東北春大豆種質資源表型分析及綜合性評價
2024-02-20 00:00:00楊祥波陳亮宇楊松楠陳喜鳳邢偉明李雪瑩叢煒軒臧振原臧遠波張君
新疆農業科學 2024年12期
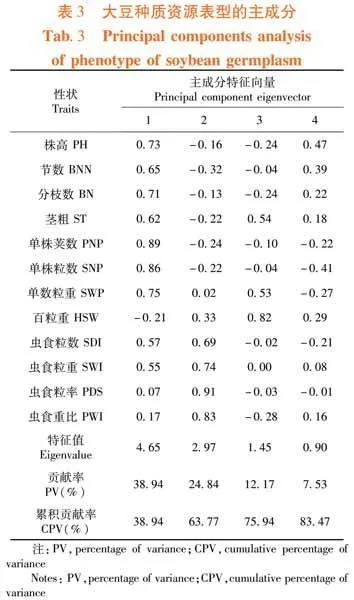
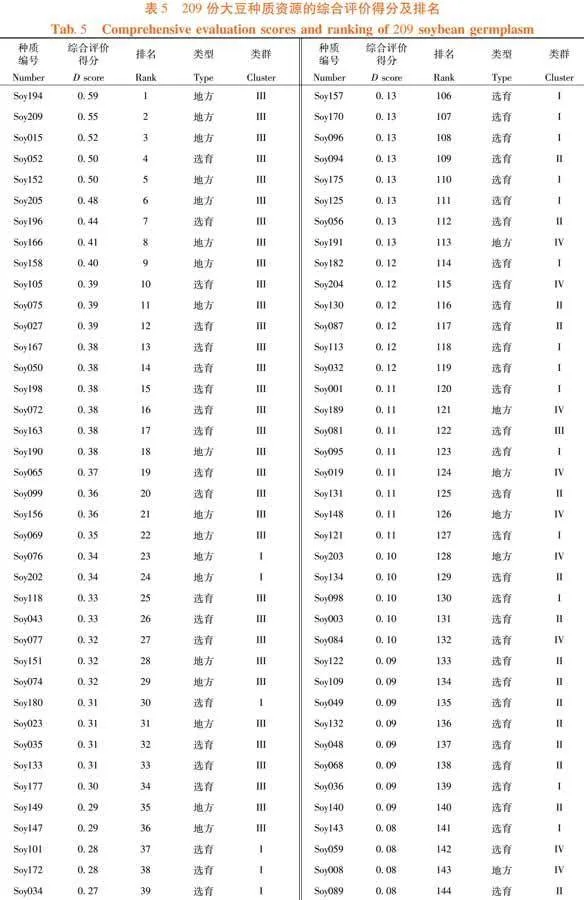
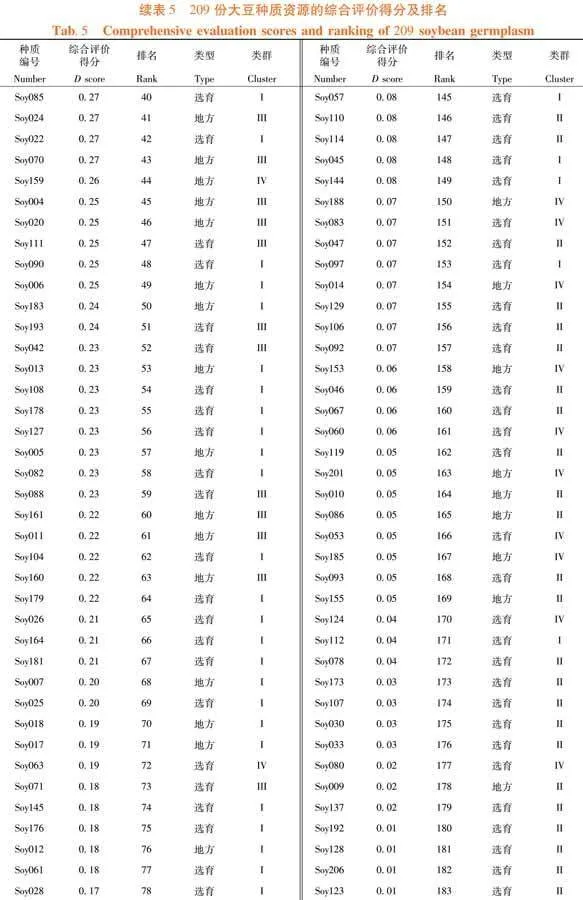

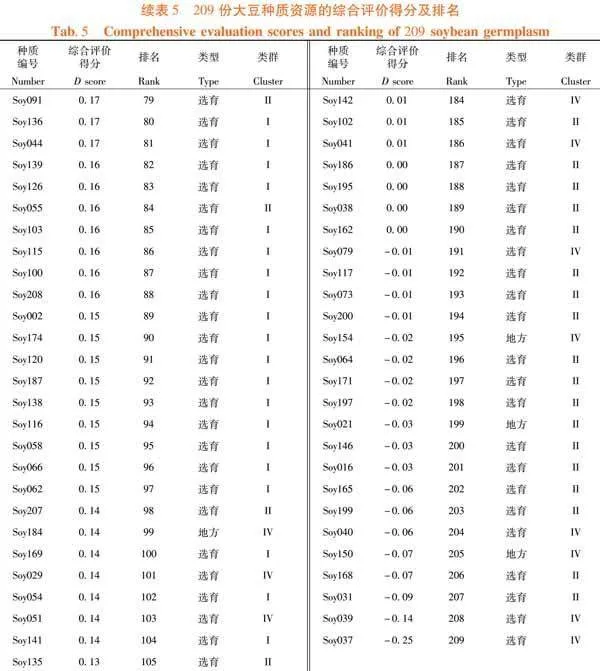
摘 要:【目的】分析東北春大豆種質資源的表型特征并篩選優質材料。
【方法】選用209份材料,種植3年,對12個大豆性狀進行描述性統計、聚類分析、分段線性回歸分析和綜合評價等。
【結果】表型變異系數及多樣性指數范圍指出節數、莖粗和百粒重變化幅度較為穩定,分枝數、蟲食重比和單株莢數等9個性狀變異豐富;所有性狀可被主成分分析濃縮為產量、抗蟲、百粒重及株型因子,其中產量因子與株型正相關,與百粒重、蟲食粒數和蟲食重比負相關;種質資源可劃分為4個類群,I類群株型適中且莢數多,II類群植株矮小且單株產量低,III類群株型理想、產量最高,IV類群與I類群相似但抗蟲性差;分段線性回歸得出理想高產大豆范圍在株高102.03 cm、19.93個節數、3.5個分枝、莖粗11.39 mm、豆莢99.73個和粒數286.09個左右,且盡可能不受大豆食心蟲的取食;在DTOPSIS(dynamic technique for order preference by similarity to ideal solution)法下,種質資源的綜合得分在0.59~0.25,平均得分為0.16。
【結論】東北春大豆資源表型多樣性豐富,I類群、III類群可分別用于株型改良和高產育種。株型適中、粒數較多且抗蟲是高產大豆的主要特征。Soy194、Soy052和Soy196可作為優異種質運用于大豆育種中。
關鍵詞:大豆;農藝性狀;聚類分析;分段線性回歸;綜合評價
中圖分類號:S565.1"" 文獻標志碼:A"" 文章編號:1001-4330(2024)12-2921-13
0 引 言
【研究意義】大豆(Glycine max)是我國主要的糧油飼兼用作物[1]。提高大豆單產是育種者的主要目標之一[2, 3]。種質資源的收集與表型評定是挖掘優良材料、提高種質利用效率以促動大豆產量增長的關鍵[4, 5]。
東北地區作為大豆的主產區,評估當地種質資源的性狀表現有助于發掘影響產量的重要性狀因子及優異材料,對培育高產大豆具有重要意義。
【前人研究進展】趙朝森等[6]在江西省對439份大豆材料進行調查研究,在評估其19個性狀多樣性的同時,發現蛋白質含量、蛋脂總含量高而脂肪含量低是江西大豆種質的突出特點并篩選出3份高蛋白材料;徐澤俊等[7]基于303份黃淮海地區大豆種質資源內13個表型,認為生育日數、株高、單株粒數、單株粒重和蛋白質含量是該區域主要評價指標;聶波濤等[3]以334份北方春大豆的12個性狀作為對象,不僅探討了株型因素及品質性狀對單產的影響方向,而且計算出高產高油及高產高蛋白理想株型的性狀指標范圍。地方大豆品種是種質創新的基礎材料之一[8],亦往往攜帶當地主要病蟲害抗性基因及較強的適應性[9, 10],將其納入考察范圍,即可用于高產品種的改良,也有助于突破大豆育種同質化的瓶頸[11]。此外,大豆食心蟲是影響東北春大豆產區的主要害蟲之一,其啃食籽粒以造成產量損失[12]。雖然在品種審定中將食心蟲害作為考量標準,但在近年來鮮見其作為評價指標的報道。這削弱了東北地區種質資源評價的科學性及合理性。