黃連素下調NRAV和PI3K/AKT通路減輕RSV感染致HEp-2細胞的損傷
2024-04-23 02:12:50崔玉娟蘇東霞胡丹東
中國藥理學通報 2024年4期
崔玉娟,趙 輝,蘇東霞,張 瑩,胡丹東
(1. 西北師范大學生命科學學院,甘肅 蘭州 730070;2. 北京市延慶區疾病預防控制中心,北京 102100;3. 北京市延慶區市場監管檢驗檢測監控中心,北京 102100)
呼吸道合胞病毒(respiratory syncytial virus,RSV)屬于副粘病毒科、肺病毒屬,是一種含有包膜的反義單鏈RNA病毒,在全球范圍內具有高度的流行性,發病率呈現逐年上升的趨勢,存在較大的威脅。RSV主要感染者為兒童和老人,感染后的癥狀以支氣管炎或者其他下呼吸道疾病、發燒為主,隨著感染加劇可引起不同的組織病變,特別是引發肺炎等嚴重的呼吸系統疾病[1]。黃連素(berberine,BE)為從小檗屬植物中分離得到的生物堿,具有抗菌、抗病毒、抗炎、抗癌等多種作用,在抗病毒領域已經發現可適用于治療多種病毒感染性疾病,其機制與BE調節MAPK途徑、PI3K/AKT、AMPK/mTOR、NF-κB信號通路以及自噬密切相關[2]。不僅如此,BE還被發現可以支持宿主的免疫反應,提高機體內代謝能力,進一步強化對病毒的清除能力。由于BE毒性低、人體耐受性好,因此在抗病毒治療領域具有一定的潛力。
NRAV是近年來發現的長鏈非編碼RNA(long non-coding RNA,LncRNAs)家族成員之一,可通過抑制干擾素刺激基因(ISGs)轉錄調節抗病毒的活性,病毒的感染可以誘導宿主細胞NRAV的差異性轉錄,在免疫信號通路中發揮抗病毒作用,并介導病毒的復制,減少對宿主細胞病毒感染后的抗病毒先天免疫反應[3]。有研究表明,NRAV的過表達可促進體外RSV的復制,宿主細胞可能通過降低NRAV水平抑制RSV病毒感染[4]。PI3K/AKT信號通路與細胞的生長、分裂、存活有關,研究發現,病毒為了入侵細胞,并在細胞內進行有效的復制,通過激活該通路的方式對宿主細胞采取了抑制凋亡的策略,促使感染細胞逃避先天免疫反應而短期生存,從而使病毒在細胞凋亡前進行有效的復制,在不同類型的病毒感染過程中,均發揮關鍵作用[5]。本研究旨在探究LncRNA NRAV與PI3K/AKT信號通路在BE減輕RSV病毒感染致HEp-2細胞損傷中的作用及相關機制,為RSV感染性疾病的治療和藥物開發提供參考。
1 材料與方法
