光照在甲殼動物養殖中的應用研究進展
2024-02-21 07:38:28牛林洋董宏標鄭曉婷曾祥兵譚澤加張家松
漁業現代化 2024年1期
牛林洋,董宏標,鄭曉婷,曾祥兵,譚澤加,張家松
(1 中國水產科學研究院南海水產研究所,農業農村部南海漁業資源開發利用重點實驗室,廣東 廣州 510300;2 水產科學國家級實驗教學示范中心 (上海海洋大學),上海 201306;3 海南省海洋漁業資源高效利用與加工重點實驗室,海南 陵水572426;4 廣州利洋水產科技股份有限公司,廣東 廣州 510080)
光照是影響水產養殖生產的重要環境因子之一,有3個核心要素:光譜、光照強度、光照周期[1]。光譜為光通過光柵或棱鏡進行分光后,按照波長或頻率大小依次排列形成的圖案[2]。常用人工光源(高壓鈉燈、熒光燈、金屬鹵化物燈)通過添加濾光片控制光色,隨著LED燈的發展,光的波長可以由計算機控制,具有精度高、使用壽命長的特點[3]。光照強度是指單位面積上接受的光通量[4]。由于介質對光的吸收和散射,光照強度在傳播時會衰弱[5]。自然界光照周期由光周期和暗周期構成,動物根據不同的光照周期,進化出不同的生物節律[6]。
在影響甲殼動物養殖的諸多環境因子(光照、溫度、pH、鹽度、營養等)中,光照為關鍵因子之一,對甲殼動物生長發育、行為、攝食、存活和繁殖等具有重要影響[7]。水體中甲殼動物會因接收光照信號強弱而產生上浮和潛底等行為[8]。有研究表明,補充額外光源可促進幼蝦攝食,加速生長,進而縮短養殖時長[9],適當延長光周期能顯著提高甲殼動物幼體攝食率和存活率,保證幼體生長穩定性[10]。因此,提供適宜的光源,對養殖動物苗種培育與養殖生產具有重要現實意義。
本研究從光譜、光照強度、光照周期三方面進行闡述,綜合了近年來光照參數對甲殼動物行為、攝食、生長發育及產卵繁殖等方面的研究成果,以期為甲殼動物養殖提供理論及實踐參考。
1 光在養殖水體中的傳播特點
光源指能發出可見光的物體,可分為自然光源及人工光源[11]。光環境的構成包括光譜成分、光照強度和光照周期。光在水體與空氣中傳播有較大差異,空氣因密度小而對光的吸收與散射相對較小,水體密度為空氣的800多倍,因而對可見光有嚴重的吸收和散射[12]。水體對于自然光的吸收在不同光譜區域各有差異,水體對光譜中紫外(10~380 nm)和紅外(780~2 526 nm)部分具有吸收作用,而在可見光譜區段(380~780 nm),對紅光(622~780 nm)、黃光(577~597 nm)和淺綠色光(565~577 nm)吸收最強[13]。