣各個點位的相對豐度均最高,在細菌群落中占絕對優(yōu)勢,與國內(nèi)已報道的黃海、威海、膠州灣、椒江口及西滬港等海域沉積物樣品的最大優(yōu)勢類群相同[27-31]。除變形菌門外,海州灣沉積物樣品中還發(fā)現(xiàn)擬桿菌門、酸桿菌門、綠彎菌門、放線菌門、浮霉菌門、疣微菌門和硬壁菌門等優(yōu)勢細菌門,這些菌門在我國近海沉積物微生物群落中均有報道[32-34]。此外,海州灣不同點位沉積物樣品間的優(yōu)勢菌屬及海州灣與其他海域沉積物間的優(yōu)勢菌屬均存在較大差異,可能與環(huán)境溫度、季節(jié)、深度等環(huán)境條件及受陸地、人類活動影響不同有關(guān)[28,35]。
微生物群落間的相互作用對維持生態(tài)系統(tǒng)結(jié)構(gòu)功能穩(wěn)定具有重要意義,對微生物群落共現(xiàn)網(wǎng)絡(luò)的研究能夠進一步認識微生物之間的相互作用[36-37]。楊艷等研究發(fā)現(xiàn)長江三峽上游水域樣品微生物共現(xiàn)網(wǎng)絡(luò)中的節(jié)點主要屬于變形菌門、藍細菌門和擬桿菌門,占所有節(jié)點的 87.5%[37];王洵等研究發(fā)現(xiàn)橫山水庫表層水樣和底層水樣微生物共現(xiàn)網(wǎng)絡(luò)中的主要優(yōu)勢物種為變形菌門、放線菌門和擬桿菌門[38]。本研究發(fā)現(xiàn),海州灣沉積物樣品共現(xiàn)網(wǎng)絡(luò)中,大多數(shù)節(jié)點屬于變形菌門、綠彎菌門、酸桿菌門和放線菌門,與其他微生物相比,這些微生物在維持網(wǎng)絡(luò)結(jié)構(gòu)和功能中起重要作用[37]。如變形菌門、綠彎菌門和放線菌門廣泛參與碳、氮、硫等一系列重要元素生物地球化學(xué)循環(huán)過程,酸桿菌門在各生態(tài)系統(tǒng)中均具有特定的驅(qū)動作用及菌群穩(wěn)定性功能[39-44]。此外,相關(guān)研究表明,豐度較高的中心微生物能夠通過數(shù)量變化,豐度較低的中心微生物能夠通過介體、傳遞及聯(lián)絡(luò)等功能影響微生物群落的結(jié)構(gòu)組成和多樣性[37,45]。
海洋沉積物中的微生物驅(qū)動碳、氮和硫等許多的生物地球化學(xué)循環(huán)[46]。FAPROTAX預(yù)測的功能集中在海洋和湖泊中微生物的生物地球化學(xué)循環(huán),適用于有關(guān)S循環(huán)、N循環(huán)、H循環(huán)及C循環(huán)等的功能預(yù)測[47]。例如,王璐采用FAPROTAX軟件對黃海北部海洋牧場海水和沉積物細菌群落進行功能預(yù)測,研究發(fā)現(xiàn)黃海海水和沉積物微生物優(yōu)勢功能類群主要與碳、氮元素循環(huán)有關(guān),包括化能異養(yǎng)、有氧化能異養(yǎng)、光合作用、光合營養(yǎng)、光能自養(yǎng)、有氧光能自養(yǎng)、硝化、氨氧化、發(fā)酵、硝酸鹽還原等[48];李明月等通過FAPROTAX 數(shù)據(jù)庫預(yù)測渤海和南黃海表層沉積物中細菌功能,研究發(fā)現(xiàn)目標站位沉積物中蘊含著活躍的硫 化合物呼吸、硫酸鹽呼吸、化能異養(yǎng)、有氧化能異養(yǎng)、光合作用、硝化及好氧亞硝酸鹽氧化等硫循環(huán)、碳循環(huán)和氮循環(huán)功能[49]。在本研究中,通過FAPROTAX預(yù)測發(fā)現(xiàn)海州灣沉積物樣品中的微生物與硫化合物呼吸、亞硫酸鹽呼吸、化能異養(yǎng)、有氧化能異養(yǎng)、發(fā)酵、硫酸鹽呼吸、硝化、亞硝酸鹽氧化等硫循環(huán)、碳循環(huán)和氮循環(huán)等功能有關(guān),主要參與生物地球化學(xué)循環(huán)中的硫循環(huán)。
參考文獻:
[1]" 羅文強,趙剛,張彥彥,等.海州灣海洋牧場人工魚礁區(qū)建設(shè)前后海洋環(huán)境變化分析[J]. 海洋湖沼通報, 2021(1):33-40.
[2]" 季相星,姜毅,王普力. 2015—2018年海州灣及鄰近海域浮游植物群落結(jié)構(gòu)特征[J]. 環(huán)境監(jiān)控與預(yù)警, 2021,13(1):47-51.