1.1 細胞、主要試劑和儀器人喉表皮樣癌細胞(HEp-2)、RSV病毒株購自美國ATCC公司。BE購自北京索萊寶科技有限公司(CAS號:2086-83-1,分子量:336.36,純度:≥98%)。NRAV過表達質粒(pc-DNA3.1-NRAV)、NRAV低表達質粒(sh-lncRNA NRAV)、NRAV陰性對照質粒(NC-lncRNA NRAV)、空白質粒(pc-DNA3.1),購自湖南普拉特澤生物科技有限公司;PI3K激活劑740Y-P(M00988)購自北京百奧菜博科技有限公司;RPMI 1640培養基、DMEM培養基、二甲基亞砜(DMSO)(200-664-3)、MitoSOX工作液(1821370-28-8),購買自西格瑪奧德里奇(上海)貿易有限公司;CCK-8試劑(HY-K0301)購自德國默克公司;一抗抗體PI3K、AKT、NLRP3、ASC、caspase-1、PINK1、Parkin、P62、Beclin1、LC3 I、LC3 II、BNIP3購自美國Cell Signaling Technology公司;IL-1β(SEKH-0002)、IL-6(SEKH-0013)、IL-8(SEKH-0016)、TNF-α(SEKH-0047)ELISA檢測試劑盒購自北京索萊寶科技有限公司;PBA緩沖液、JC-1試劑盒(C2005)、ATP試劑盒(S0026)購自上海碧云天生物技術有限公司。流式細胞分析儀(CytoFLEX型)購自美國美國貝克曼庫爾特公司;酶標儀購自賽默飛世爾科技(中國)有限公司;全自動蛋白印跡系統(Qblot型)購自上海曼博生物醫藥科技有限公司;激光共聚焦顯微鏡(OLS5100型)購自日本奧林巴斯公司。
1.2 方法
1.2.1細胞培養、分組及處理 HEp-2細胞用含有10%胎牛血清的DMEM培養基在37 ℃、5% CO2培養箱中培養,取對數生長期HEp-2細胞按照1×105/孔接種于6孔板中,細胞生長度達80%~90%時備用。取適量BE在DMSO溶液中充分溶解,配置濃度分別為5、10、15 μmol·L-1的BE溶液,RSV處理劑量為200 pfu/孔。將HEp-2細胞隨機分為:pcDNA3.1-NRAV組(細胞轉染pcDNA3.1-NRAV,RSV處理)、sh-lncRNA NRAV組(細胞轉染sh-lncRNA NRAV,RSV處理)、NC-lncRNA NRAV組(細胞轉染NC-lncRNA NRAV,RSV處理)、Control組(不用RSV和BE處理)、RSV組(RSV處理)、RSV+5 μmol·L-1-BE組(RSV和5 μmol·L-1的BE處理)、RSV+10 μmol·L-1-BE組(RSV和10 μmol·L-1的BE處理)、RSV+15 μmol·L-1-BE組/RSV+BE組(RSV和15 μmol·L-1的BE處理),RSV+BE+740Y-P組(RSV、15 μmol·L-1的BE和50 μg·L-1的740Y-P處理)、RSV+BE+pcDNA3.1組(細胞轉染pcDNA3.1,且RSV、15 μmol·L-1的BE處理)、RSV+BE+pcDNA3.1-NRAV組(細胞轉染pcDNA3.1-NRAV,且RSV、15 μmol·L-1的BE處理)對照組和感染組則加入DMSO 20 μL,BE處理組加入相應濃度的BE溶液20 μL。各組細胞在37 ℃、5% CO2的細胞培養箱中培養24 h后,收集細胞檢測相應指標。