變異及多樣性分析、相關性分析、聚類分析等方法被廣泛用于大豆種質資源的評價系統中[6, 7, 13],尤其是主成分分析和聚類分析常用于對材料的分群,以總結其類群血緣、性狀特點以提供雜交組配的合理道路[13, 14]。【本研究切入點】種質的優異與否依賴對測量性狀的綜合評價。DTOPSIS法因其綜合了性狀方向選擇、對性狀的客觀賦權及基于現有對象的相對優劣評價的優點,被陸續用于玉米[15]、谷子[16]和羊草[17]等優異育種材料的選擇,為作物表型綜合評價提供了科學合理的量綱方式。需綜合209份東北地區春大豆的選育和地方材料,對農藝性狀和抗食心蟲性狀進行多年鑒定,以完善大豆種質資源評價體系并篩選優異種質。【擬解決的關鍵問題】評估材料群體表型多樣性、相關性及單株產量影響因子、篩選高產抗蟲的大豆種質資源,為東北地區大豆新品種的選育提供有益資源與表型參考。
1 材料與方法
1.1 材 料
選用209份在吉林省可正常成熟的大豆種質資源,由吉林農業大學農學院經濟作物改良團隊提供。選育材料156份,其中154份為國內材料,2份為國外材料(以下均簡稱選育);國內地方材料53份(以下簡稱地方)。
所有材料2021~2023年種植于吉林省長春市吉林農業大學大豆區域技術創新中心試驗田(43°80′N,125°41′E)。每份材料等量點播為一行,行長4 m,行距0.6 m,株距8 cm。田間管理及農藝措施如同常規農業生產方式。依據不同種質資源的生育期特點,在其完全成熟后收獲,用于后續表型考察。
1.2 方 法
1.2.1 試驗設計
選取每行中連續生長且長勢良好的3株植株進行室內考種。株高(Plant height / PH)、節數(Branch node number / BNN)、分枝數(Number of branches / BN)、莖粗(Stem thickness / ST)、單株莢數(Pod number per plant / PNP)、單株粒數(Seed number per plant / SNP)、單株粒重(Seed weight per plant / SWP)和百粒重(One hundred seed weight / HSW)依據《大豆種質資源數據質量標準(GB 4404.2-2010)》鑒定,但單株莢數記錄癟莢等無效莢,單株粒重不淘汰蟲食粒。大豆食心蟲造成的蟲食粒數(Seed damaged by insect / SDI)及蟲食余粒重(Damaged seed weight by insect / SWI)在考察粒數及粒重時清點,而后用于計算蟲食粒率(Percentage of damaged seeds by insect of total seed number per plant / PDS)與蟲食重比(Percentage of damaged seeds weight by insect of total seed weight per plant / PWI),其計算公式為:
蟲食粒率 = (蟲食粒數 / 單株粒數)× 100%;
蟲食重比 = [(百粒重 / 100) × 蟲食粒數 - 蟲食余粒重] / 單株粒重 × 100%。
1.2.2 表型數據統計及遺傳多樣性
基于R 4.3.2(https://www.r-project.org/)利用所有種質資源的表型平均值進行描述性統計等分析。其中,方差分析使用agricolae包運算;聚類分析應用scale函數處理原始數據使之標準化,并采用歐式距離及離差平方和法進行[18, 19],NbClust包用于確認最佳聚類簇數[20];主成分分析則使用factoextra包和FactoMineR包對標準化后的數據進行,PCA函數下scale.unit設置成FALSE,ncp設置為10;分段線性回歸分析通過SiZer包對原始數據進行,piecewise.linear函數下CI設置為TRUE,bootstrap.samples設置為1 000。其余統計則利用R自帶函數及tidyverse包下的對應函數運算,所有統計結果的可視化基于ggplot2、ggtree(聚類樹)和corrplot包(相關性圖)繪制。此外,除上述標注的參數外,其余參數設置為默認情況。
