中性粒細胞胞外誘捕網在膿毒癥病理生理及臨床應用中的研究進展
2024-03-26 06:53:08王蕓飛李燕
安徽醫藥 2024年4期
關鍵詞:研究
王蕓飛,李燕
作者單位:山西醫科大學第二醫院急診醫學科,山西 太原030001
膿毒癥是由于宿主對感染的反應失調而導致的危及生命的器官功能障礙綜合征[1],世界各地有無數病人因此而過早死亡或面臨長期殘疾。由于是一種潛在感染引起的綜合征,標準的全球疾病、傷害和風險因素負擔研究(GBD)只能用來估計其風險與負擔:2020年Lancet發布的膿毒癥相關全球疾病負擔報告中顯示,2017年全世界估計有48.9萬例敗血癥病例和11.0萬例敗血癥相關死亡,占全球所有死亡人數的19.7%[2]。正因為這較高的發生率及可怕的病死率,2017年5月世界衛生組織(WHO)認定膿毒癥為“全球醫療優先關注的疾病”。
自“膿毒癥”這一概念提出以來,我們與之已經斗爭了近30多年,但目前仍沒有一種治療方法可以持續、長期地挽救膿毒癥病人的生命,其根本原因可能是膿毒癥起源的異質性及與之伴隨的病理生理機制的未知性。目前普遍認為膿毒癥的病理學特征是炎癥失衡和免疫抑制并存[3]。中性粒細胞是感染發生后的第一道防線,在早期到達感染或炎癥部位,并通過吞噬、氧化爆發、脫顆粒和產生中性粒細胞胞外誘捕網(NETs)等功能發揮抗感染、促炎等作用。骨髓粒細胞生成以50~100億次/日來維持其周轉。自從2004年Brinkman等[4]發現中性粒細胞通過擠壓排出DNA-蛋白質結構形成NETs來捕獲和殺死組織中的細菌以來,越來越多的研究探索NETs在膿毒癥發生發展中的作用和機制,并由此提出許多臨床治療新方向。
1 產生機制
目前的研究發現,NETs的產生是感染過程中,中性粒細胞在外化其細胞內容物(即飾有抗菌蛋白和絲氨酸蛋白酶的DNA框架)的同時發生程序性細胞死亡,稱為NETosis。早在人們對NETs形成機制有所認識之前,Urban、Backman[5]就通過蛋白質組學分析基本確定了其框架,即:以組蛋白(H)、DNA為主要成分(約70%),內含顆粒酶和多肽,如中性粒細胞彈性蛋白酶(NE)、抗菌肽(LL-37)、髓過氧化物酶(MPO)、組織蛋白酶G、白細胞蛋白酶3(PR-3)、乳鐵蛋白、明膠酶、溶菌酶C以及鈣衛蛋白等。一般來說,所有形式的NETosis都需要細胞內膜的重組,從而允許胞質內的各種酶和核內的DNA及組蛋白結合。綜合最新的研究結果,NETs的產生可以分為自殺式的NETosis和保留生命的NETosis,也有實驗發現這兩者在刺激后有時間上的先后差別[6],同時DNA組分可有線粒體或核DNA之分。