自然海水中,散射作用導致光波衰減60%,吸收作用導致光波衰減40%;而在養殖水體中,由于浮游植物的光吸收,紅光衰減得最多,藍光次之,綠光衰減最少[14]。甲殼動物養殖過程中,光波在養殖水體中的傳播會受到水分子、無機溶質、懸浮物等多種因素影響。相關研究表明,養殖池內海水的透光率隨水深的增加而呈現下降趨勢,當水深為30 cm時,綠光透光率最大,白光最小,隨著水深增加,綠光穿透率始終高于其余波長的光[15]。秦菲[16]研究同樣發現,LED光照在養殖水體中會隨水體深度的增加而衰減,此外,還會受到總懸浮固體(TSS)和化學需氧量(COD)的影響。根據不同光波在養殖水體中的傳播特性,實際生產中,選擇適宜的光照參數對甲殼動物養殖尤為重要。
2 甲殼動物的光感覺器官
普遍認為,甲殼動物具有單眼(ocellus)和復眼(compound eye)兩種光感受器[17],其中,單眼結構相對簡單,僅能感覺光線的強弱,不能呈現具體的像[18];而復眼是大多數甲殼動物成體的視覺器官,不同甲殼動物復眼的形態差異不大,如凡納濱對蝦(Litopenaeusvannamei)[19]、日本沼蝦(Macrobrachiumnipponense)[20]、鋸緣青蟹(Scyllaserrata)[21]、三疣梭子蟹(Portunustrituberculatus)[22]復眼均呈半球形。凡納濱對蝦復眼經H&E染色可顯示復眼由許多小眼構成,超微結構由外至內分為折光系統、感光系統和色素細胞三部分,其中折光系統包括角膜、角膜生成細胞和晶錐;感光系統由眾多小視網膜突起的微纖毛組成,微纖毛的排列順序在光、暗環境下不同,如羅氏沼蝦微纖毛在自然光環境下排列凌亂,而黑暗環境下則排列整齊[23];色素細胞則具有遮擋光線的作用[17]。甲殼動物的感光單元結構為感光細胞,由感光部、細胞體部和軸突構成[24]。感光細胞通過吸收和釋放存在于潴泡囊和線粒體中的Ca2+改變其體積,光敏感性隨著Ca2+濃度升高而降低[25-26]。甲殼動物對光色的敏感程度與其色素細胞對光色的過濾性相關[27];自然光環境下,日本沼蝦視覺細胞中色素顆粒分散到細胞中,感桿束邊緣小囊泡數量增多,溶酶體出現并靠近感桿束[28],這與中華絨螯蟹(Eriocheirsinensis)光環境下視覺細胞觀察結果相似[29]。Sanudin等[30]研究發現,凡納濱對蝦幼體攝食主要依賴復眼進行,光照是影響其幼體攝食的主要因素之一,隨著不斷生長發育,復眼的分辨力逐漸增強,其主動攝食的能力亦逐步提高;進入成體階段后,復眼在探測食物過程中的作用逐漸減弱,而由化學感受器起主導作用[31]。
