西南樺節間莖段植株再生體系的建立
2024-01-20 09:47:58郭俊杰王春勝尹海鋒
林業科學研究 2023年6期
王 歡,郭俊杰,王春勝,尹海鋒,曾 杰
(1.中國林業科學研究院熱帶林業研究所,熱帶林業研究國家林草局重點實驗室 廣東 廣州 510520;2.南京林業大學,江蘇 南京 210037)
西南樺(Betula alnoidesBuch.-Ham.ex D.Don)是中國南方造林面積較大的鄉土珍貴闊葉樹種之一,其木材材質優良,是制作木地板、高檔家具、樂器等的優良材料[1];其樹皮可用于治療感冒、產后疼痛以及消化系統和骨科疾病等[2],樹皮提取物具有抗HIV-1 整合酶,并具有抗炎、降血脂、減肥之功效[3-5]。西南樺在生物多樣性和地力維持、水源涵養以及碳素固定等方面亦發揮著重要作用,被廣泛應用于生態公益林和珍貴用材林建設。隨著分子生物技術的快速發展,利用基因工程進行遺傳改良已成為林木育種的重要途徑之一。遺傳轉化是開展植物遺傳改良的必要技術,同時也是驗證基因功能、解析性狀形成機制的關鍵技術,而高效穩定的轉化受體再生體系是遺傳轉化的必要前提。
中國西南樺的離體培養始于2000 年[6],主要有直接器官發生[7-9]、間接器官發生[10]以及體胚發生[6]等構建方式。目前能形成完整植株的主要是通過直接器官發生途徑,特別是腋芽萌發途徑,但該途徑增殖率通常較低,且只適合于西南樺優良品種的規模擴繁,并不適用于遺傳轉化。間接器官發生是離體培養中通過外植體誘導愈傷組織再分化的形態建成過程[11],相比直接器官發生,其潛在繁殖系數更高,作為分子育種的平臺也更具優勢[12-13]。目前,西南樺通過間接器官發生方式的再生體系尚未見成功報道。
關于樺木科的間接器官發生,目前以白樺(Betula platyphyllaSuk)和歐洲白樺(Betula pendulaRoth)研究較多,所用外植體一般為種子[14-16]、葉片[17-19]、根段[20]、莖段[21]等。以種子為外植體研發的再生體系,具有分化率高且再生芽多的優勢,但是,由于該體系需要“切種子”,對于細小粒的樺木種子而言技術要求極高,且每粒種子的遺傳背景不同,嚴格地說,它并不利于開展轉基因植株的分子試驗。