經典的(或自殺式的)NETosis,由實驗時的典型代表佛波醇-12-肉豆蔻酸酯-13醋酸酯(PMA)或體內的病原相關模式分子(PAMPs)和損傷相關分子模式(DAMPs)等激活,通過模式識別受體(如TOLL樣受體TLR)、補體系統受體(CR)、Fc受體(FcγRIIA和FcγRIIIb)或細胞因子受體等膜受體識別,激活蛋白激酶C(PKC)及其下游的Raf-MEK-ERK通路(絲裂原活化蛋白激酶MAPK上游信號通路)和NADPH氧化酶2(NOX2)-活性氧自由基(ROS)通路,胞質中蓄積的ROS、Ca2+作為第二或第三信使激活肌動蛋白及制孔蛋白(如gasdermin D,GSDMD),協助胞質或噬天青顆粒中的NE、肽基精氨酸脫亞胺酶4(PAD4)、MPO或去乙酰化酶(HDAC)等入核,促進核染色質解縮,激活DNA酶并啟動NETosis[6-12]。局部染色質重塑、組蛋白的表觀遺傳修飾是NETosis的先決條件[13],最終以中性粒細胞死亡而收場。
近年來,研究發現了“保留生命的NETosis”,即一部分中性粒細胞(約10%)被刺激(如一定濃度的脂多糖)后,通過PRR傳導信息,在被激活后約10 min擠出裝載核DNA的囊泡而不破壞質膜,這一過程并不產生大量的ROS,最終產生NETs和保留了吞噬、遷移和趨化等能力的無核粒細胞[12]。以上反應中,解聚的核物質是如何在保持細胞整體形態的情況下裝載、運輸、釋放到保外的,其機制仍有待進一步研究。以上差別可能是刺激強度的、胞內鈣蓄積的濃度及氧化還原代謝狀態有差異,才能在產生NETs、級聯激活免疫反應的同時,保留中性粒細胞的吞噬和趨化能力,最大程度避免了強刺激下迅速的中性粒細胞解體[6],延長了固有免疫的續航能力,為膿毒癥免疫抑制研究提供了新思路。
隨著研究的逐步深入,人們發現線粒體可能在ROS積累和鈣蓄積環節有著不可替代的作用,甚至線粒體DNA可能是NETs中DNA骨架之一[7-8]。對斑馬魚的研究證實了DNA聚合酶γ、線粒體的DNA聚合酶(Polg)對中性粒細胞運動、線粒體ROS富集的影響[8]。同時,膿毒癥發生時,小腦和海馬中線粒體的呼吸鏈復合體連接發生異常,具體表現為復合體Ⅰ連接底物氧化、ADP刺激呼吸和呼吸受體控制率減少[7]。有研究發現中性粒細胞暴露于細菌后的5 min內發生的早期抗菌事件中,NETs的形成依賴于線粒體呼吸鏈復合體Ⅲ的功能[7-8]。此過程中糖酵解更加活躍,ATP/ADP或乳酸、磷酸烯醇式丙酮酸等糖酵解產物是否可以如信號分子一樣發揮關鍵的激發甚至啟動作用?線粒體在中性粒細胞的抗菌反應中發揮的作用具有廣闊的研究潛力,這也可能將是未來研究突破的重要方向。
隨著對NETosis研究的深入,人們發現刺激的異源性可能是產生機制多樣化的根源:實驗狀態下由PMA刺激誘導,NETosis表現出相對固定的Raf-MEK-ERK和NOX2依賴的特性,而在復雜的內環境中不同種類及濃度的PAMPs和DAMPs帶來豐富的可能性,研究者們嘗試從膿毒癥來源的異質性探究NETosis的節點,見表1。
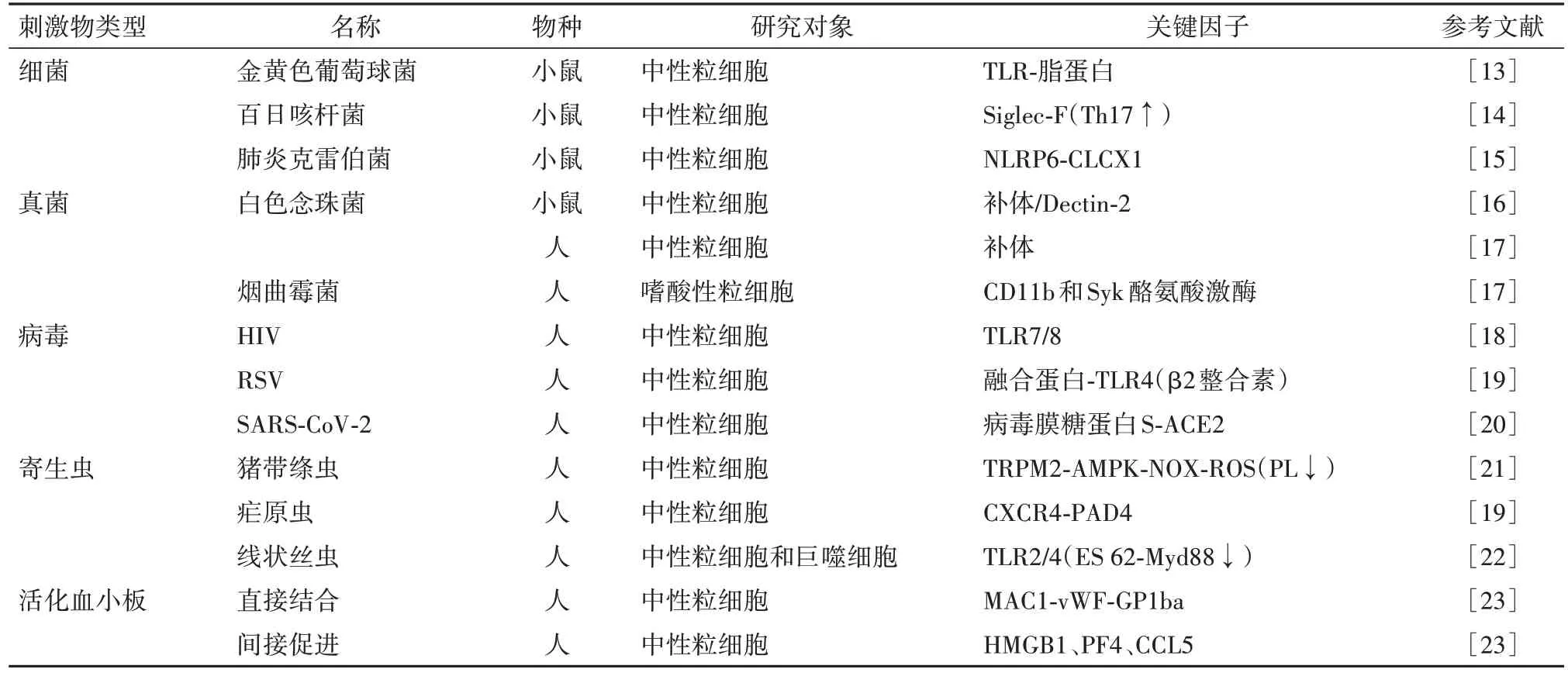
表1 膿毒癥來源異質性形成胞外誘捕網節點的研究