Shannon-weaver多樣性指數(H′)用于衡量表型的遺傳多樣性[13, 21]。簡要而言,先以各表型的平均值(X-)和標準差(δ)將群體分為10個類群,第一類的數值 lt; X- - 2δ,第10類的數值 ≥ X- + 2δ,其他類群間相差0.5δ。隨后,計數每個類群的分布頻次。公式H′= -ΣPiLnPi被用于計算指數,Pi為第i個類群內的頻次數。
1.3 數據處理
使用基于主成分分析的DTOPSIS法對種質資源進行賦分。首先,依據生產實際將12個性狀分為正向、中性和逆向三個類型。其中,節數、分枝數、單株莢數、單株粒數和單株粒重有助于大豆單產的提高,歸為正向類型;株高和莖粗與植株的生物產量相關,但過高可能造成產量的降低,歸類成中性類型;蟲食粒數、蟲食粒重、蟲食粒率及蟲食重比反映大豆產量的損失,定義為逆向類型。針對不同類型,采用無量綱化處理[15]:正向類型Mij = Nij / Nimax,其中,i表示第i個表型,j表示第j個品種,Nij是j品種的表型值,max表示該表型的最大值;中性類型Mij = N-i / [ N-i + ABS(N-i - Nij) ],其中N-i表示平均值,ABS為絕對值;逆向類型為Mij = Nij / Nimin,其中,min表示最小值。其次,對無量綱化的數據進行主成分分析,選取對應性狀的特征向量使之同標準化后的表型數值進行乘積和運算并獲得F值[3, 13],依照模糊隸屬函數法Uij = (Fij-Fimin) / (Fimax - Fimin) 求得各種質資源下每個主成分的U值[3, 22]。最后,以主成分的貢獻率作為權重(W)分別與U值相乘獲得綜合評價得分(D)[3],即D = W1 × U1 + W2 × U2 + Wi × Ui。
2 結果與分析
2.1 大豆種質資源的遺傳多樣性
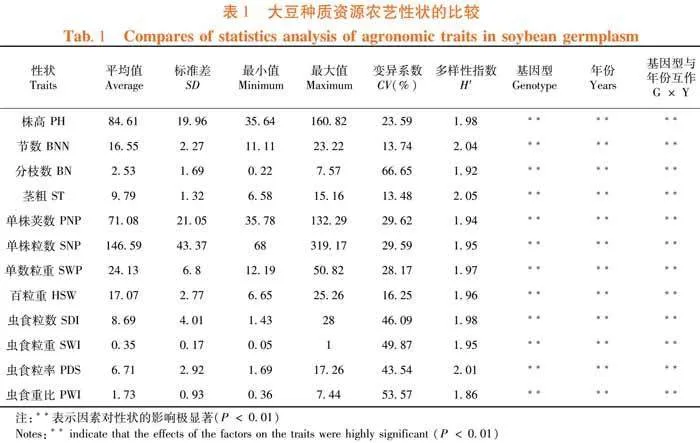
研究表明,各性狀的變異系數為13.74%~66.65%,從大到小依次為分枝數、蟲食重比、蟲食粒重、蟲食粒數、蟲食粒率、單株莢數、單株粒數、單株粒重、株高、百粒重、節數和莖粗。可見所用種質在抗蟲性及產量性狀上存在較大的差異,但百粒重的變異系數較小,僅為16.25%。多樣性指數的跨度為1.86~2.05,從大到小依次是莖粗、節數、蟲食粒率、株高、蟲食粒數、單株粒重、百粒重、單株粒數、蟲食粒重、單株莢數、分枝數和蟲食重比,上述性狀在各區間均有分布。此外,12個性狀均受年份(環境)的極顯著影響,同時還具有種質與年份間的極顯著互作。試驗所涉種質遺傳多樣性豐富,適合評價種質資源。表1
2.2 大豆各性狀的相關性
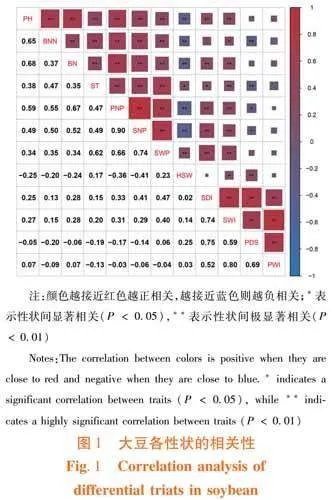
研究表明,株高、分枝數、節數、莖粗、單株莢數、單株粒數和單株粒重間兩兩呈現極顯著的正相關關系,但除莖粗及單株粒重外,這些性狀與百粒重為極顯著的負相關。其中,單株莢數、單株粒數及單株粒重三者間相關系數均大于0.6,尤其是前兩者間的系數為0.9。莢數是控制大豆單株產量的主要因素。百粒重與莖粗為顯著正相關、與單株粒重為極顯著正相關,與蟲食粒數呈現顯著正相關、與蟲食粒率為極顯著正相關;4個抗蟲有關的性狀間彼此也表現為極顯著正相關;蟲食粒數和蟲食粒重同株高、節數、莢數、粒數和粒重間具有極顯著的正向關系,但前者僅與莖粗有顯著正相關性,后者同莖粗和分枝數分別為極顯著和顯著的正向關系;蟲食粒率則與分枝數、莖粗、莢數和粒數呈負相關性,其中前2個性狀關系顯著,后兩者關系極顯著。在選育大豆品種時應考慮到莢數、株高和百粒重的平衡,以保證在增產的同時減少蟲食損失。圖1