甲殼動物眼柄神經內分泌系統為X器官-竇腺復合體(XO-SG complex),是甲殼動物內分泌調節中心[32]。凡納濱對蝦復眼經H&E染色顯示(圖1)[19]。
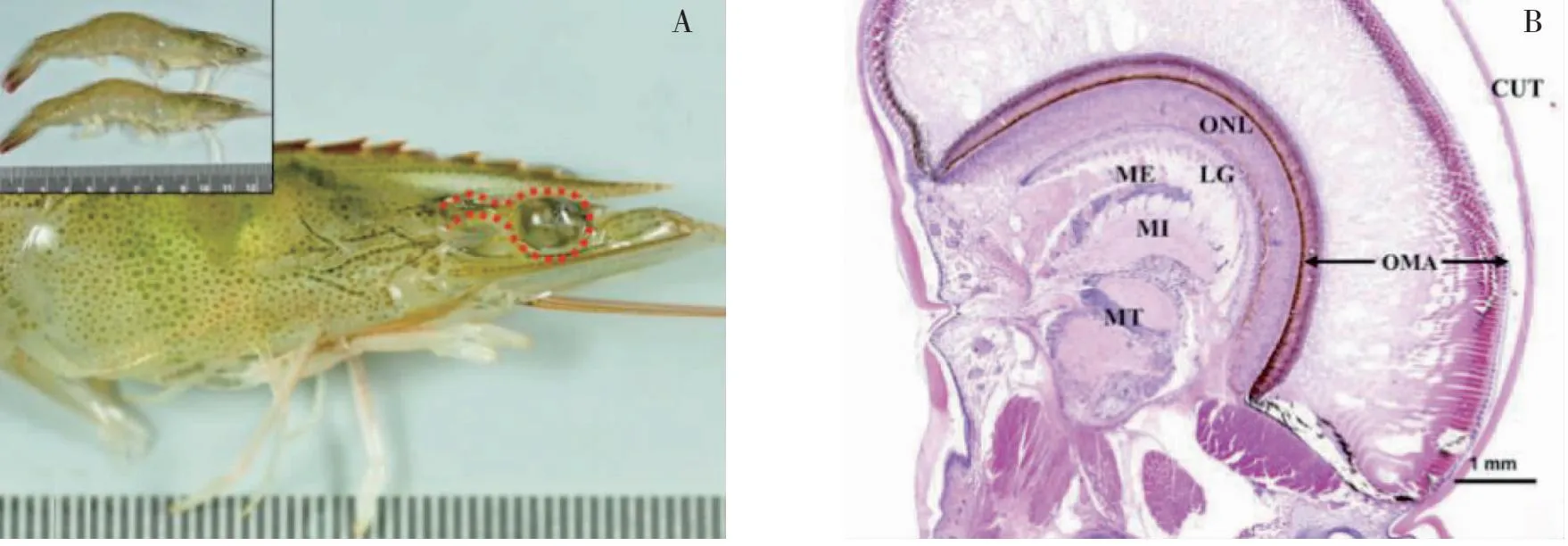
注:H&E,染色顯示的結構;CUT,角質層;OMA,小眼;ONL, 視神經層;LG,神經節層;ME,外髓質;MI,內髓質;MT,終髓
從最外層角質層向內依次為角膜、小眼、視神經層,神經節層、外髓質、內髓質、終髓。竇腺位于內髓和終髓之間,儲存蛻皮、生長、性成熟和代謝等相關激素[33]。光照的變化會引起甲殼動物體內激素或神經遞質水平的變化,進而影響其生理狀況[34]。Nakamura[35]研究發現,光照能夠通過影響視神經脊髓終端細胞活性以調節日本囊對蝦(Marsupenaeusjaponicus)攝食節律。此外,連續光照能夠引起克氏原螯蝦(Procambarusclarkii)眼柄、大腦、丘腦神經節、食管下神經節中5-羥色胺(5-HT)濃度顯著升高,進而刺激克氏原螯蝦的卵母細胞成熟[36]。
3 光照對甲殼動物養殖生產的影響
3.1 光譜成分對甲殼動物的影響
甲殼動物對不同波長的光敏感程度因種而異,而同物種不同的生長階段對于光譜的敏感度也有差異[37]。據報道,摘除觸角的日本沼蝦在紅光和綠光光照區域的平均攝食量遠大于藍光和黃光區域,其趨光行為在紅光下頻率最高[38]。克氏原螯蝦擁有晝夜兩套感光系統,其紅光感光系統發育形成較藍光感光系統緩慢,進而導致幼體階段對藍光敏感,成蝦階段則對紅光敏感[39]。長毛明對蝦(Fenneropenaeuspenicillatus)同樣具有雙重光感受器,分別對藍光和紅光兩種波長的光敏感[40]。另有研究發現,相同光照強度下,三疣梭子蟹在紅光下的趨光率最高,黃光下趨光率最低[41]。
不同種類甲殼動物適宜生長的光譜環境有所差異。Emmerson等[42]研究發現,藍光和綠光可改善印度明對蝦(Fenneropenaeusindicus)成蝦的生長特性。采用金鹵燈為光源養殖凡納濱對蝦時,與其他光源組相比,不延長光照時間就能顯著提高對蝦體內蝦青素含量,改善其體色,增加對蝦攝食率并促進生長[10]。表明養殖過程中補充光源時選擇適宜的光譜有助于促進養殖對象生長。