利用無性系組培苗的根、莖、葉作為外植體,可使受體材料保持相對一致。有研究發現,利用白樺莖段、葉柄和葉片均能誘導愈傷組織和不定芽分化,莖段的效果整體上優于葉片和葉柄[22];歐洲白樺根較難誘導愈傷組織形成,且其愈傷組織易老化,需要較長時間誘導才能分化出不定芽[20]。前期研究發現,西南樺莖段與白樺[22]一樣較易誘導愈傷組織和不定芽分化,但因節間較短容易切到芽點,增加了操作難度,且材料利用率不高。因此,增加西南樺試管苗的節間長度對莖段再生的外植體取材非常重要。
本研究以西南樺無性系TC2 試管苗為材料,針對愈傷組織誘導、不定芽分化、生根等器官間接發生階段開展試驗,重點研究激素濃度、暗培養時間和莖段外植體培育條件等對不定芽誘導的影響,以期建立高效穩定、重復性良好的西南樺間接器官發生再生體系,為今后開展西南樺組織培養以及利用基因工程進行其遺傳性狀改良奠定基礎。
1 材料與方法
1.1 試驗材料及條件
以課題組前期培養的西南樺無性系TC2 試管苗為材料;如無特別注釋,其培養條件為光照強度2 000 lx,光照時間16 h·d-1,溫度25 ℃ ± 2 ℃。
1.2 研究方法
1.2.1 莖段再生階段培養基的篩選 莖段再生階段培養基的篩選試驗中,選取繼代培養30 d 的TC2 試管苗,剪取其帶頂芽莖段,接種于培養基WPM + 0.15 mg·L-1NAA + 20 g·L-1蔗糖 + 5.8 g·L-1瓊脂(pH5.8),培養30 d 后避開腋芽剪取4~7 mm 節間莖段用于愈傷組織誘導試驗。
(1)愈傷組織誘導階段 愈傷組織誘導研究包括基本培養基篩選、外源激素的濃度篩選以及暗培養時間篩選3 個單因素試驗(表1),分別含4、9 和3 個處理(水平)。所有試驗每個處理均3 次重復,每次10 個莖段。
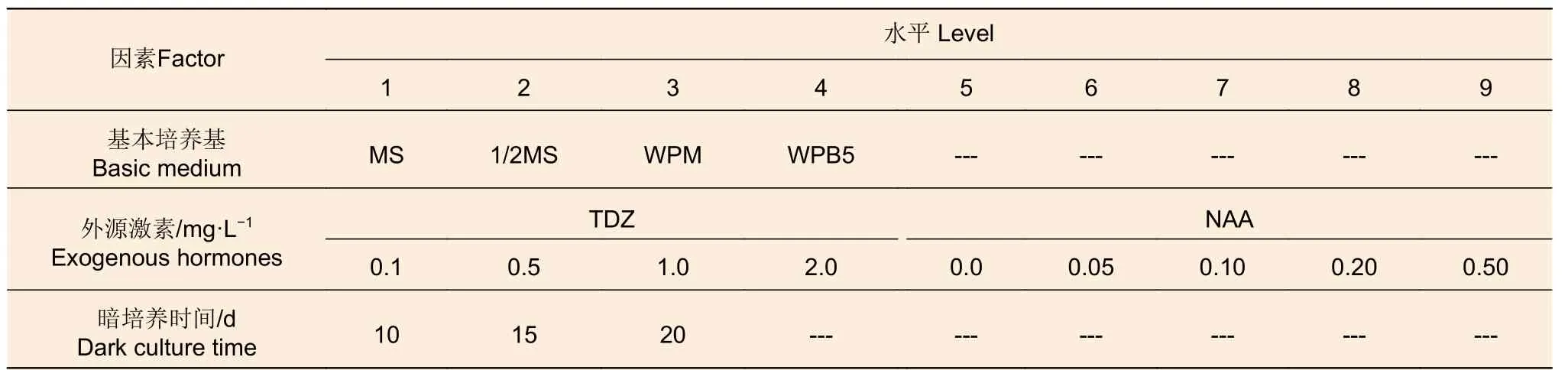
表1 愈傷組織誘導階段的單因素試驗Table 1 Single-factor experiments at callus induction stage