2.3 聚類分析與主成分
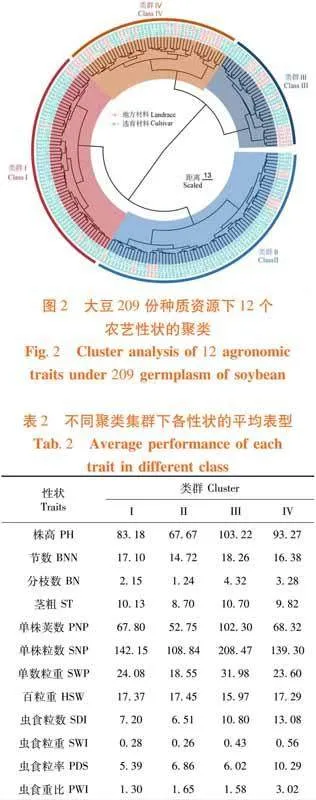
研究表明,地方材料與選育材料被分割為兩個部分,類群I和II主要由選育材料構成,類群III和IV容納了大多數的地方材料。各類群的具體特點如下,類群I包含70個材料,主要為黑龍江省和吉林省選育的品種,其特點為株高適中、節數較多,擁有較高的莢數和百粒重、分枝跨度大,剩余性狀均為中等水平;類群II有61份材料,以黑龍江省選育的品種為主。特征為植株較矮、節數和分枝數少,莢數少且單株粒重低;類群III有46份資源,地方材料與育成品種各半,來源及選育單位覆蓋遼寧、吉林和黑龍江三省,因而類群內部的節數、百粒重和籽粒抗蟲食性間差異較大。整體表現出高植株、單株莢數多且單株粒重大的特點;類群IV有32份種質,組成情況與類群III相似,而表現與類群I相近。該類種質的籽粒易受昆蟲取食,蟲食重比雖有較大的跨度但依舊高于其余類群。圖2,表2
所有性狀可劃分為4個主成分,累積貢獻率為83.47%,解釋了209份大豆資源中12類表型的大部分信息。除第4主成分的特征值為0.9,其余主成分均大于1。其中,第1主成分中作用最大3個性狀為單株莢數、粒數和粒重,特征值分別為0.89、0.86和0.75,反映了產量因素。同時,株高、分枝數、節數和莖粗的特征值也較高,體現了株型與產量因素的密切關系;第2主成分內蟲食粒率、蟲食重比、蟲食粒重和蟲食粒數的特征向量最大,分別為0.91、0.83、0.74和0.69,體現了植株的抗蟲食因素;第3主成分下,百粒重的特征值為0.89,其遠高于其他表型。也是產量因素之一,但側重于大豆籽粒的平均重量;第4主成分主要由株高和節數控制,特征值分別是0.47和0.39,可稱為株型因素。表3
2.4 分段線性回歸
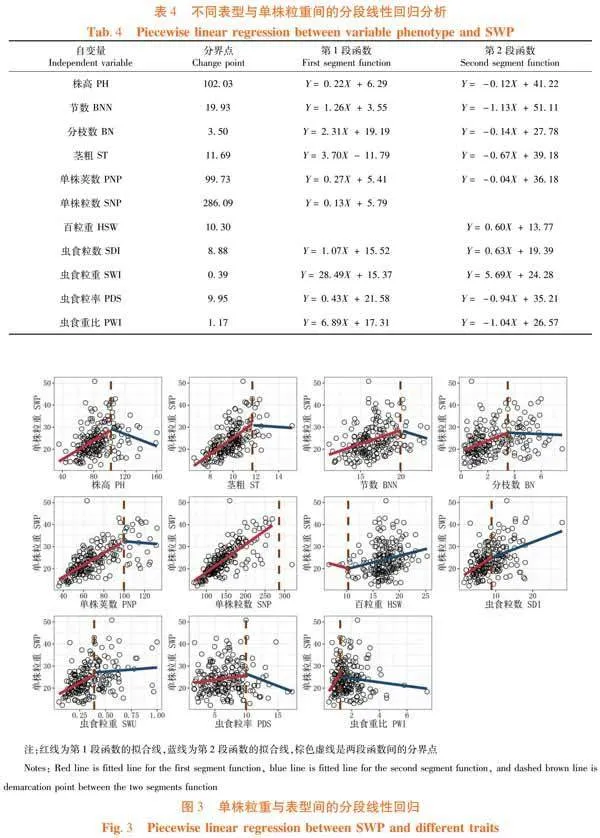
研究表明,單株粒重和百粒重由于在分界點前后所余數據量極少,分別無法獲得有效的第2段及第1段函數;株高、節數、分枝數和莖粗均經歷第1段隨數量增加而促進單株粒重提高的過程,同時分別在高102.03 cm、19.93個節、3.5個分枝及11.69 mm莖粗后導致粒重的下降;單株粒數和百粒重持續保持對粒重的正向貢獻。然而,單株莢數在超過99.73個后不利于粒重的增長,其可能同無效莢數的出現有關;蟲食粒數和蟲食粒重雖均保持與單株粒重的正向關系,但在8.88個和0.39 g后增幅放緩。蟲食粒率和蟲食重比則呈先增后減的趨勢,轉折點分別在9.95%和1.17%。使用本種質群體作為選育高產的材料時應注意株型和莢數適當,且不易受蟲害為害的資源。表4,圖3
2.5 種質資源綜合評價
研究表明,前3個主成分的累積貢獻率接近80%(79.5%),選擇前3個主成分作為重要主成分。隨后,株高(X1)、分枝數(X2)、節數(X3)、莖粗(X4)、單株莢數(X5)、單株粒數(X6)、單株粒重(X7)、百粒重(X8)、蟲食粒數(X9)、蟲食粒重(X10)、蟲食粒率(X11)和蟲食重比(X12)的標準化數據分別帶入下述公式計算F值:
F1 = -0.024 9X1 + 0.053 5X2 + 0.192 7X3 - 0.004 0X4 + 0.146 4X5 + 0.115 7X6 + 0.086 6X7 - 0.034 8X8 - 0.046 7X9 - 0.046 0X10 + 0.019 9X11 - 0.016 3X12.