光照是除營養外影響甲殼動物繁殖與幼體發育的另一重要因素[43]。黑暗條件下紅條鞭腕蝦(Lysmatavittata)親蝦的抱卵率為0,自然光照下抱卵率可提高到(95.8±3.6)%,適宜的光照能夠對紅條鞭腕蝦親蝦抱卵起到促進作用[44]。光譜對甲殼動物繁殖的影響亦是因種而異。研究發現,藍光環境能夠促進東方巨對蝦(Penaeusplebejus)卵巢發育[45];綠光和藍光協同條件下,印度明對蝦產卵量顯著高于自然光條件[42]。光譜對主要甲殼動物生長生理的影響如表1所示。
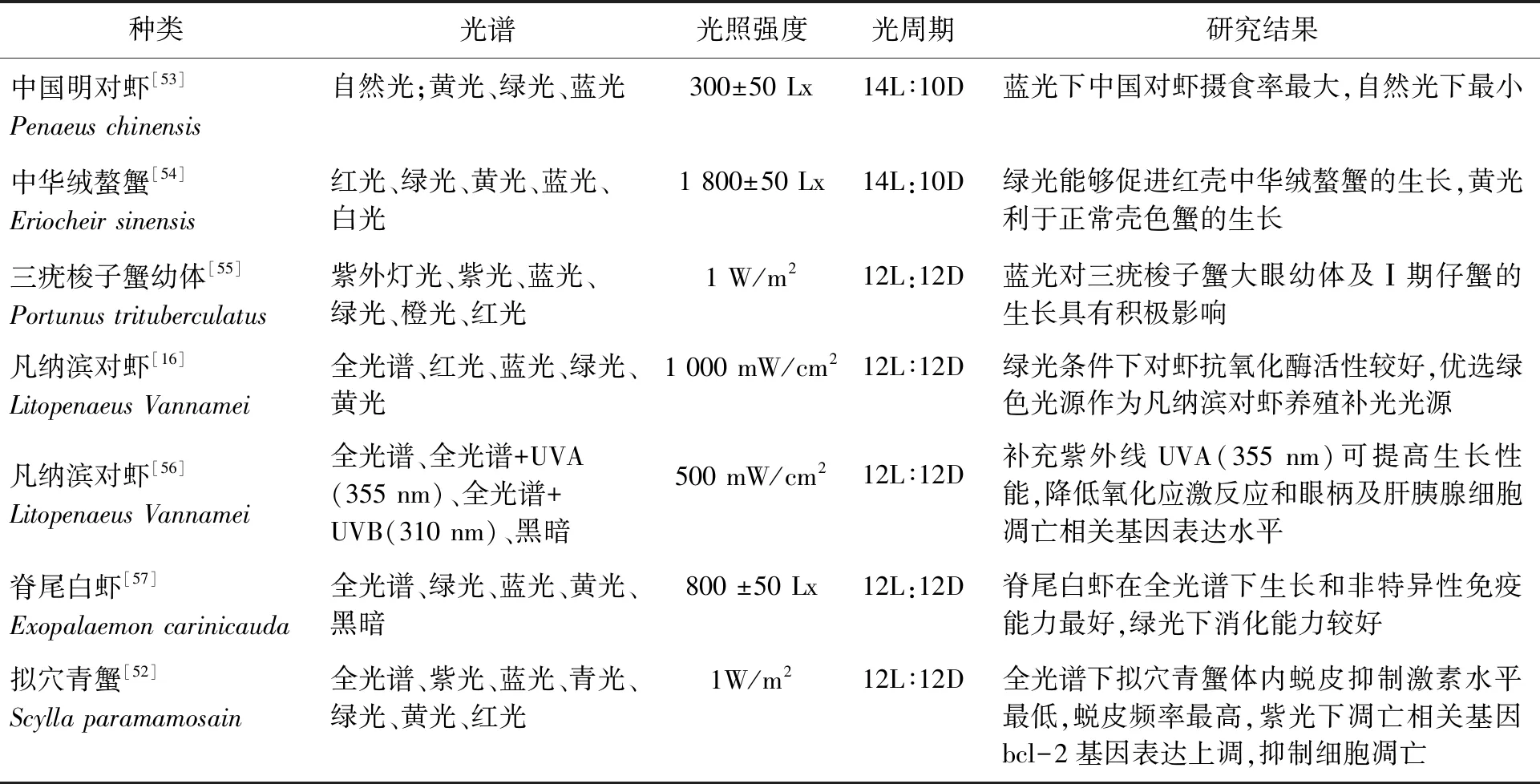
表1 光譜對主要甲殼動物生長影響
光譜通過影響甲殼動物的生理機能調控其生長與免疫。研究表明,甲殼動物的內在生理機制極易受到外界環境因素的刺激[46],一些甲殼動物如凡納濱對蝦、斑節對蝦(Penaeusmonodon)、中華絨螯蟹等在受到環境因素脅迫,機體內的活性氧(ROS)增加,常通過分泌超氧化物歧化酶(SOD)、過氧化氫酶(CAT)等抗氧化相關酶以分解體內過多的活性氧,減少機體損傷[47]。藍光條件下凡納濱對蝦胃蛋白酶(PEP)和胰蛋白酶(TRY)顯著高于其他光照組,藍光和綠光條件下脂肪酶(LPS)活性顯著高于其他組,結果表明,藍光和綠光通過調控消化酶的分泌,促進對蝦的消化吸收[48]。綜上,不同種類及生理階段甲殼動物需要提供不同的光譜范圍,使其能更快響應環境應激,維持內環境穩態,提高養殖效率。
光譜對甲殼動物蛻皮的影響與其對不同波長的光敏感性不同有關。甲殼動物蛻皮受內分泌系統和神經系統共同調節[49],同時也受到環境因子(光照、季節變化等)的刺激[50]。普遍認為甲殼動物蛻皮是位于頭胸部前端的Y器官分泌蛻皮激素(molting hormone,MH)與位于眼柄的X器官竇腺復合體分泌蛻皮抑制激素(molt-inhibiting hormone ,MIH)互相拮抗作用的結果[51]。全光譜下擬穴青蟹(Scyllaparamamosain)體內MIH水平最低,蛻皮頻率最高,紫光和黃光下MIH顯著上調,蛻皮頻率最低[52],表明光譜通過調控蛻皮相關激素水平,來影響甲殼動物的蛻皮頻率與生長。研究發現,凡納濱對蝦在藍光下蛻皮總次數顯著高于其他光譜組,且藍、綠光下對蝦的生長速度較快[48]。
3.2 光照強度對甲殼動物的影響