1.1 細菌與NETs 金黃色葡萄球菌是一種有較高發病率(北美人口中發病率約4.3~38.2例/100 000人年[14])和病死率(10%~30%[14])的病原體,一般認為其通過脂蛋白與TLR結合促進NETs形成。而最近一項研究發現,耐甲氧西林金黃色葡萄球菌通過TLR-2/1誘導的NETosis并不依賴脂蛋白,此過程也不需要NOX2、ROS參與[15]。百日咳桿菌中Ig樣凝集素(Siglec)-F可能是NETs生成的關鍵途徑,并可以被IL-17增強。一項以肺炎克雷伯菌為研究對象的實驗證明,革蘭陰性菌以核苷酸結合的寡聚化結構域樣受體(NLRP)6為上游驅動因子激活NETosis,而外源性重組趨化因子配體(CXCL1)可挽救NLRP6-/-的小鼠中性粒細胞的趨化、聚集、顆粒釋放及NETs形成的能力[17]。
1.2 真菌與NETs 免疫受損病人機會性、侵襲性的真菌感染的發病率和病死率大大增加,是膿毒癥的重要來源。既往普遍認為,真菌誘導的NETosis主要是與PMA刺激相似的ROS依賴的PKC途徑[15]。越來越多的文獻指出NETs在機會性真菌感染中的激活途徑不是單一的,而是受病原體、感染部位和宿主免疫狀態共同影響。如實驗鼠中由CR3負責與調理后的白色念珠菌結合,由PRR(如Dectin-2)識別未調理的白色念珠菌。人類中性粒細胞則使用CR3識別未調理的白色念珠菌和煙曲霉菌[17]。關于煙曲霉菌的研究發現,嗜酸性粒細胞外陷阱的釋放是通過CD11b和Syk酪氨酸激酶的溶解性非氧化機制實現的,依賴于Src家族、Akt、鈣和p38-MAPK信號通路[18]。關于真菌刺激NETs產生的研究相對局限,有待進一步的研究。
1.3 病毒與NETs 以新型冠狀病毒為典型代表的病毒感染因其多變的臨床癥狀和嚴重程度給全球醫療系統帶來巨大壓力。一方面,目前的研究認為病毒普遍通過中性粒細胞表面PRRs誘導NETs形成,例如:①人類免疫缺陷病毒(HIV)通過TLR-7/8刺激中性粒細胞產生NETs;②呼吸道合胞病毒(RSV)在β2整合素參與下通過融合蛋白與TLR4結合誘導NETosis。另一方面,病毒也可以不通過PRRs依賴的途徑誘導NETs形成,如通過炎性趨化因子和細胞因子如干擾素和白細胞介素(IL)-8或激活的血小板[19]參與NETs形成。在針對新冠肺炎的研究中發現,SARS-CoV-2引發的NETosis啟動于病毒膜糖蛋白S與血管緊張素轉換酶2(ACE2)[20]之間的分子相互作用,由絲氨酸蛋白酶(TMPRSS2)及PAD4參與下一步的代謝反應。該研究還發現ACE2與SARS-CoV-2導致的免疫抑制反應有關[20]。
1.4 寄生蟲與NETs 隨著社會經濟文化水平的提高,寄生生物所致的疾病對于社會生活的影響逐漸減小,目前關于寄生因素與NETs形成及其機制的報道相對較少。一部分寄生蟲具有出色的免疫抑制能力,可以躲避免疫細胞的抓捕,在體內潛伏感染數年之久。這能否為膿毒癥后免疫功能紊亂特別是免疫抑制狀態的研究提供新方向?蠕蟲有獨特的在人體中建立慢性、無癥狀感染的能力,最近一項研究嘗試探討以豬帶絳蟲為代表的蠕蟲對于免疫反應的調節功能。