F2 = -0.018 1X1 + 0.013 5X2 + 0.021 8X3 - 0.002 3X4 + 0.014 3X5 + 0.007 1X6 -0.027 0X7 - 0.034 9X8 + 0.098 7X9 + 0.085 8X10 + 0.162 8X11 + 0.115 8X12.
F3 = 0.043 3X1 + 0.023 0X2 - 0.100 4X3 + 0.011 2X4 + 0.040 1X5 + 0.058 8X6 + 0.076 7X7 + 0.012 9X8 - 0.009 9X9 - 0.000 8X10 + 0.014 7X11 + 0.024 7X12.
依照U值及DTOPSIS法計算公式對F值進行處理。已知F1至F3的貢獻率分別為40.73%、27.5%和11.27%,換算成權重為0.51、0.35和0.14。F2的主要部分是蟲食性狀,因此為權重添加方向。故綜合評分分值D = 0.51U1 - 0.35U2 + 0.14U3。209份大豆種質資源的綜合分數,范圍從0.59~-0.25,平均得分為0.16。其中,排名前50份種質資源幾乎全來自于III類,并且前10個有7份為地方材料。Soy194的綜合分數最高,表現為株型因素水平中等但分枝數多、有較多的莢數和粒數,單株粒重高(40.64 g,第7)且籽粒不易受昆蟲取食(蟲食粒率1.78%,蟲食重比0.58%)。綜合分數最高的選育材料是Soy052,其D值為0.5,排名第4。該種質為吉林省選育的品種,主要表現為極多的粒數(319.17個)、極小的百粒重(6.65 g)和好的抗蟲性(蟲食粒率1.70%,蟲食重比0.96%),其余性狀水平中等。排名第7的Soy196,是綜合分數第2的選育材料。作為吉林省選育品種,其特點是株型較大(株高、節數和莖粗均約為平均值的1.2倍),較多的莢數和粒數,單株粒重極高(42.56克,排名第2),但其抗蟲性一般(蟲食粒率5.24%,蟲食重比1.37%)。表5,表6
3 討 論
3.1
種質資源的性狀分析及表型評定是種質創新、品種改良的基礎[7]。對209份大豆種質的12個性狀分析的結果表明,目標群體內部具有較高的遺傳多樣性,適宜擇優作為育種材料并加以利用。同時,除節數、莖粗和百粒重外的性狀具有較高的變異系數,與前人對北方春大豆及東北種質群體的研究結果相符[3, 23]。分枝數的變異系數遠高于聶波濤等[3]的結果,可能是地方材料較多的緣故。對地方種質的表型調查可拓寬育種視野及可用資源[9]。蟲食粒數、蟲食粒重、蟲食粒率和蟲食重比在種質資源的變異系數均高于40%則顯示出東北春大豆在抗食心蟲上的能力差異,在其中挑選抗性品種將有助于減少田間損失及提高大豆產品品質,以利于產能的穩定增長。此外,上述4個性狀與其余性狀的相關性及分段線性回歸分析的結果顯示,蟲食粒數、蟲食粒重與單株莢數、粒數和粒重顯著正相關,蟲食粒率及蟲食重比與產量構成因素成反比或無顯著相關性。前2個指標易成為綜合評定中的干擾項目,而后2個指標適合作為評價抗蟲能力的標準。但是蟲食粒率常用于衡量大豆抗食心蟲能力[24, 25],而蟲食重比鮮見于相關研究。在今后的綜合評價體系及抗食心蟲研究中應將蟲食重比作為考察的指標之一。
3.2
前人對大豆性狀的相關性分析指出株型因子與產量因子常呈現正相關的特性[7, 13, 19],研究也發現株高、莖粗、分枝數和節數與單株莢數、粒數及粒重間具有極顯著的正相關性。然而,分段線性回歸分析的結果顯示出合理的株型及產量因子范圍更有益于單株粒重的提高。在研究所涉種質內,高產大豆理想數值為株高102.03 cm、19.93節、分枝3.5個、莖粗11.39 mm、豆莢99.73個、粒數286.09個,盡可能不受食心蟲的侵害。其結果與北方春大豆的研究結果相近[3]。此外,百粒重雖然正向調控單株粒重,但是其與株高、莢數和粒數等因子極顯著負相關。主成分分析將12個性狀濃縮為4個累積貢獻率大于80%的因素,其分別為產量、抗蟲、百粒重及株型因子,4個主成分可決定東北春大豆的多樣性。同時,在各主成分的特征向量中,百粒重也表現出與產量因子負相關,與抗蟲因子正相關的特點。因而,百粒重只需在適當范圍,可不作為高產大豆的主要選擇性狀。東北高產春大豆選育應優先選擇有效莢數和粒數較多、高抗食心蟲的中等株型材料。
通過聚類分析209份種質資源被分割為4個類群,與SSR標記聚類結果一致的是[26],4個類群中的依照選育品種及地方材料被分為2個大類。但具有絕大多數地方材料的類群,內部表現為地方材料和選育品種混合的特征,與北方春大豆的結果不符[3]。這些結果反映出所用種質不一而造成的特征,也可能是由于表型受遺傳及環境因素的共同調控致使結果不一的情況[7]。從類群的種質特點及性狀的平均表現上而言,第I類有中等株型及較多的莢數但單株產量不高,可與優異資源雜交以綜合株型及產量的優勢;第II類群體無株型和產量優勢,其原因可能為多隸屬黑龍江的早熟品種,在吉林省地區無法發揮其優勢,故不建議作為吉林省育種的主要材料;第III類群體具有高產特性,莢數和粒重高于其他類群。平均株高在103.22 cm,屬于理想類型。應結合抗蟲特點挑選優異種質作為主要的育種親本;第IV類的表現與第I類相似但抗蟲性不佳,可擇優選擇改造其抗蟲或產量特性。
DTOPSIS法被認為能夠較強地分辨種質資源的優劣而被用于大豆品種的表型綜合評價中[3, 27, 28]。研究利用攜帶性狀選育方向的無量綱方式及主成分賦權法計算了209份材料的D值。其中,排名第1的Soy194兼具株型和莢數的優勢且有較好的抗食心蟲特征,可作為骨干親本參與品種選育工作;排名第1的選育品種Soy052是多粒、高抗食心蟲的小粒品種,即可用于粒數和抗蟲的改良材料,也可作為特用納豆或芽豆的育種資源;排名第2的選育品種Soy196植株高大,單株產量排名第2但抗蟲性較差,若改善其株型及提高食心蟲抗性可獲得高產的大豆品種。另一方面,綜合評價排名靠前的材料均在III和I類群中,與高產類群重疊的情況證明了DTPOSIS法的合理性。同時,D值與株型因子和除百粒重外的產量因子極顯著正相關、與蟲食指數極顯著負相關的結果也進一步證實了DTOPSIS法可容納研究對象的重要指標以用于分辨材料間的優劣程度。此外,綜合評價指數前5%的材料中(共11份),有8份屬于地方材料,而3份選育材料中有2份為吉林省的品種,在大豆品種選育中要注意結合地方材料及種質區域的優良材料以針對性地培育高產穩產的優異品種。
4 結 論