不同種類與生長階段的甲殼動物最適光照強度具有差異,其行為活動因光照強度而發生改變。已有研究表明,幼體階段的甲殼動物對光照具有正反應,而成體階段對光照則呈負反應[58]。光照會影響甲殼動物的垂直移動行為,白天光照強度減弱和夜晚的光照強度增強均會引起對蝦的上浮或潛底[59]。如三疣梭子蟹喜弱光,懼強光,白天多潛伏在海底層或活動于靠近底層的水域,夜間或生殖抱卵期才游到上層[60]。自然條件下,大多數種類對蝦同樣于白天潛底,夜間上浮[61]。然而Kenyon等[62]研究發現,仔蝦的上浮和潛底可能不受光照強度的影響,褐蝦(Penaeusesculentus)仔蝦在光暗環境下皆呈浮游狀態,而不下潛。這與褐蝦仔蝦的復眼結構與對環境的高適應性有關。甲殼動物的攝食、發育及蛻皮等活動與其行為變化都有相關性。
關于光照強度與甲殼動物攝食之間的關系,Petersen等[63]提出了兩種攝食模型:峰值攝食模型曲線和S型攝食模型曲線,其中,符合峰值攝食模型曲線的動物主要依賴視覺進行攝食,存在一個適宜的光照強度范圍使動物的攝食量達到最大,光照強度過高或過低都會影響動物的攝食;S型攝食模型曲線為隨著光照強度的減弱,動物的攝食量逐步增高,當光照強度降低到一定程度,攝食量達到最大且基本恒定。大多數蝦蟹類動物的攝食模型屬于峰值攝食模型。無論是處于哪一期的羅氏沼蝦幼體,在完全黑暗的條件下都幾乎不攝食,隨著光照強度的增大,攝食率迅速升高,而到1 000 Lx以上攝食率變化不大[64]。研究發現高光照度下(5 000 Lx)中國明對蝦的呼吸、排泄耗能較高,而用于生長的能量比例降低[65]。中國對蝦在夜間的攝食量要高于在日間,且在夜間有兩個攝食高峰,分別是18:00—21:00和03:00—06:00[66]。養殖車間需要根據相應物種攝食所需光照強度靈活調整光照條件,育苗階段尤為重要。
光照強度通過影響食物轉化效率和能量分配比例調控對蝦生長發育[67]。中國明對蝦在5 500 Lx照度下的特定生長率(SGR)和食物轉化率(FCE)最低,在300 Lx照度下最高[53]。Neal等[68]研究發現,自然光條件下池塘養殖凡納濱對蝦養殖產量比弱光池塘中高出48%,分析其原因可能在于光照對輪蟲和其他浮游動物的生長具有促進作用,為對蝦提供了天然的餌料來源。相同光周期(14L∶10D)下,光照度波動組(2 700±1 800 Lx)凡納濱對蝦的攝食率(FI)顯著高于恒定光照度組(2 700 Lx),而飼料轉化率(FCR)低于恒定光照組,且2 700±1 800 Lx光照度下凡納濱對蝦的生長狀況優于其他波動光強組[69]。養殖車間內適宜的光照強度對提高甲殼動物幼體存活率及攝食率有重要作用。