Chauhan等[21]發現,中帶絳蟲(M.Corti)的排泄/分泌因子(稱為寄生配體,PL)通過阻斷電壓依賴型離子通道TRPM2的激活、AMP依賴的蛋白激酶(AMPK)和自噬來抑制ROS刺激下的NEYosis,PL是中性粒細胞功能的重要生物調節因子。另一項關于線狀絲蟲的研究發現,線狀絲蟲分泌的、均一的、含有磷酸膽堿的糖蛋白ES-62通過下調MyD88基因表達阻斷TLR2/4接頭-轉導分子驅動的炎癥反應[22]達到免疫逃逸的效果。寄生蟲也能促進NETs的產生:瘧原蟲感染的紅細胞釋放巨噬細胞移動抑制因子(MIF),通過依賴于CXC趨化因子受體4(CXCR4)的機制引起PAD4介導的NETosis[20]。
1.5 活化的血小板與NETs 靜息循環中,血小板是血液中的哨兵。血小板激活后,可以通過脫顆粒統一釋放PAMPs,如高遷移率族蛋白1(HMGB1)、血小板因子4(PF4)和調節活化正常T細胞表達和分泌的趨化因子(RANTES,如CCL5)等,與其在中性粒細胞上的配體結合間接促進NETs的形成[23],也可以通過物理作用直接激活中性粒細胞。血小板-中性粒細胞直接結合是血小板驅動Netosis的重要環節。對于人類的研究發現,中性粒細胞表達的整合素MAC-1可以通過vWF因子與血小板異二聚體糖蛋白1ba(GP1ba)結合,是體外誘導NETosis的關鍵步驟[24]。
2 NETs的病理生理作用及其臨床應用
急性感染期間,NETs可以通過限制傳播、物理捕獲和破壞分解等方式傳播來發揮其抗菌活性,如表2。細胞外染色質表面的負電荷可以與白念珠菌和利什曼原蟲等生物體結合[24],組蛋白則以正電荷結合病毒包膜蛋白上的負電分子[19]。被吸附后的微生物會被高濃度的抗菌蛋白(如組蛋白、組織蛋白酶G、腸毒素、MPO等)殺死或滅活。此外,NETs也可以作為PAMAs,進一步激活抗炎反應,如前文所述HMGB1、PF4、CCL5和組蛋白等物質可以觸發包括中性粒細胞本身在內的免疫細胞釋放更多的細胞因子和趨化因子[25]。

表2 NETs組分的臨床應用研究
然而,過度擴大的免疫反應伴隨的往往是糟糕的病理結果。NETs的調節異常,無論是過度產生還是降解不足,都有可能帶來組織損傷、高凝狀態乃至血栓形成的嚴重后果,如與肺損傷相關的急性呼吸窘迫綜合征和與凝血異常有關的彌散性血管內凝血均已被發現與NETs的過度產生有極大的關系[25]。組蛋白釋放后可結合核苷酸結合寡聚化結構域2(NOD2)和免疫球蛋白結構域4(VSIG4)或核苷酸結合的寡聚化結構域樣受體蛋白3(NLRP3),進而激活胱天蛋白酶1(caspase-1)、制孔蛋白(GSDMD)引起細胞焦亡[27]。還可以通過補體發揮作用,引起包括膿毒癥心肌病在內的一系列并發癥。