1.2.2細胞轉染 細胞在60 mm培養皿中以3.0×105個/皿的密度培養24 h。然后,使用Lipofectamine 2000轉染試劑將pcDNA3.1-NRAV、sh-lncRNA NRAV、NC-lncRNA NRAV、pcDNA3.1質粒3 μg轉染至HEp-2細胞中,在37 ℃、5%CO2的培養箱中培養48 h。
1.2.3qRT-PCR檢測mRNA表達水平 收集培養24 h后的各組HEp-2細胞,用TRIzol法提取總RNA,反轉錄后進行PCR,以GAPDH為內參,采用2-ΔΔCT法定量NRAV、RSV-F、NS2的相對表達水平。
1.2.4Western blot檢測HEp-2細胞相關蛋白表達 收集培養24 h后的各組HEp-2細胞,用RIPA裂解液提取總蛋白,用BCA法測濃度,十二烷基硫酸鈉-聚丙烯酰胺凝膠電泳(SDS-PAGE),并轉膜,用5%脫脂奶粉封閉,加入相應一抗(1 ∶1 000),4 ℃過夜。加入辣根過氧化物酶標記的二抗(1 ∶5 000),37 ℃孵育1 h。ECL顯色、暗室曝光。以GAPDH為內參,利用Quantity One軟件分析各條帶灰度值。目的蛋白表達水平=目的蛋白灰度值/GAPDH灰度值×100%。

Tab1 Primer sequences
1.2.5CCK-8實驗檢測HEp-2細胞存活率 收集培養24 h后的各組HEp-2細胞,加入100 μL CCK-8試劑,37 ℃孵育1 h,檢測450 nm處的吸光度值,計算細胞存活率。細胞存活率=實驗組吸光度值/對照組吸光度值×100%。
1.2.6流式細胞術檢測HEp-2細胞凋亡率和線粒體膜電位 HEp-2細胞培養48 h后,收集細胞,用冰冷的PBS洗滌兩次,并在70%冰冷的乙醇中在4 ℃下固定過夜。在PBS中再水化15 min后,將細胞在黑暗中用PI溶液染色30 min,通過流式細胞術分析細胞凋亡率。HEp-2細胞培養24 h后,2 000 r·min-1離心5 min,棄上清,加入500 μL的JC-1工作液,37 ℃下孵育20 min。2 000 r·min-1離心5 min,棄上清,緩沖液洗滌細胞,并重懸,用流式細胞儀分析各組HEp-2細胞線粒體膜電位。線粒體膜電位較高時,JC-1聚集在線粒體基質中,形成聚合物,呈紅色熒光。線粒體膜電位較低時,JC-1為單體,不能聚集在線粒體基質中,呈綠色熒光。線粒體膜電位用紅色熒光細胞的比例表示。
1.2.7檢測HEp-2細胞中ATP水平 收集培養24 h后的各組HEp-2細胞的培養液上清,加入ATP檢測溶液,用多功能酶標儀測量發光強度。隨后根據標準曲線計算ATP含量(nmol·mg-1蛋白)。