甲殼動物一般呈階梯式生長,即在蛻皮后快速生長,蛻皮之后到下一次蛻皮之前其尺寸幾乎很少增加[70]。對蝦的蛻皮和生長是結合進行的,但蛻皮是一種節律性極強的活動,主要由體內激素調節,生長則受到周圍環境中餌料的影響[71]。饑餓狀態下一些甲殼動物依舊能夠蛻皮,但蛻皮后的干重和能量比攝食組的少[72]。三疣梭子蟹幼體生長適宜的光照強度為9.96 μmol /(m2·s),該光強能夠保證幼蟹的存活率、蛻皮率及生長性能[73]。
當前,光照強度對于甲殼動物繁育影響的相關研究仍不足。研究表明強光(11 00 Lx)能夠抑制墨吉明對蝦(Penaeusmerguiensis)卵巢成熟并降低其產卵量,弱光環境(2 Lx)下則相反[74]。弱光條件下可以促進紅額角對蝦(Litopenaeusstylirostris)和凡納濱對蝦的繁殖[75]。交配后的擬穴青蟹在完全黑暗條件下其卵巢中類胡蘿卜素及蛋白質含量低于低光強(7.00±1.50 Lx)環境[76]。低光強或暗光條件下對大部分甲殼動物的性腺發育及繁殖有利,光強過高則不利于甲殼動物性腺的發育、成熟及產卵。光強對主要甲殼動物生長的影響如表2。
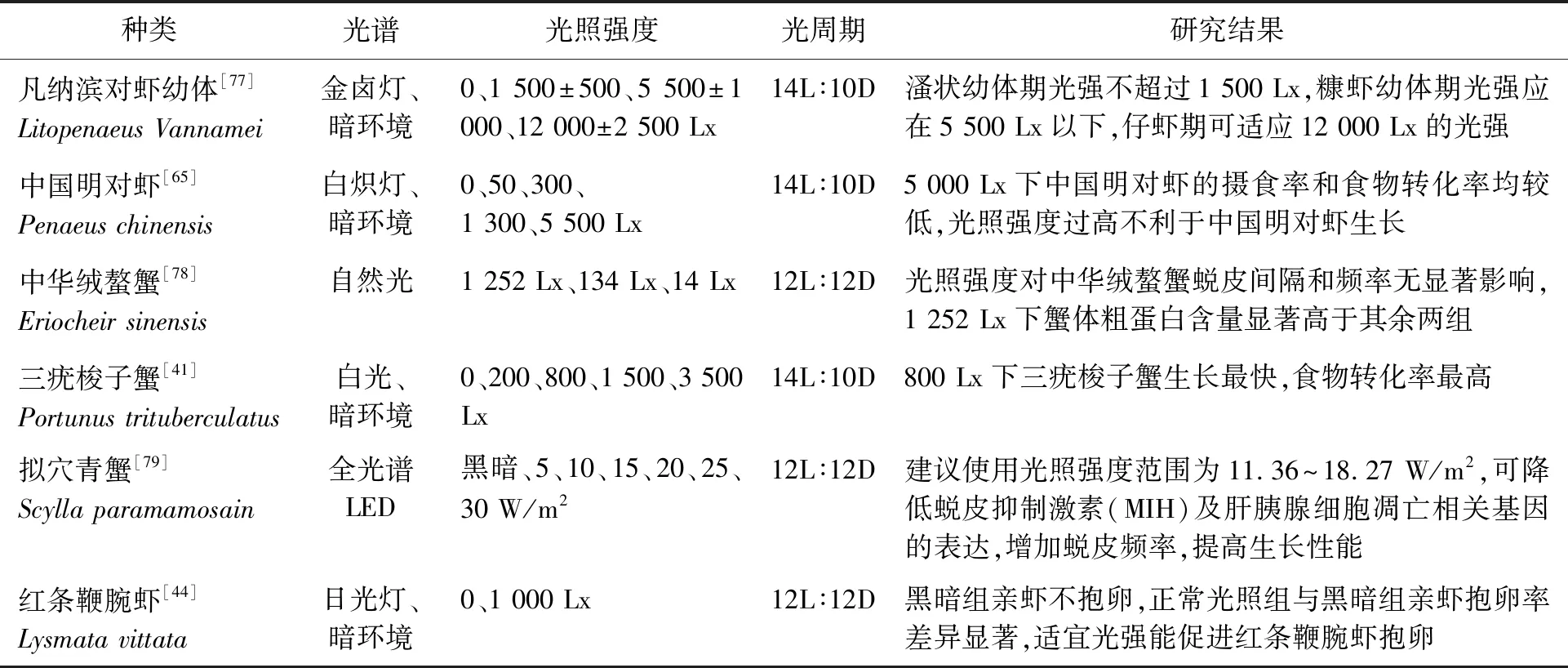
表2 光照強度對主要甲殼動物生長的影響
3.3 光照周期對甲殼動物的影響
生態環境中大多數的生物都經歷著晝夜交替。因此,許多生物也進化出了相應的生理感官和行為活動來響應這種晝夜變化并及時對自身行為做出調整,如感光器官和生物鐘機制[80]。普遍認為,眼柄組織是甲殼動物的特有感光器官,光線對其神經分泌細胞的分泌活動具有調控作用[81]。甲殼動物在不同光周期下的生長、代謝、存活和繁殖具有物種差異性[82]。
光周期是光環境中重要的因素之一,探究不同光周期下甲殼動物的攝食及生長很有必要。Sanudin等[83]研究發現,小規格(<0.5 cm)凡納濱對蝦培育時應延長光周期[12L(Light)∶12D(Dark)~24L∶0D],而大規格(>1 cm)對蝦的攝食行為、生長狀況和存活率不受光周期的影響。相關研究表明凡納濱對蝦養成期攝食活動受光周期影響較小,表明其攝食行為是一種內源性行為[84]。You等[10]研究發現,連續光照條件下,凡納濱對蝦的攝食率并沒有顯著的提高。在羅氏沼蝦養殖過程中,適當延長光照周期,能夠增加其日攝食量,促進幼體生長發育[85]。Andrés等[86]研究發現,持續黑暗導致遠海梭子蟹(Portunuspelagicus)幼體存活率和發育率降低,明確遠海梭子蟹幼體最適生長光周期為18L∶ 6D。