NETs還是免疫血栓形成的重要啟動物質,其結構成分可以直接激活血小板,促進血栓形成,并抑制纖溶和天然抗凝途徑。血栓形成特別是膿毒癥相關免疫血栓形成被認為是膿毒癥不良結局的重要因素。從膿毒癥病人身上收集的1.5~10.0 kbp的雙鏈DNA大片段與Ⅻ因子和高分子量激肽原結合可以加強凝血反應。組蛋白可以通過TLR2和TLR4激活血小板并增加凝血酶的生成,誘導釋放多磷酸[24],暴露磷脂酰絲氨酸和激活V因子,這些變化反過來促進凝血酶原復合物的組裝[30]。組蛋白還可以通過激活調節蛋白C,進一步發揮血栓前作用。在體外,組織蛋白酶G和NE可以降解人內皮細胞上的組織因子途徑抑制物(TFPI)進一步增強凝血。DNA與纖溶酶和纖維蛋白形成三元復合物,改變血漿凝塊中的纖維蛋白結構發揮抗纖溶作用。NETs還可以捕獲TFPI,是唯一能有效抑制重要的蛋白水解酶復合體、Tf-FⅦa和凝血酶原酶活性的內源蛋白[31]。
隨著NETs病理作用逐漸被肯定,其在臨床診療過程中的應用也逐步被開發出來。對其組分濃度的監測使人們得以窺見膿毒癥發生、惡化的趨勢,特別是瓜氨酸化組蛋白H3(CitH3)、血清鈣衛蛋白(S100A8/A9異源二聚體)等特異性相對較高的指標被發現可能是膿毒癥診斷和預后的新型生物標志物[26,32]。Pan等[26]發現,CitH3對感染性急性胰腺炎(AP)和非感染性AP的診斷能力優于PCT,并提出CitH3濃度>43.05 ng/L作為截斷值區分病情嚴重程度和臨床轉歸,曲線下面積(AUC)及其95%CI為0.93(0.86,1.00)。血清鈣衛蛋白作為一種抗菌促炎蛋白復合體,占中性粒細胞胞質中可溶性蛋白的60%,被認為是診斷多種炎癥性疾病的生物標志物[33],基線濃度與膿毒癥病人的總生存期成反比。NE2和CitH3,以及內皮衍生生物標志物Syndecan-1均被發現與膿毒癥所致DIC相關[28]。Keir等[29]嘗試探索以NETs的濃度指導靜脈用藥,發現病人在接受大環內酯類藥物治療后,NETs(如組蛋白-彈性酶復合體等)濃度與銅綠假單胞菌感染病人病情惡化的減輕有關。
目前NETs的檢測主要是通過顯微鏡、ELISA、蛋白質印跡法、流式細胞術等方法,識別、命名和量化的不一致大大阻礙了臨床應用,甚至出現了細胞組學高含量篩選(HCS)的新方法,這將大大促進NETs的相關研究及臨床使用。
總之,NETs在早期控制和促炎反應方面發揮了局限、分解和促炎的重要作用,同時其過度產生也是過度炎癥反應并發生免疫功能紊亂、器官功能障礙的重要環節。因此研究者們嘗試通過抑制過度的NETs釋放來改善膿毒癥的病情進展和預后。
3 治療新思路
由于NETs的過量產生在膿毒癥病理過程中占有重要地位,因此,最近研究者們一直在努力尋找抑制NETs的形成或消除多余的NETs的潛在治療方法,如表3。相關研究發現NETs聚集可能會加重膿毒癥,同時阻塞胰管導致胰腺炎;而阻斷NETosis已被證明可以改善癥狀和預后[34]。雖然NETosis隨激活物質的變化存在有極大異質性,但絲氨酸蛋白酶途徑(如Raf-MEK-ERK通路)、NOX2-ROS途徑及核物質解聚的各種酶類(如NE、PAD4、MPO或去乙酰化酶等)都是相對統一、固定的,這也是近年來抑制NETs形成的研究熱點。