1.2.8MitoSOX染色檢測線粒體活性氧(mtROS)水平 HEp-2細胞培養24 h后,加入終濃度5 μmol·L-1的MitoSOX工作液,37 ℃避光孵育20 min,PBS洗滌,加入DAPI工作液,室溫避光孵育15 min,PBS洗滌,激光共聚焦顯微鏡下觀察。紅色為mtROS,藍色為細胞核。
1.2.9ELISA檢測HEp-2細胞炎性因子分泌水平 收集培養24 h后的各組HEp-2細胞的培養液上清,按照ELISA試劑盒說明書檢測IL-1β、IL-6、IL-8、TNF-α水平。
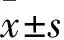
2 結果
2.1 LncRNA NRAV對RSV感染HEp-2細胞的影響qRT-PCR檢測LncRNA NRAV的轉染效率,pcDNA3.1-NRAV組中NRAV表達水平明顯高于sh-lncRNA NRAV組與NC-lncRNA NRAV組(P<0.05),而sh-lncRNA NRAV組最低(P<0.05)(Fig1A)。過表達NRAV使RSV活性基因RSV-F、NS2明顯升高(P<0.05),敲低NRAV后活性降低(Fig1B)。pcDNA3.1-NRAV組細胞活性最低,凋亡率最高(P<0.05),sh-lncRNA NRAV組結果相反(Fig1C-E)。過表達NRAV可以抑制感染RSV的HEp-2細胞的活性,促進病毒復制,并促進感染細胞凋亡。
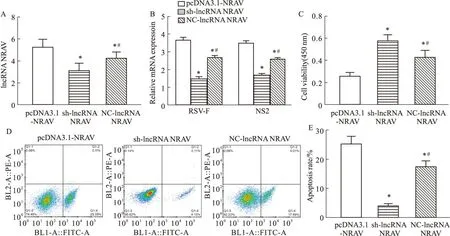
Fig1 Effect of LncRNA NRAV on RSV infection in HEp-2 cells
2.2 BE對RSV感染HEp-2細胞中NRAV和PI3K/AKT信號通路的影響RSV感染后明顯降低了HEp-2細胞中的NRAV表達水平,BE處理后進一步降低了NRAV的表達水平,且呈劑量依賴性(P<0.05)(Fig2A)。信號通路檢測結果顯示,RSV感染HEp-2細胞后,細胞中PI3K、AKT蛋白的表達水平成功上調,PI3K/AKT信號通路被激活,Control組HEp-2細胞中PI3K、AKT的表達水平最低(P<0.05)。與RSV組相比,BE組干預的3組細胞中PI3K、AKT蛋白表達水平均下降,且隨濃度的增加其表達量呈劑量依耐性降低(P<0.05)(Fig2B、C)。
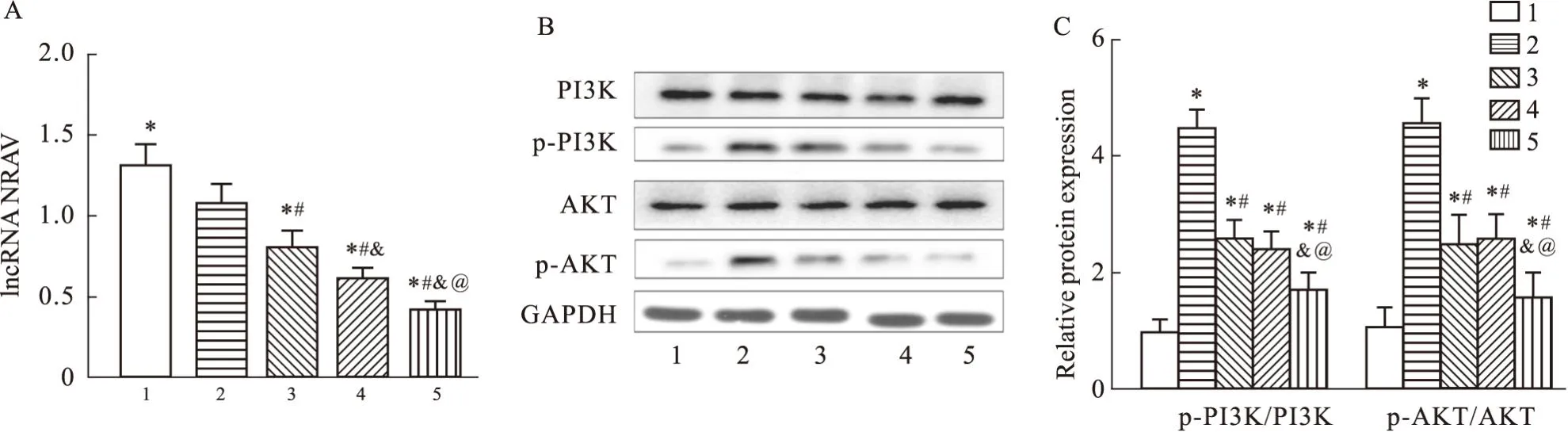
Fig2 Effect of BE on NRAV and PI3K/AKT signaling pathways in RSV infected HEp-2 cells