光照周期的改變對甲殼動物蛻皮同樣產生影響。延長光照周期會抑制甲殼動物蛻皮抑制激素(MIH)的合成和釋放,從而縮短其蛻皮周期,短時間光照則相反[87]。光照周期延長使日本囊對蝦體內MIH激素水平顯著升高,蛻皮激素(EH)水平下降,黑暗條件MIH濃度顯著低于其他光照組,EH濃度顯著高于光照條件,結果表明黑暗狀態有利于日本囊對蝦蛻殼[88]。黑暗條件下中華絨螯蟹大眼幼體蛻殼率降低、成活率低;光照可刺激并促進大眼幼體的蛻殼與生長,但在仔蟹I期以后黑暗條件下有利于其蛻殼生長[89]。
在甲殼動物生物學研究中,常通過調控光照周期來影響其產卵繁殖[90]。適當延長光周期(16L∶8D)可促進克氏原螯蝦雌蝦性腺發育,而過長的光照(20L∶4D)對雌蝦性腺發育會有抑制作用[91]。Matsuda等[92]研究發現,水溫為25 ℃時日本囊對蝦卵巢發育和產卵最佳光照時間是12.5~14 h。據Akinouchi 等[93]報道,光照度為2~5 Lx,光照周期為17L∶7D時,斑節對蝦的產卵量和孵化率均高于其他光照組。光照周期對主要甲殼動物生長的影響如表3所示。
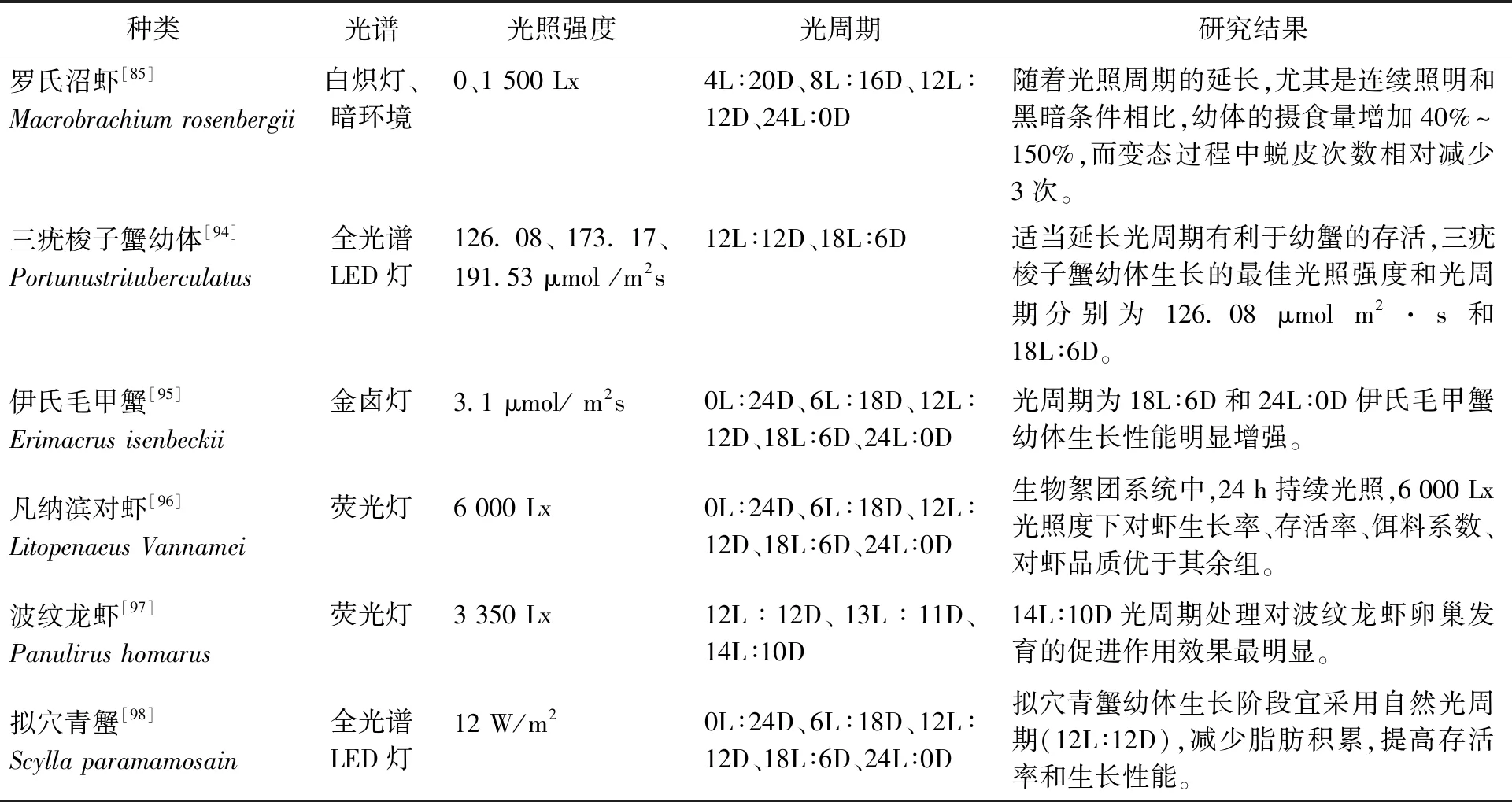
表3 光照周期對主要甲殼動物生長的影響
4 問題與展望
4.1 光照參數的優化
光環境是設施漁業生產系統構建需要重視的因素之一,其在調控水產動物幼體發育、攝食行為及產卵繁殖等方面具有重要意義,如何營造并利用好光環境是實現水產養殖高效生產的重要環節。
(1)光譜的選擇需要考慮到不同波長光照在養殖水體中的傳播特性,同時需要注意養殖對象對光照的敏感程度。自然光源及傳統人工光源發出的光譜范圍廣,缺乏對光譜的精準調控。(2)光照強度的設置對甲殼動物的攝食、生長及繁育有重要作用。不合理的光強將引起甲殼動物的應激反應,導致其生長受到阻滯。(3)光照周期對甲殼動物的攝食、蛻皮及產卵繁殖都有一定的影響。甲殼動物養殖是現代水產養殖中重要的組成部分,不同種類甲殼動物的最適光環境參數都有差異,生產過程中需要根據品種特性靈活調整光照參數。此外,光照參數對甲殼動物內分泌系統中各種激素和神經遞質的生理調節仍然需要更為全面深入的研究。結合代謝組學、多組學、表觀遺傳學、全基因組關聯分析等技術,全方面探索光照對甲殼動物相關基因調控的具體分子通路和作用機制,可作為未來探究光照對甲殼動物生長、發育、繁殖等的研究方向。
4.2 光源的選擇