基本培養基試驗中,各處理的激素添加均為1.0 mg·L-1TDZ 和0.1 mg·L-1NAA;外源激素濃度梯度試驗以WPB5 為基本培養基,選用TDZ 和NAA 的不同濃度(表1)進行試驗;暗培養時間優化試驗按表1 中的3 個時長進行培養,所用培養基為WPB5 + 1 mg·L-1TDZ + 0.1 mg·L-1NAA +20 g·L-1蔗糖 + 5.8 g·L-1瓊脂 (pH5.8)。統計愈傷誘導率后將表1 中所有試驗處理的愈傷組織轉接至培養基WPM + 1 mg·L-16-BA + 0.5 mg·L-1GA3+30 g·L-1蔗糖 + 5.8 g·L-1瓊脂 (pH5.8)中進行后續分化培養,在正常光照條件下培養60 d 后統計發生分化的愈傷組織數與每個莖段分化的不定芽數(高度5 mm 以上),期間30 d 繼代1 次,并計算分化率、凈增殖系數和總增殖系數。
(2)愈傷組織分化不定芽階段 以WPM 為基本培養基,采用6-BA(A)、GA3(B)和NAA(C)3 種激素(3 因素),每因素包括2 水平,構成完全隨機試驗(表2),分析各激素組合的不定芽誘導效果,以篩選分化培養基。選擇在上述最佳愈傷組織誘導培養基上培養出的大小相近的愈傷組織,分別接種于8 種組合處理,每個處理3 次重復,每次重復接種10 塊愈傷組織。每隔30 d 轉接(繼代)1 次,60 d 后觀察統計不定芽誘導情況,包括發生分化的愈傷組織數、每個莖段分化的不定芽數、分化率、凈增殖系數和總增殖系數。
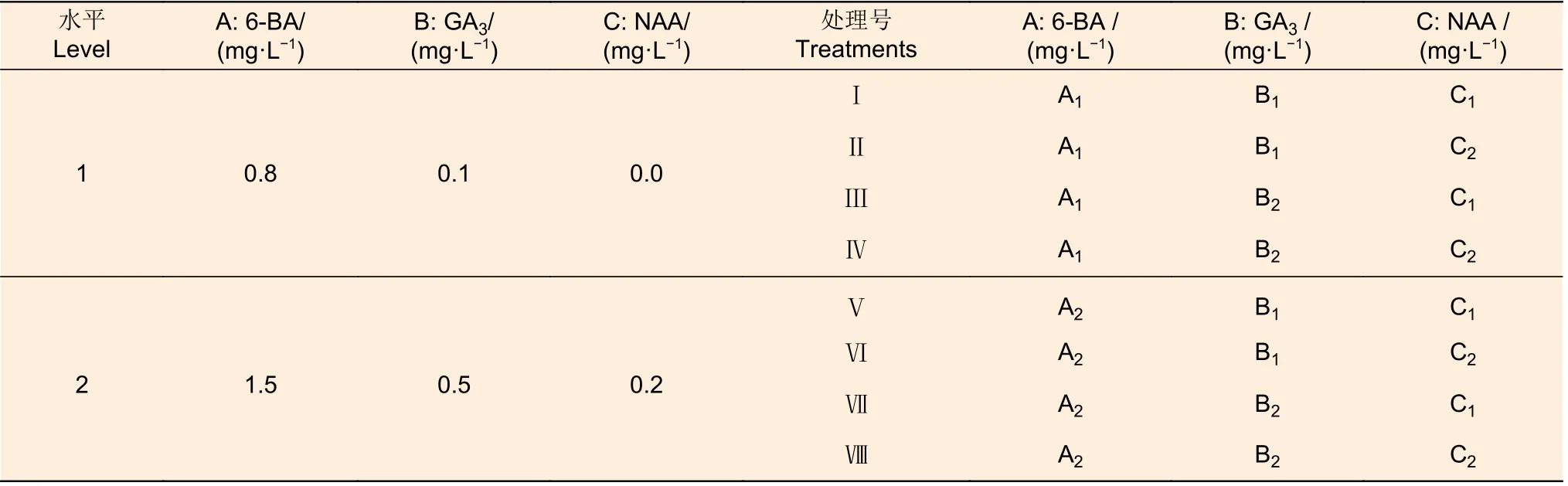
表2 愈傷組織分化階段激素組合試驗Table 2 Hormone combination experiment at callus differentiation stage
(3) 生根培養階段 待分化的不定芽長至2~3 cm,剪取單芽接種至1/2MS 或WPM 培養基,附加NAA(0.1 和0.2 mg·L-1)或IBA (0.1 和0.2 mg·L-1)、20 g·L-1蔗糖和 5.8 g·L-1瓊脂進行生根誘導,以篩選生根培養基。試驗設計詳見表3。每個處理3 次重復,每次重復12 株。培養過程中追蹤統計各組合生根起始時間,于12 d 和30 d 后統計生根率和生根條數,并測定30 d 時根長。