2.3 過表達NRAV、激活PI3K/AKT通路逆轉BE引起的通路抑制RSV感染HEp-2細胞后,BE抑制了由病毒感染引起的PI3K/AKT信號通路激活(P<0.05)。PI3K激活劑740Y-P處理后,逆轉了BE對RSV感染HEp-2細胞中PI3K/AKT信號通路的抑制作用,PI3K、AKT的表達水平明顯升高(P<0.05)。過表達NRAV同樣可以逆轉BE對RSV感染HEp-2細胞中PI3K/AKT信號通路的抑制作用,差異具有統計學意義(P<0.05),與740Y-P處理組相比無統計學意義(P>0.05)(Fig3)。
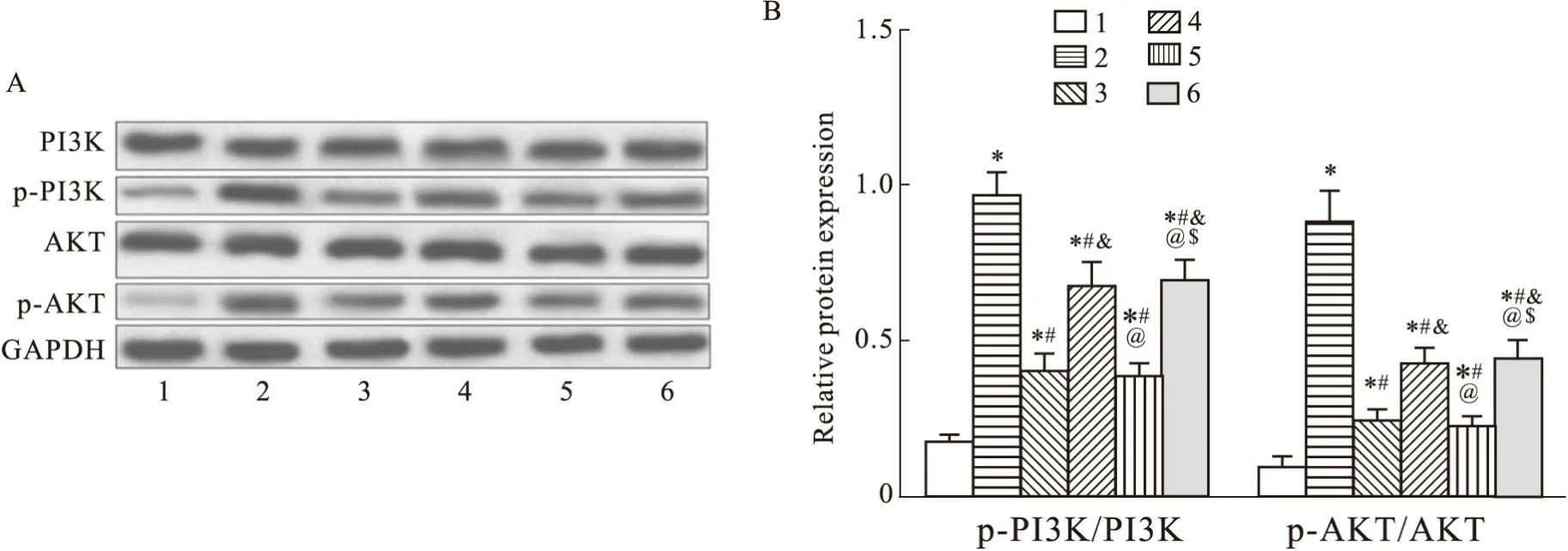
Fig3 Overexpression of NRAV and activation of PI3K/AKT pathway reverse pathway inhibition caused by BE
2.4 過表達NRAV、激活PI3K/AKT通路加重線粒體損傷RSV感染降低了HEp-2細胞的ATP、線粒體膜電位(P<0.05),并明顯增加了HEp-2細胞的mtROS水平(P<0.05)。BE處理則增加了RSV感染HEp-2細胞的ATP、線粒體膜電位(P<0.05),并降低了HEp-2細胞的mtROS水平(P<0.05)。PI3K激活劑740Y-P處理、過表達NRAV均可逆轉BE對RSV感染HEp-2細胞ATP、線粒體膜電位、mtROS水平的影響(P<0.05),加重了線粒體損傷(Fig4)。
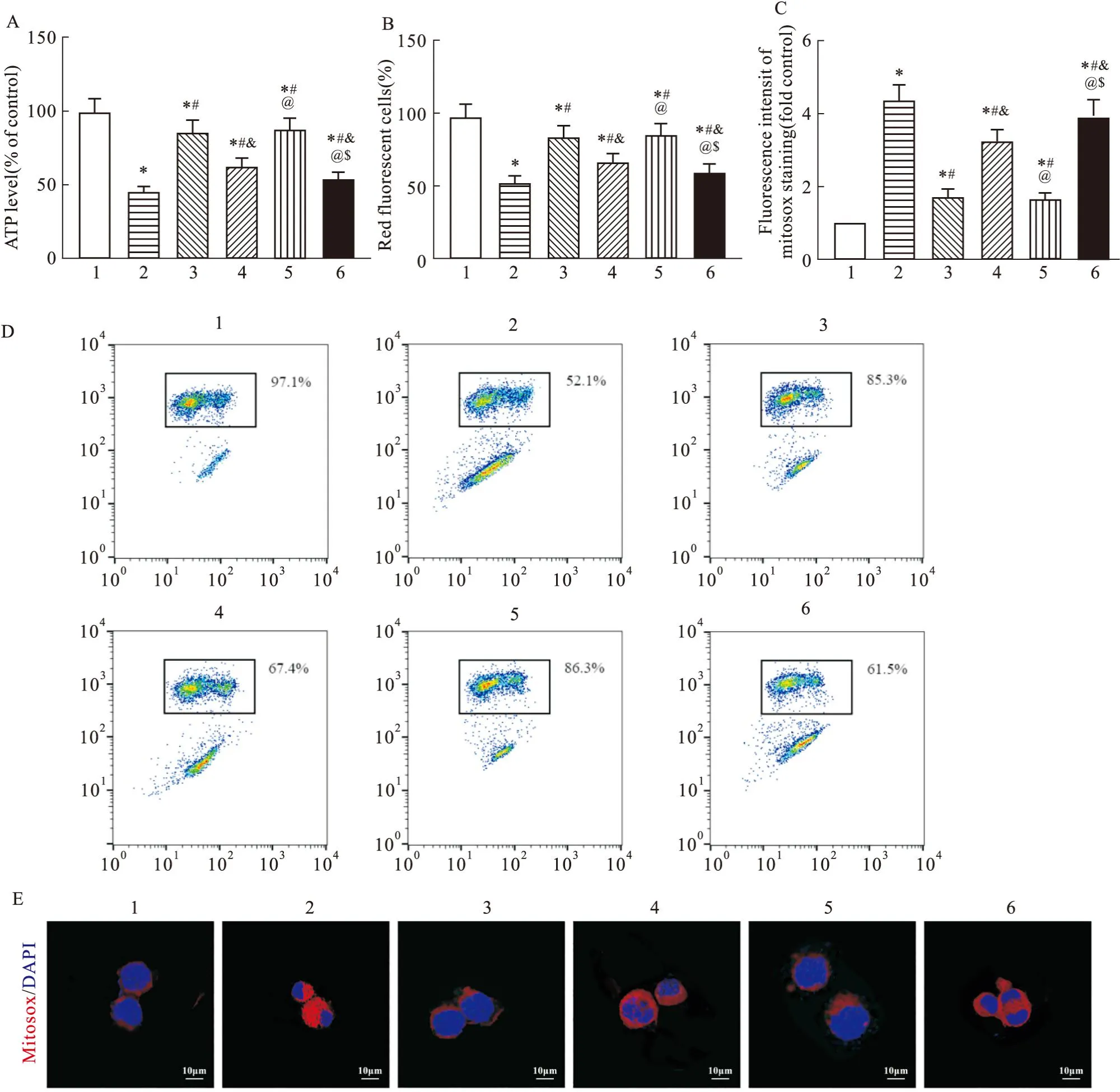
Fig4 Overexpression of NRAV and activation of PI3K/AKT pathway exacerbate mitochondrial damage
2.5 過表達NRAV、激活PI3K/AKT通路降低線粒體自噬水平BE干預明顯增加了RSV感染HEp-2細胞線粒體自噬標志蛋白PINK1、Parkin、Beclin1、LC3Ⅱ/LC3Ⅰ、BNIP3的表達水平(P<0.05),降低了p62的表達水平(P<0.05),誘導PINK1/Parkin、BNIP3途徑共同介導了線粒體自噬,減輕細胞損傷。PI3K激活劑740Y-P處理、過表達NRAV則逆轉了BE對自噬標志蛋白的影響(Fig5)。
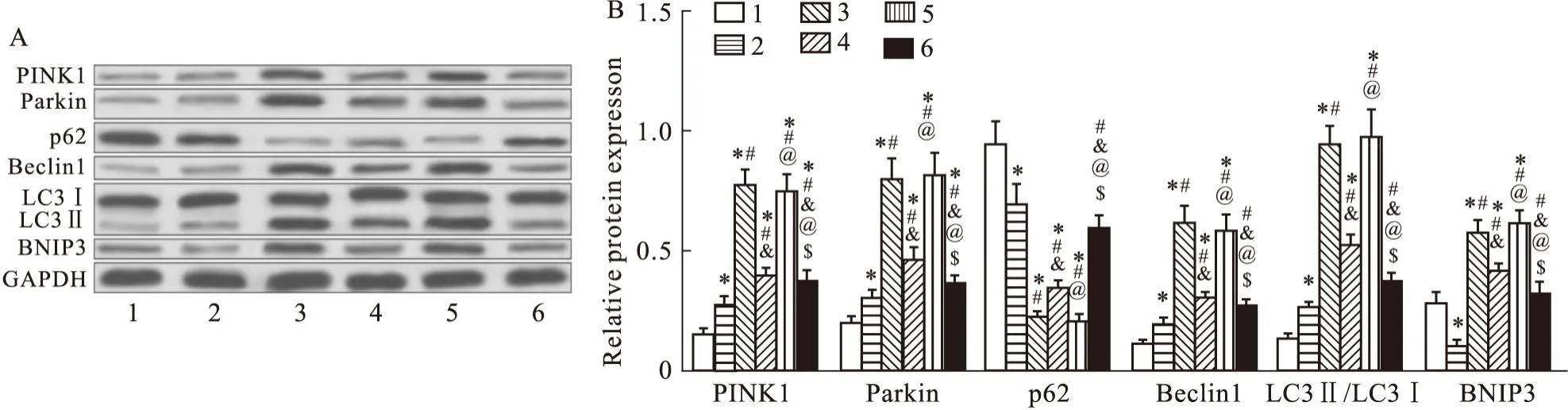
Fig5 Overexpression of NRAV and activation of PI3K/AKT pathway reduce mitochondrial autophagy levels