209份東北春大豆連續3年的12個性狀中除節數、莖粗和百粒重外的性狀變異系數較高,可結合育種目標擇優利用;產量、抗蟲、百粒重及株型因子可用于概括東北春大豆的表型特點,其中產量因子與株型正相關,與百粒重、蟲食粒數和蟲食重比負相關;以單株粒重為因變量的分段線性回歸指出略高于群體平均值的株型水平、多粒數及優異的大豆食心蟲抗性是高產大豆的主要特征;聚類分析顯示I類群、III類群可分別用于株型優化、高產育種中,IV類群在使用時需要考慮其對食心蟲的抗性問題,II類群不適宜作為吉林省中部產區的育種資源;DTPOSIS法對大豆種質資源的綜合評價具有較好的效果,其排名突出的Soy194、Soy052和Soy196可分別作為骨干親本、粒數改良和粒重改良材料。
參考文獻(References)
[1]查霆, 鐘宣伯, 周啟政, 等. 我國大豆產業發展現狀及振興策略[J]. 大豆科學, 2018, 37(3): 458-463.
ZHA Ting, ZHONG Xuanbo, ZHOU Qizheng, et al. Development status of China’s soybean industry and strategies of revitalizing[J]. Soybean Science, 2018, 37(3): 458-463.
[2] 徐瑤, 冷蘇鳳, 張玉明, 等. 1982-2021年江蘇省審定大豆品種主要農藝性狀、產量、品質及抗性演變分析[J]. 中國油料作物學報, 2022, 44(4): 780-789.
XU Yao, LENG Sufeng, ZHANG Yuming, et al. Evolution analysis of main agronomic traits, yield, quality and resistance of soybean varieties released in Jiangsu Province from 1982 to 2021[J]. Chinese Journal of Oil Crop Sciences, 2022, 44(4): 780-789.
[3] 聶波濤, 劉德泉, 陳健, 等. 北方春大豆品種農藝和品質性狀分析與綜合評價[J]. 作物學報, 2024, 50(9): 2248-2266.
NIE Botao, LIU Dequan, CHEN Jian, et al. Analysis and comprehensive evaluation of agronomic and quality traits of spring soybean varieties in Northern China[J]. Acta Agronomica Sinica, 2024, 50(9): 2248-2266.
[4] 邱麗娟, 李英慧, 關榮霞, 等. 大豆核心種質和微核心種質的構建、驗證與研究進展[J]. 作物學報, 2009, 35(4): 571-579.
QIU Lijuan, LI Yinghui, GUAN Rongxia, et al. Establishment, representative testing and research progress of soybean core collection and mini core collection[J]. Acta Agronomica Sinica, 2009, 35(4): 571-579.
[5] 常汝鎮, 孫建英, 陳一舞. 中國大豆種質創新的內容與方法[J]. 作物品種資源, 1995, (1): 2-4.
CHANG Ruzhen, SUN Jianying, CHEN Yiwu. Content and Methods of Soybean Germplasm Innovation in China[J]. China Seed Industry, 1995, (1): 2-4.
[6] 趙朝森, 王瑞珍, 李英慧, 等. 江西大豆種質資源表型及品質性狀綜合分析與評價[J]. 大豆科學, 2019, 38(5): 686-693.
ZHAO Chaosen, WANG Ruizhen, LI Yinghui, et al. Comprehensive analysis and evaluation of the phenotype and quality traits of Jiangxi soybean germplasm resources[J]. Soybean Science, 2019, 38(5): 686-693.
[7] 徐澤俊, 齊玉軍, 邢興華, 等. 黃淮海大豆種質農藝與品質性狀分析及綜合評價[J]. 植物遺傳資源學報, 2022,23(2): 468-480.
XU Zejun, QI Yujun, XING Xinghua, et al. Analysis and evaluation of agronomic and quality traits in soybean germplasms from Huang-Huai-Hai Region[J]. Journal of Plant Genetic Resources, 2022,23(2): 468-480.
[8] 曾維英, 賴振光, 譚玉榮, 等. 廣西地方大豆種質資源的收集與評價鑒定篩選[J]. 大豆科學, 2021, 40(3): 354-361.
ZENG Weiying, LAI Zhenguang, TAN Yurong, et al. Collection, evaluation and screening of local soybean germplasm in Guangxi[J]. Soybean Science, 2021, 40(3): 354-361.
[9] 蓋鈞鎰, 崔章林. 我國南方大豆地方品種群體特點和特異種質的發掘與遺傳基礎研究[J]. 中國農學通報, 1993, 9(2): 1-5.
GAI Junyi, CUI Zhanglin. Studies on the properties of soybean Landrace population from Southern China and on the germpiasm with specific target traits and their genetic background[J]. Chinese Agricultural Science Bulletin, 1993, 9(2): 1-5.
[10] 常汝鎮, 孫建英, 邱麗娟. 中國大豆種質資源研究進展[J]. 作物雜志, 1998, (3): 7-9.