隨著設施漁業智能化發展,工廠化養殖將是漁業智能化發展的重要方向。相較于傳統養殖方式(池塘養殖、網箱養殖)具有節水節地、高產高效、生態環保、全程可控等優點。目前,大多數工廠化養殖車間用的還是傳統光源,例如金鹵燈、熒光燈等,這些光源不僅耗能高,長時間照射還會產生較高的溫度,而且傳統光源還含有對養殖動物生長不利的光譜成分,尤其會對甲殼動物的生長造成一定的負面影響。自20世紀90年代以來,國內外學者圍繞光照對水產動物的影響逐漸開展相關研究,從傳統光源對水生動物的影響逐步轉變為更加精準的光譜、光周期和光照強度的調控,人們逐漸意識到LED光源對水產養殖的增益效果。使用LED養殖燈對甲殼動物生長光環境進行精準調控是未來對蝦養殖效率的關鍵提升環節之一。設計針對養殖動物各生長時期(如對蝦標苗期、標粗期、養成期、繁殖期等)的LED補光策略,以滿足養殖動物對光照的需求。目前,大多數養殖車間對于LED補光系統中光照參數的搭配設計不完善,造成資源、成本的浪費。因此,建立設施漁業系統中對光譜、光強、光周期的需求體系,是未來推廣智能化設施漁業的重要步驟之一。
5 結論
水產養殖中,養殖水體直接影響光的傳播,甲殼動物接收到的光是經過空氣與水體散射和吸收過后的,相關研究表明,綠光與藍光在養殖水體中的穿透性較好。本綜述中,多項研究表明,綠光與藍光能夠促進甲殼動物的攝食、幼體生長、蛻皮、產卵,增強抗氧化能力。不同甲殼動物對光強的適應性有所差異,如中國明對蝦在300 Lx光照度下生長最快,三疣梭子蟹在800 Lx下生長最快,凡納濱對蝦能夠適應較高的光照度,可達到12 000 Lx。甲殼動物產卵階段,應使用較低的光強,有利于其卵巢發育。適當延長光照周期(如18L∶6D),對大多數甲殼動物的生長及產卵繁殖均有利,而不同種類甲殼動物的蛻皮行為對光照周期的響應有差異。實際生產階段,應兼顧光照三要素的作用,不同時期選擇合適的光照參數,以適應動物的生長。
□
猜你喜歡
中學生數理化·八年級物理人教版(2022年3期)2022-03-16 05:55:08
當代陜西(2021年2期)2021-03-29 07:41:24
小讀者(2021年2期)2021-03-29 05:03:48
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
媽媽寶寶(2017年3期)2017-02-21 01:22:28
紅領巾·萌芽(2016年1期)2016-09-10 07:22:44
中國塑料(2016年3期)2016-06-15 20:30:00