2.6 過表達NRAV、激活PI3K/AKT通路增加凋亡率、加重炎癥反應RSV組凋亡率為27.22%,與Control組相比細胞損傷嚴重(P<0.05)。BE干預后,細胞活性升高,細胞凋亡率降低至9.75%(P<0.05)。通路激活劑740Y-P和過表達pcDNA3.1-NRAV質粒干預后,細胞活性降低,而細胞凋亡率又分別上升至18.58%和22.98%,細胞損傷嚴重(P<0.05)(Fig6A-C)。BE干預后明顯抑制了炎性小體的活性(P<0.05),而通路激活劑740Y-P和過表達pcDNA3.1-NRAV質粒的干預,逆轉了BE的作用效果,炎性小體被再次活化,加重了細胞的炎癥反應(Fig6D-F)。
3 討論
RSV是在人類中廣泛流行的病原體,目前尚缺乏特效的藥物,嚴重威脅人類健康,對RSV抗病毒分子機制的研究將為未來抗病毒藥物的研發提供新靶點和新方向。BE作為一種傳統的中藥提取物,在多種病毒感染性疾病中表現出顯著的抗病毒功效。研究表明,BE可以有效地靶向抑制宿主AP1和阻斷病毒蛋白,發揮對HPV的抗病毒能力[6],并通過抑制逆轉錄酶的活性抑制HIV的復制[7],通過誘導線粒體自噬,降低線粒體ROS的產生,進而抑制IAV引發的炎癥反應[8]。眾所周知,病毒感染過程及許多抗病毒藥物的藥理機制還涉及LncRNAs的調控,NRAV在RSV、IAV感染期間明顯下調,過表達NRAV可促進RSV在A549和BEAS-2B細胞中的復制,促進IAV的復制和毒力,NRAV的下調可能是宿主抗病毒先天免疫反應的一部分[4,9]。而我們過往的研究還發現,BE可以通過NRAV、RAB5C靶向競爭結合miR-299-3p,抑制JNK/p38 MAPK信號通路和PI3K/AKT信號通路提高HSV-1感染HEp-2細胞的存活率,并降低凋亡率[10]。基于上述結果,本研究從過表達NRAV和激活PI3K/AKT信號通路入手,探究BE對RSV感染HEp-2細胞的影響。
在本研究中,過表達NRAV后,感染RSV的HEp-2細胞活性降低,凋亡率增加,促進了病毒復制,而敲低NRAV則出現了完全相反的結果。NRAV的過表達,能夠使RSV大量增殖,破壞宿主的正常細胞結構,促使細胞的加速凋亡,從而造成一系列的炎癥反應與病理損傷產生。RSV感染細胞后,宿主激活先天免疫反應使NRAV的表達水平明顯降低,BE干預處理可進一步降低RSV感染的HEp-2細胞NRAV表達水平,結果表明NRAV可能參與BE的抗病毒過程。
研究發現,在許多病毒感染期間PI3K/AKT信號通路會被病毒激活,并進一步促進病毒入侵和復制[11]。在病毒感染早期,進入宿主細胞后病毒為了在宿主細胞內有效的增殖,采取了激活PI3K/AKT信號通路促進感染細胞生存的策略,使病毒在細胞死亡之前能夠大量增殖。細胞的抗病毒先天免疫反應通過抑制PI3K/AKT信號通路促進感染細胞凋亡[5],抑制病毒的復制。