[3]" 張濤,沃佳,劉逸文,等.海州灣偶見魚類的種類組成及時空分布[J].應(yīng)用生態(tài)學(xué)報,2023(3):787-795.
[4]" 王彩霞,王毅波,劉鵬遠,等.渤海三灣表層水域細菌群落結(jié)構(gòu)多樣性及其環(huán)境因子分析[J].微生物學(xué)通報,2018,45(9):1956-1971.
[5]" 羅同陽,高慶華,董聰,等.白洋淀水域夏季細菌群落結(jié)構(gòu)及與環(huán)境因子的關(guān)系[J].淡水漁業(yè),2021,51(1): 96-102.
[6]" KINGHORN A D, CHIN Y W, SWANSON S M.Discovery of natural product anticancer agents from biodiverse organisms[J]. Current Opinion in Drug Discovery amp; Development, 2009, 12(2): 189-196.
[7]" ANDREA D C, PETRA P, ESTER E, et al. The role of metal contamination in shaping microbial communities in heavily polluted marine sediments[J]. Environmental Pollution, 2020, 265: 114823-114830.
[8]" 曹洋,孫鶴銘,劉利,等.冬季衡水湖沉積物微生物群落結(jié)構(gòu)特征及影響因素[J].環(huán)境工程技術(shù)學(xué)報, 2023, 13(1):154-163.
[9]" CATANIAA V, CAPPELLOB S, DI GIORGI V D, et al. Microbial communities of polluted sub-surface marine sediments[J]. Marine Pollution Bulletin, 2018, 131: 396-406.
[10] LIU J W, LIU X S, WANG M, et al. Bacterial and archaeal communities in sediments of the North Chinese marginal seas[J]. Microbial Ecology, 2015, 70: 105-117.
[11] 施金金,孟祥實,童金榮,等.海州灣近岸海域大型浮游動物時空分布[J].水產(chǎn)養(yǎng)殖,2020,41(11):32-37.
[12] 李雪童,徐賓鐸,薛瑩,等.海州灣秋季魚類β多樣性組分分析及其與環(huán)境因子的關(guān)系[J].海洋學(xué)報,2022,44(2):46-56.
[13] 鄭江鵬,王長友,趙永剛,等.海州灣漁港經(jīng)濟區(qū)及鄰近海域沉積物主要污染物分布特征和潛在風(fēng)險[J].海洋環(huán)境科學(xué),2022,41(5):731-737.
[14] 張碩,施斌杰,謝斌,等.連云港海州灣海洋牧場浮游動物群落結(jié)構(gòu)及其與環(huán)境因子的關(guān)系[J]. 生態(tài)環(huán)境學(xué)報,2017,26(8):1410-1418.
[15] 謝斌,張碩,李莉,等.海州灣海洋牧場浮游植物群落結(jié)構(gòu)特征及其與水質(zhì)參數(shù)的關(guān)系[J].環(huán)境科學(xué)學(xué)報,2017,37(1):121-129.
[16] 楊曉改.海州灣及其鄰近海域浮游生物群落結(jié)構(gòu)及其與環(huán)境因子的關(guān)系[D]. 青島:中國海洋大學(xué),2015.
[17] 黃備,邵君波,母清林.黃海西部海域沉積物細菌群落及其對環(huán)境因子的響應(yīng)[J].生態(tài)環(huán)境學(xué)報,2019,28(7):1423-1433.
[18] 張帥,李曉康,劉禎祚,等.基于高通量測序技術(shù)分析青島市典型海灘沉積物的微生物多樣性[J].海洋環(huán)境科學(xué),2021,40(3):417-424,456.
[19] MARTIN M. Cutadapt removes adapter sequences from high-throughput sequencing reads[J]. EMBnet Journal, 2011, 17(1): 10-12.
[20] ZHANG J, KOBERT K, FLOURI T, et al. PEAR: a fast and accurate Illumina Paired-End read mergeR[J]. Bioinformatics, 2014, 30(5): 614-620.
[21] SCHMIEDER R, EDWARDS R. Quality control and preprocessing of metagenomic datasets[J]. Bioinformatics, 2011, 27(6): 863-864.
[22] EDGAR R C. UPARSE: highly accurate OTU sequences from microbial amplicon reads[J]. Nature Methods, 2013, 10(10): 996-998.
[23] EDGAR R C. SINTAX: a simple non-Bayesian taxonomy classifier for 16S and ITS sequences[J]. BioRxiv, 2016, 9: 1-20.
[24] SCHLOSS P D, WESTCOTT S L, RYABIN T, et al. Introducing mothur: open-source, platform independent, community-supported software for describing and comparing microbial communities[J]. Applied and Environmental Microbiology, 2009, 75(23): 7537-7541.
[25] 秦松,丁寧.專家論海洋生物基因資源的研究與利用[J].生物學(xué)雜志,2006,23(1):1-4.
[26] DANG H Y, LOVELL C R. Microbial surface colonization and biofilm development in marine environments[J]. Microbiology and Molecular Biology Reviews, 2016, 80(1): 91-138.
[27] 白潔,李海艷,張健,等.黃海西北部沉積物中細菌群落16S rDNA多樣性解析[J].中國環(huán)境科學(xué),2009,29(12):1277-1284.