抑制不同的位點可能帶來相似的效果。早期靜脈注射氨甲環酸可能通過經典的NE/MAPK途徑有效地抑制NETosis。針對PAD的研究發現抑制PAD2或PAD4均能顯著減少膿毒癥誘導的NETs成分[26],Cl-脒抑制PAD4阻斷組蛋白瓜氨酸化,進而阻止脂多糖(LPS)或PMA介導的NETosis。同樣抑制HDAC也被證明可以治療膿毒癥:鋅依賴賴氨酸脫乙酰酶是Ⅰ/Ⅱb類HDAC,是誘導NETosis所必需的,其抑制劑ricolinostat被證明可以在不影響其他抗炎反應的基礎上,有效抑制NETs的過量釋放,并已進入藥物Ⅱ期臨床試驗[12]。另外第二代NE抑制劑Sivelestat已在日本和韓國用于與全身炎癥反應相關的急性肺損傷(ALI)病人的臨床應用,其新型載體ICMV(ICMV-SIVE)更容易被中性粒細胞吸收,可能具有更好的療效[35]。

表3 膿毒癥治療新方案
傳統醫學也提供了新思路:姜酮治療可抑制核因子2(Nrf2)介導的ROS形成而明顯減輕NETs形成和炎癥反應,同時保持中性粒細胞的吞噬和趨化活性[36-37]。川芎中的主要成分刺五加內酯Ⅰ(SEI)顯著減少了NETs的形成,對CLP小鼠肺損傷有保護作用[38]。
還有一部分研究嘗試從血小板-中性粒細胞相互作用的角度進行突破:氯吡格雷、普拉格雷、替卡格雷和康瑞爾等P2Y12抑制劑通過抑制血小板P-選擇素的表達抑制血小板-白細胞相互作用[39]。外源性的重組血栓調節蛋白(RTM)可以抑制LPS模型中NETs形成,抑制除IL-1β外的所有細胞因子升高,提高膿毒癥存活率[40]。
已有的研究肯定了通過NETs尋找膿毒癥相關過度炎癥反應的治療的可行性和科學性,探索既能抑制過度炎癥反應又不影響正常免疫細胞吞噬作用的新型治療方案可能是未來膿毒癥相關治療研究的重點突破方向。
總之,膿毒癥是一種促炎反應與抗炎反應同時亢進的高度異質性綜合征。中性粒細胞作為先天免疫的第一道防線,外化核及胞質內容物是其發揮作用的重要途徑。NETs本身的產生及釋放可以為病情的進展提供優良的“示蹤劑”,但過度的NETosis也可能成為免疫紊亂的罪魁禍首,尋找抑制NETs過度形成而不影響正常中性粒細胞吞噬、消化功能的藥物,以及恰當的用藥時間和劑量可能是膿毒癥治療的新方向。
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
遼金歷史與考古(2019年0期)2020-01-06 07:45:20
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
電子制作(2018年11期)2018-08-04 03:26:04
汽車工程學報(2017年2期)2017-07-05 08:13:02
國際商務財會(2017年8期)2017-06-21 06:14:14
電子制作(2017年23期)2017-02-02 07:17:19