CHANG Ruzhen, SUN Jianying, QIU Lijuan. Research progress on soybean germplasm resources in China[J]. Crops, 1998,(3): 7-9.
[11] 王繼亮, 宗春美, 王德亮, 等. 東北大豆種質群體在佳木斯的表型鑒定及利用探析[J]. 中國油料作物學報, 2021, 43(6): 996-1005.
WANG Jiliang, ZONG Chunmei, WANG Deliang, et al. Identification, evaluation and improvement utilization of Northeast China Soybean Germplasm Population in Jiamusi[J]. Chinese Journal of Oil Crop Sciences, 2021, 43(6): 996-1005.
[12] Zhao G Y, Jiang Z F, Li D M, et al. Molecular loci associated with seed isoflavone content may underlie resistance to soybean pod borer (Leguminivora glycinivorella)[J]. Plant Breeding, 2015, 134(1): 78-84.
[13] 孟珊, 徐婷婷, 朱小品, 等. 江蘇大豆地方種質資源表型多樣性分析[J]. 植物遺傳資源學報, 2023,24(2): 419-436.
MENG Shan, XU Tingting, ZHU Xiaopin, et al. Diversity analysis of soybean landraces collected from Jiangsu Province using phenotypic traits[J]. Journal of Plant Genetic Resources, 2023,24(2): 419-436.
[14] 李艷花, 杜成章, 陳紅, 等. 重慶大豆地方資源多樣性評價及群體表型特點研究[J]. 植物遺傳資源學報, 2013, 14(6): 1025-1030.
LI Yanhua, DU Chengzhang, CHEN Hong, et al. Study of genetic diversity and population morphological characteristics of soybean landraces in Chongqing[J]. Journal of Plant Genetic Resources, 2013, 14(6): 1025-1030.
[15] 齊建雙, 夏來坤, 黃保, 等. 基于熵權的DTOPSIS法和灰色局勢決策法在玉米品種區域試驗中的應用探討[J]. 作物雜志, 2021, (1): 60-67.
QI Jianshuang, XIA Laikun, HUANG Bao, et al. Discussion on the application in the regional experiment of maize varieties by entropy DTOPSIS mode and grey situation decision methods[J]. Crops, 2021, (1): 60-67.
[16] 宋慧, 郭巖, 邢璐, 等. 基于灰色關聯度、DTOPSIS與灰色局勢決策法的谷子品種綜合評價[J]. 中國農業大學學報, 2023,28(11): 42-56.
SONG Hui, GUO Yan, XING Lu, et al. Comprehensive evaluation of foxtail millet varieties based on grey correlation degree, DTOPSIS and situational decision-making method[J]. Journal of China Agricultural University, 2023,28(11): 42-56.
[17] 高山, 閆程銘, 萬暢, 等. 基于灰色關聯度法和DTOPSIS法對羊草種質資源綜合評價[J]. 草地學報, 2024, 32(2): 599-609.
GAO Shan, YAN Chengming, WAN Chang, et al. Comprehensive evaluation of germplasm resources of Leymus chinensis based on grey correlation method and DTOPSIS method[J]. Acta Agrestia Sinica, 2024, 32(2): 599-609.
[18] 段會軍, 張彩英, 王省芬, 等. 河北省大豆品種主要農藝性狀及聚類分析[J]. 河北農業大學學報, 2003,26(2): 5-9.
DUAN Huijun, ZHANG Caiying, WANG Xingfen, et al. The main agronomic traits and cluster analysis of soybean varieties in Hebei[J]. Journal of Agricultural University of Hebei, 2003,26(2): 5-9.
[19] 黎松松, 賴建軍, 張紅梅, 等. 江蘇鮮食春大豆種質資源表型鑒定及綜合評價[J]. 大豆科學, 2022, 41(4): 385-396.
LI Songsong, LAI Jianjun, ZHANG Hongmei, et al. Phenotyping identification and comprehensive evaluation of fresh spring soybean germplasms in Jiangsu Province[J]. Soybean Science, 2022, 41(4): 385-396.
[20] Charrad M, Ghazzali N, Boiteau V, et al. NbClust: AnRPackage for determining the relevant number of clusters in a data set[J]. Journal of Statistical Software, 2014, 61(6): 1-36.
[21] Shannon C E, Weaver W. The mathematical theory of communication[M]. Champaign, IL, US: University of Illinois Press, 1949: 117.
[22] 孫東雷, 卞能飛, 陳志德, 等. 花生種質資源表型性狀的綜合評價及指標篩選[J]. 植物遺傳資源學報, 2018, 19(5): 865-874.
SUN Donglei, BIAN Nengfei, CHEN Zhide, et al. Comprehensive evaluation and index screening of phenotypic traits in peanut germplasm resources[J]. Journal of Plant Genetic Resources, 2018, 19(5): 865-874.
[23] 蘇江順, 齊玉鑫, 楊君, 等. 東北大豆種質群體在吉林省白城市的表現及其潛在的育種意義[J]. 土壤與作物, 2019, 8(1): 1-10.
SU Jiangshun, QI Yuxin, YANG Jun, et al. Performance and genetic potential of Northeast China soybean germplasm population in Baicheng City of Jilin Province[J]. Soils and Crops, 2019, 8(1): 1-10.
[24] 張君, 蔡立楠, 姚丹, 等. 大豆抗食心蟲主基因+多基因遺傳效應分析[J]. 中國農學通報, 2012,28(36): 227-230.
ZHANG Jun, CAI Linan, YAO Dan, et al. Analysis of major gene plus polygene inheritance effects on soybean pod borer resistance[J]. Chinese Agricultural Science Bulletin, 2012,28(36): 227-230.
[25] 崔娟, 喬方, 秦貝華, 等. 大豆食心蟲成蟲對大豆品種的產卵選擇性和幼蟲適生性分析[J]. 中國油料作物學報, 2023, 45(3): 592-599.
CUI Juan, QIAO Fang, QIN Beihua, et al. Oviposition selectivity and larval fitness of soybean pod borer, Leguminivora glycinivorella(Lepidoptera: Olethreutidae)to different soybean varieties[J]. Chinese Journal of Oil Crop Sciences, 2023, 45(3): 592-599.