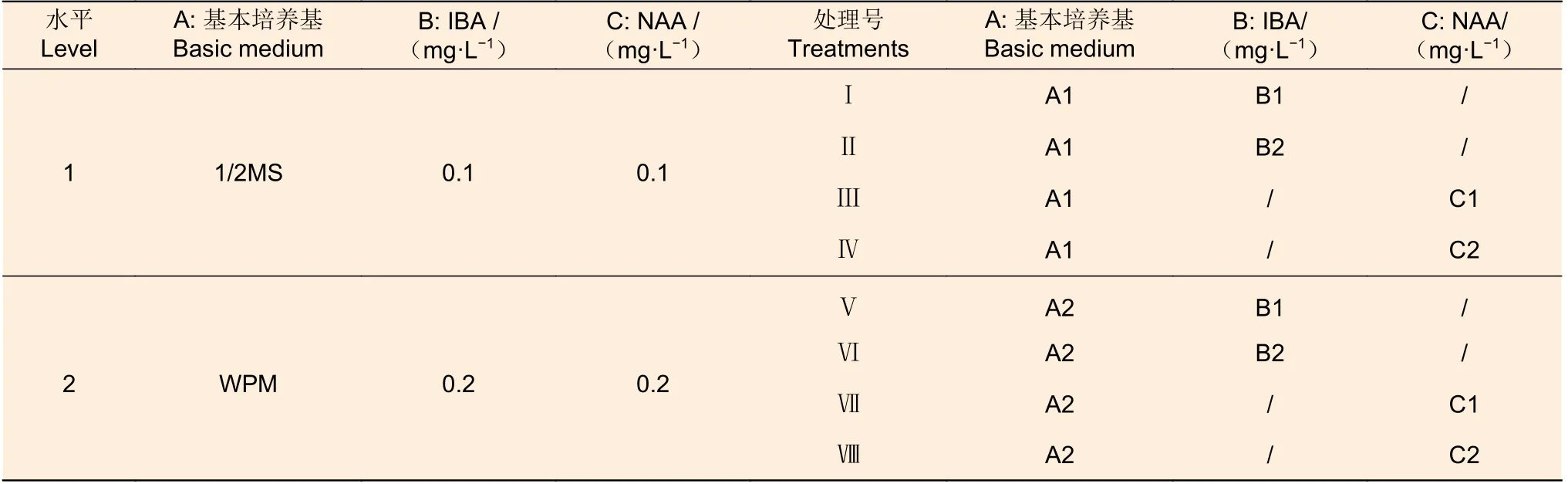
表3 生根誘導試驗設計Table 3 Experimental design for rooting induction
1.2.2 預培養條件的優化 因研究過程中發現外植體的狀態影響各階段的試驗效果,于是針對預培養條件進行如下優化。選取繼代培養30 d 的TC2 試管苗,剪取其帶頂芽莖段,接種于培養基WPM + 0.15 mg·L-1NAA + 20 g·L-1蔗糖 + 5.8 g·L-1瓊脂(pH5.8),于8 種條件下進行培育(表4)。每個處理3次重復,每次重復10 株,30 d 后統計株高和節間長度。分別取8 個處理的節間莖段,同時統計每株可取平均莖段數,然后接種于最佳愈傷組織誘導培養基(WPB5 + 1.0 mg·L-1TDZ + 0.2 mg·L-1NAA + 20 g·L-1蔗糖 +5.8 g·L-1瓊脂 );暗培養15 d 后轉至最佳愈傷組織分化的培養基(WPM + 0.8 mg·L-16-BA + 0.5 mg·L-1GA3+ 30 g·L-1蔗糖 + 5.8 g·L-1瓊脂)。每個處理3次重復, 每次重復24 個莖段,75 d 后統計不定芽分化情況,統計指標同(2)。

表4 預培養條件篩選試驗Table 4 Screening test on preculture conditions of internode
1.3 數據分析
統計各試驗數據,計算以下指標:
Y=(n/N) × 100%,F=(m/M) × 100%,R=(q/Q) × 100%,Tt=Na/m,Tn=Na/M
Y為愈傷誘導率,n為愈傷組織誘導數,N為接種莖段數;F為愈傷分化率,m為分化愈傷組織數,M為接種愈傷組織數;R為生根率,q為生根株數,Q為接種數;Tt為總增殖系數,Tn為凈增殖系數,Na為不定芽總數。使用Microsoft Excel 2010 軟件進行數據整理,使用IBM SPSS Statistics13.0 軟件進行方差分析和Duncan 多重比較。