研究發現NRAV的表達水平隨BE濃度的增加而降低,PI3K/AKT信號通路相關蛋白的表達水平隨BE濃度的增加而降低,推測BE通過上述分子機制發揮對RSV的抗病毒作用。進一步用PI3K激活劑740Y-P處理后,在BE的作用下通路蛋白的表達水平仍然明顯升高,逆轉了BE對病毒感染HEp-2細胞中PI3K/AKT信號通路的抑制作用,而過表達NRAV也得到了同樣的抑制結果。通過上述實驗可知,在過表達NRAV以及激活PI3K/AKT信號通路后,BE對PI3K/AKT信號通路的抑制作用被逆轉,BE則可能通過下調NRAV并抑制PI3K/AKT信號通路抑制RSV的感染能力。
病毒感染致細胞損傷與線粒體受損有關。受損線粒體的膜電位降低進一步導致ATP合成受阻,且釋放ROS,導致ROS大量積累,進而誘導細胞與組織損傷[12]。線粒體損傷受線粒體自噬的調控,線粒體自噬是一種專一降解損傷或老化線粒體的選擇性自噬,其對維持線粒體數量及正常功能、保持線粒體ROS平衡非常重要[13],并且在抵御病原體感染中發揮重要作用。PINK1/Parkin通路和BNIP3通路的激活均可介導自噬小體對損傷線粒體的識別和包裹,進而誘導線粒體自噬[14-15]。在本研究中,RSV感染HEp-2細胞后,線粒體內膜兩側外正、內負的線粒體膜電位下降,線粒體膜電位的異常影響氧化磷酸化和線粒體ATP合成,同時釋放了大量的ROS,導致HEp-2細胞的線粒體嚴重受損。PINK1在線粒體外膜積累,進一步招募Parkin并泛素化受體蛋白和適配器蛋白,PINK1/Parkin和介導的線粒體自噬被激活。在BE干預后,線粒體功能得到了恢復,線粒體膜電位升高、ATP水平升高、ROS水平降低,PINK1/Parkin和BNIP3介導的線粒體自噬均被激活,BE改善了線粒體功能,維持了線粒體的穩定。而740Y-P處理和過表達NRAV后,在BE的作用下線粒體膜電位降低、ATP合成減少、細胞中ROS水平升高,線粒體功能仍然受損,線粒體自噬蛋白水平降低,自噬通路被抑制,逆轉了BE對RSV感染HEp-2細胞線粒體功能的改善作用。過表達NRAV對線粒體功能損傷的程度要大于740Y-P激活PI3K/AKT信號通路后對線粒體的損傷程度。有文獻顯示[16],抑制PI3K/AKT信號通路可通過促進線粒體自噬,促進線粒體功能恢復,進而減輕細胞損傷。因此,結合本次研究的結果可知,BE可能通過下調NRAV并抑制PI3K/AKT信號通路提高線粒體膜電位、提高ATP水平、降低mtROS水平,激活線粒體自噬,改善RSV導致的線粒體功能障礙和細胞損傷。