[26] 林凡云, 邱麗娟, 常汝鎮, 等. 山西省大豆地方品種與選育品種農藝性狀及SSR標記遺傳多樣性比較分析[J]. 中國油料作物學報, 2003,25(3): 26-31.
LIN Fanyun, QIU Lijuan, CHANG Ruzhen, et al. Genetic diversity of Landrace and bred varieties of soybean in Shanxi[J]. Chinese Journal of Oil Crop Scieves, 2003,25(3): 26-31.
[27] 昝凱, 陳亞光, 申為民, 等. 黃淮海夏大豆(南片)品種(系)農藝性狀的綜合分析及評價模型構建[J]. 大豆科學, 2023, 42(2): 129-137.
ZAN Kai, CHEN Yaguang, SHEN Weimin, et al. Comprehensive analysis of agronomic characters of summer sowing soybean varieties(lines) in Huang-Huai-Hai Region(south) and constructing A comprehensive evaluation model[J]. Soybean Science, 2023, 42(2): 129-137.
[28] 昝凱, 周青, 張志民, 等. 灰色關聯度和DTOPSIS法綜合分析河南區域試驗中大豆新品種(系)的農藝性狀表現[J]. 大豆科學, 2018, 37(5): 664-671.
ZAN Kai, ZHOU Qing, ZHANG Zhimin, et al. Gray correlation analysis and DTOPSIS method for comprehensive agronomic performance analysis of new soybean varieties(lines) in Henan regional test[J]. Soybean Science, 2018, 37(5): 664-671.
Phenotype analysis and comprehensive evaluation of spring soybean germplasm resources from northeast China
YANG Xiangbo1, CHEN Liangyu2, YANG Songnan2, CHEN Xifeng2, XING Weiming3, LI Xueying2, CONG Weixuan2, ZANG Zhenyuan2, ZANG Yuanbo4, ZHANG Jun2
(1. College of Agriculture, Jilin Agricultural Science and Technology University, Jilin Jilin 132101, China; 2. College of Agronomy, Jilin Agricultural University, Changchun 130118, China; 3. Agricultural and Technology Extension Station of Jilin Province, Changchun 130033, China; 4. Comprehensive Office of Yongping Township Government in Jilin Province, Fuyu Jilin 131200, China)
Abstract:【Objective】 To enhance the utilization efficiency of spring soybean germplasm resources in northeast China and identify high-quality materials.
【Methods】 A total of 209 cultivars and landraces underwent a series of statistical analyses, including descriptive statistics, cluster analysis, segmented linear regression analysis, and a comprehensive evaluation of 12 traits over a three-year period.
【Results】 The range of phenotypic coefficients of variation and diversity indices indicated that branch node number, stem thickness, and one hundred seed weight showed a relatively stable pattern.In contrast, nine traits, including number of branches, percentage of damaged seeds weight by insect of total seed weight per plant, and pod number per plant, exhibited a high degree of variability.Principal component analysis revealed that all traits could be condensed into yield, insect resistance, one hundred seed weight, and plant architecture factors.The yield factor was positively correlated with plant architecture factor and negatively correlated with one hundred seed weight, percentage of damaged seeds by insect of total seed number per plant, and percentage of damaged seeds weight by insect of total seed weight per plant.Germplasm resources could be classified into four categories.Class I was characterized by moderate plant architecture and high pod number.Class II was distinguished by short plants and low yield per plant.Class III was defined by ideal plant architecture and the highest yield.Class IV was similar to Group I but exhibited poor insect resistance.The results of piecewise linear regression analysis indicated that the optimal high-yielding soybean characteristics exhibited a plant height of approximately 102.03 cm, 19.93 nodes, 3.5 branches, 11.39 mm stem thickness, 99.73 pods, and 286.09 seeds, with minimal feeding by soybean pod borer.The DTOPSIS (dynamic technique for order preference by similarity to ideal solution) method calculated D scores of the germplasm ranging" from 0.59 to -0.25, with a mean of 0.16.
【Conclusion】 The phenotypic diversity of spring soybeans in northeast China is considerable, with Class I and Class III representing promising sources for plant architecture improvement and high-yield breeding, respectively.High-yielding soybeans are distinguished by their moderate plant architecture, high seed number, and insect resistance.The three resources Soy194, Soy052, and Soy196, which have achieved excellent D scores, are exemplary germplasm for soybean breeding.
Key words:soybean; agronomic traits; cluster analysis; piecewise liner regression; comprehensive evaluation
Fund projects:Jilin Science and Technology Development Program Project \"Breeding of High-yielding and High-quality New Cultivar of Soybean and Application of Gene Editing Technology to Germplasm Creation\" (20240303009NC)
Correspondence author:ZHANG Jun (1968 -), male, from Shuangliao,Jilin,professor, doctor,research direction: crop genetics and breeding, (E-mail) zhangjun@jlau.edu.cn
基金項目:吉林省科技發展計劃項目“大豆高產、優質新品種選育及基因編輯技術應用于種質創制研究”(20240303009NC)
作者簡介:楊祥波(1980-),男,吉林人,副教授,博士,研究方向為作物遺傳育種,(E-mail)yangxiangbo1980@163.com
通訊作者:張君(1968-),男,吉林雙遼人,教授,博士,研究方向為作物遺傳育種,(E-mail)zhangjun@jlau.edu.cn
猜你喜歡
今日農業(2022年16期)2022-11-09 23:18:44
中國化肥信息(2022年5期)2022-08-30 01:58:26
今日農業(2021年20期)2021-11-26 01:23:56
今日農業(2021年14期)2021-10-14 08:35:34
下一代英才(酷炫少年)(2018年6期)2018-07-09 03:17:44
經濟研究導刊(2016年28期)2016-12-14 09:22:57
科學與財富(2016年26期)2016-12-01 21:32:23
大學教育(2016年11期)2016-11-16 20:33:18
企業導報(2016年9期)2016-05-26 20:52:48
紅領巾·萌芽(2006年3期)2006-01-01 00:00:00