2 結果與分析
2.1 愈傷組織誘導階段各因素對后續不定芽誘導和增殖的影響
2.1.1 基本培養基 方差分析結果(表5)表明:除凈增殖系數外,4 種基本培養基處理間產生的不定芽數(10 個接種莖段不定芽數量)及其分化率和總增殖系數均呈現顯著差異(P<0.05),其中,WPB5 的所有指標均最高,WPM、1/2MS和MS 間不定芽數、分化率和總增殖系數依次顯著地降低,凈增殖系數差異不顯著,說明采用WPB5作為基本培養基,可顯著地促進西南樺莖段再生不定芽的形成,提高其增殖系數,因此,WPB5 是西南樺莖段再生不定芽培養的優選培養基。
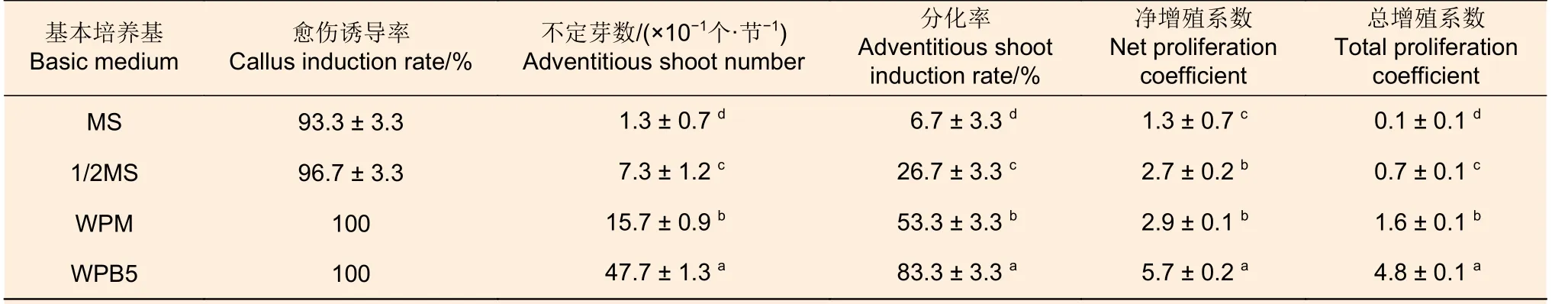
表5 愈傷組織誘導階段基本培養基對后續西南樺不定芽分化的影響Table 5 Effect of basic medium at callus induction stage on subsequent adventitious bud differentiation of Betula alnoides
2.1.2 外源激素 由表6 可知:隨著外源激素TDZ 或NAA 濃度的增加,不定芽數、分化率和總增殖系數均呈現先增加后下降的趨勢,凈增殖系數在兩種激素中表現不同,隨著TDZ 濃度增加呈遞增趨勢,而隨NAA 濃度增加則呈現先增加后下降的趨勢; TDZ 或NAA 不同濃度處理間4 個指標均存在顯著差異。TDZ 的4 個濃度處理中,不定芽數、分化率和總增殖系數以1 mg·L-1處理為最高,分別為49、83.3%和4.9,顯著地高于其它處理;凈增殖系數最高的TDZ 濃度為2 mg·L-1,但與1 mg·L-1處理差異不顯著。NAA 的5 個濃度處理中, 0.2 mg·L-1處理的不定芽數、分化率和總增殖系數均最高,分別為48.3、86.7%、4.8;凈增殖系數最高的NAA 濃度為0.1 mg·L-1;除分化率在0.1 mg·L-1和0.2 mg·L-1處理間差異顯著之外,其余指標在二者間均差異不顯著。總的來看,TDZ 或NAA 為西南樺莖段愈傷組織誘導的關鍵外源激素,其適宜濃度分別為1 mg·L-1和0.2 mg·L-1。