為進一步研究NRAV與PI3K/AKT信號通路在BE抗RSV病毒過程中的作用,本研究進行了細胞增殖、凋亡水平、炎癥反應檢測。結果發現,BE處理可增加RSV感染HEp-2細胞的存活率并降低凋亡率。740Y-P處理或過表達NRAV可逆轉BE對RSV感染的HEp-2細胞增殖、凋亡的影響。表明NRAV與PI3K/AKT信號通路可促進RSV感染導致的HEp-2細胞損傷。BE減輕RSV致HEp-2細胞損傷的機制可能與BE下調NRAV和PI3K/AKT信號通路通路有關。NLRP3炎性小體能被多種類型的病原體或危險信號激活,在機體免疫反應和疾病發生過程中具有重要作用,是機體介導免疫炎癥的關鍵調控因子。病毒感染后,線粒體損傷會釋放大量ROS,進而使NLRP3炎性小體過度活化,促進IL-1β、IL-6、IL-8、TNF-α等炎癥因子成熟釋放,誘導細胞高炎癥狀態,導致細胞損傷,進而發生細胞凋亡[17]。大量的研究證明,NLRP3炎性小體的過度活化是病毒感染導致細胞損傷的重要原因之一[18]。研究顯示,RSV感染可明顯提高HEp-2細胞內 NLRP3、ASC、caspase-l蛋白表達及IL-1β、L-6、IL-8、TNF-α分泌水平,導致 NLRP3 炎性小體過度活化,使感染細胞的存活率降低,這與既往病毒感染的結果相似。BE干預可降低RSV感染HEp-2細胞內NLRP3、ASC、caspase-l蛋白表達及炎性因子分泌水平,抑制NLRP3 炎性小體活化。740Y-P處理或過表達NRAV可再次激活炎性小體、促進炎性因子分泌,逆轉了BE抑制RSV感染的HEp-2細胞炎癥反應的作用。結果提示BE可能通過下調NRAV并抑制PI3K/AKT信號通路抑制NLRP3炎性小體活化對RSV感染HEp-2細胞起保護作用。
綜上所述,過表達NRAV和激活PI3K/AKT信號通路可以逆轉BE改善線粒體功能、激活線粒體自噬、增加細胞活性、降低細胞凋亡率、減輕炎癥反應等作用。BE減輕RSV感染所致HEp-2細胞損傷,其機制可能與BE下調NRAV和PI3K/AKT信號通路,進而激活線粒體自噬、降低線粒體損傷與炎癥反應有關。然而,BE的抗病毒作用可能不僅僅是通過上述機制進行的,可能是由其他細胞機制共同作用的結果,針對這一點將在后續的研究中進行更多的驗證,以全面了解BE對RSV的抗病毒分子機制。
猜你喜歡
美與時代·美術學刊(2022年3期)2022-04-27 01:18:15
鴨綠江(2021年35期)2021-04-19 12:24:18
考試與評價·高一版(2020年6期)2020-11-02 02:45:24
火花(2019年12期)2019-12-26 01:00:28
人大建設(2019年12期)2019-05-21 02:55:32
中國生殖健康(2019年3期)2019-02-01 06:12:26
鑿巖機械氣動工具(2016年3期)2016-03-01 04:00:25
學苑創造·A版(2015年11期)2016-01-14 09:03:27
海軍航空大學學報(2015年3期)2015-11-11 17:20:00
中國火炬(2010年8期)2010-07-25 11:34:30