[28] 劉欣,肖天,張文燕,等.膠州灣海域表層沉積物細菌多樣性[J].海洋科學(xué),2010,34(10):1-6,27.
[29] 黃備,邵君波,周斌,等.椒江口海域沉積物微生物群落及其對環(huán)境因子的響應(yīng)[J].中國環(huán)境監(jiān)測,2017,33(6): 87-94.
[30] 有小娟,李秋芬,張艷,等.象山港內(nèi)西滬港海域沉積環(huán)境細菌群落結(jié)構(gòu)的時空變化及其主要環(huán)境影響因子[J].應(yīng)用與環(huán)境生物學(xué)報,2013,19(5):742-749.
[31] 肖慧,張艷,張喆,等.青島、威海水域夏冬季表層沉積物細菌多樣性的初步研究[J].中國海洋大學(xué)學(xué)報(自然科學(xué)版),2009,39(4):641-646.
[32] 孫靜.黃東海細菌多樣性分析及其抗菌活性的初步評價[D].天津:天津商業(yè)大學(xué),2014.
[33] 楊少娟.廣西北部灣海洋沉積物細菌多樣性及生物活性評估[D].昆明:云南大學(xué),2022.
[34] 張健,趙陽國,李海艷,等.黃海西北近岸沉積物中細菌群落空間分布特征[J].海洋學(xué)報(中文版), 2010,32(2):118-127.
[35] 付新華,劉國寧,何健龍,等.山東省渤海海洋保護區(qū)典型海域表層海水微生物群落多樣性分析[J].海洋科學(xué), 2017,41(1):39-47.
[36] CHAFFRON S, REHRAUER H, PERNTHALER J, et al. A global network of coexisting microbes from environmental and whole-genome sequence data[J]. Genome Research, 2010, 20(7): 947-959.
[37] 楊艷,王浩,李凱航,等.長江三峽上游水域細菌群落結(jié)構(gòu)與功能預(yù)測[J].微生物學(xué)報,2022,62(4):1401-1415.
[38] 王洵,廖琴,王沛芳,等.水庫水深變化對不同浮游微生物群落及網(wǎng)絡(luò)互作關(guān)鍵種的影響[J]. 環(huán)境科學(xué),2023,44(7):3881-3891.
[39] 劉進,冀瑞卿,李冠霖,等.紅松和蒙古櫟菌根系變形菌門細菌的群落多樣性[J/OL].吉林農(nóng)業(yè)大學(xué)學(xué)報,2024-05-15.https://doi.org/10.13327/j.jjlau. 2023. 0087.
[40] 獻文東,張瀟橦,李文均.綠彎菌的研究現(xiàn)狀及展望[J].微生物學(xué)報,2020,60(9):1801-1820.
[41] 李益.森林土壤磷循環(huán)功能基因變化特征及其影響因素[D].西安:西北大學(xué),2022.
[42] 劉巖,李志恩,袁一鳴,等.基于高通量測序的象山港水域細菌多樣性的研究[J].海洋與湖沼,2022,53(3):645-653.
[43] 馬明星.不同養(yǎng)殖密度與模式下許氏平鲉腸道菌群多樣性研究[D].大連:大連海洋大學(xué), 2023.
[44] 劉彩霞,董玉紅,焦如珍.森林土壤中酸桿菌門多樣性研究進展[J].世界林業(yè)研究,2016,29(6):17-22.
[45] BANERJEE S, SCHLAEPPI K, HEIJDEN M G A. Keystone taxa as drivers of microbiome structure and functioning[J]. Nature Reviews Microbiology, 2018,16(9):567-576.
[46] FALKOWSKI P G, FENCHEL T, DELONG E F. The microbial engines that drive Earth’s biogeochemical cycles[J]. Science, 2008, 320: 1034-1039.
[47] 譚嘉怡,郝家厚,李羽志,等.施用污泥基營養(yǎng)激勵素對微型湯姆番茄品質(zhì)及土壤性質(zhì)的影響[J/OL]. 中國環(huán)境科學(xué),1-10[2024-04-16]. https://doi.org/10.19674/j.cnki.issn1000-6923.20230728.002.
[48] 王璐.海洋牧場季節(jié)性低氧區(qū)氮、磷營養(yǎng)鹽時空分布及微生物群落動態(tài)研究[D].濟南:山東大學(xué),2022.
[49] 李明月,楊雨虹,米鐵柱,等.海洋沉積物中細菌DNA和RNA水平群落差異[J].環(huán)境科學(xué),2020,41(5):2485-2495.
(責(zé)任編輯:易" 婧)
收稿日期:2023-08-03
基金項目:國家蝦蟹產(chǎn)業(yè)技術(shù)體系專項資金項目(CARS-48);江蘇省農(nóng)業(yè)公共服務(wù)補助專項(2022)。
作者簡介:顧穎(1997—),碩士,主要從事水產(chǎn)養(yǎng)殖與水生生物病害防控研究。E-mail: 1582826353@qq.com。
*為通信作者,E-mail: miaosun2007@163.com。