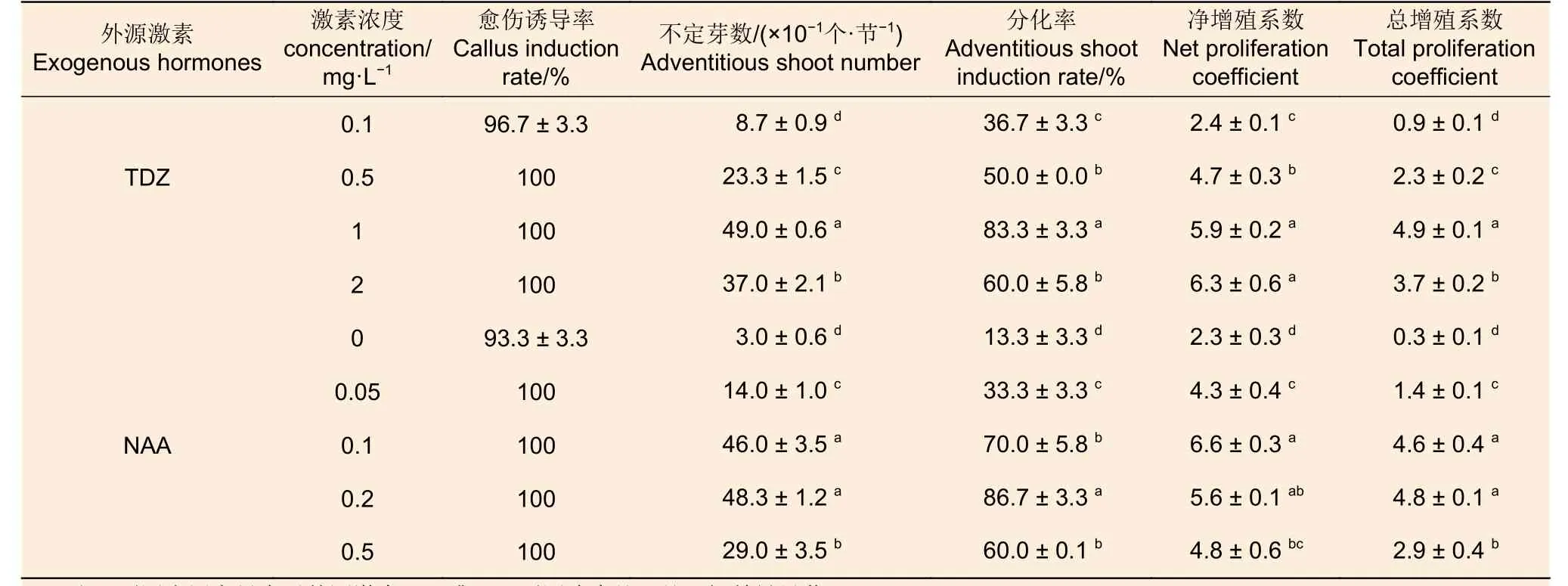
表6 愈傷組織誘導階段外源激素及其濃度對后續西南樺不定芽分化的影響Table 6 Effects of exogenous hormones and their concentrations at callus induction stage on subsequent adventitious bud differentiation of Betula alnoides
2.1.3 暗培養時間 在暗培養時間篩選試驗中,不定芽數、分化率、凈增殖系數和總增殖系數隨暗培養時間增加整體上呈現先增加后下降的趨勢(表7)。暗培養15 d 和20 d 時4 個指標均顯著高于10 d,且以15 d 暗培養處理效果最好;兩者的分化率均在80.00%以上,凈增殖系數和總增殖系數分別在5.5 和4.5 以上。由此可見,西南樺莖段愈傷組織誘導的適宜暗培養時間為15 d。
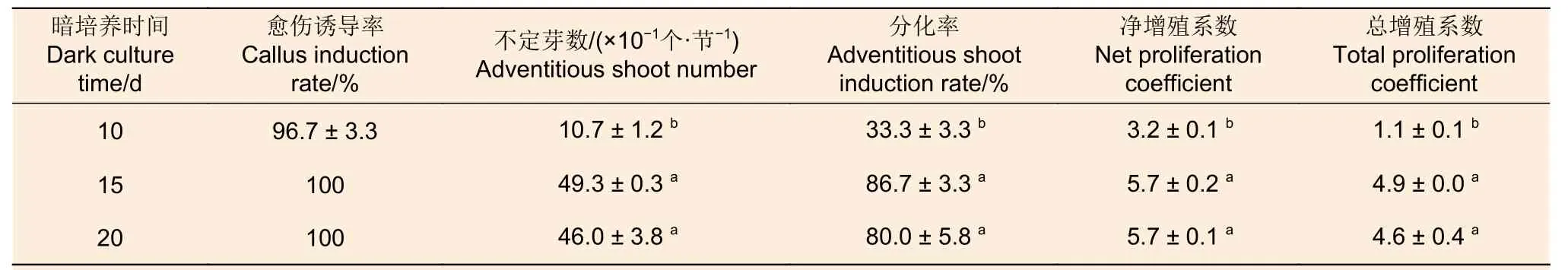
表7 愈傷組織誘導階段暗培養時間對后續西南樺不定芽分化的影響Table 7 Effect of dark culture time at callus induction stage on subsequent adventitious buds differentiation of Betula alnoides
2.2 愈傷組織分化階段不同激素組合對不定芽誘導和增殖的影響
GA3和NAA 兩種因素各水平間不定芽數、不定芽分化率和總增殖系數存在顯著差異,其凈增殖系數無顯著差異,6-BA 各水平間這些指標差異不顯著;3 種因素間交互作用不顯著(表8)。從表9可以看出:各激素組合處理的不定芽數、不定芽分化率、凈增殖系數和總增殖系數分別為38.7~49.3、63.3%~86.7%、5.4 ~6.1 倍和3.9 ~4.9倍;組合處理間不定芽數、不定芽分化率、總增殖系數和凈增殖系數均存在顯著差異(P<0.05)。組合處理間比較,處理III 的不定芽數、不定芽分化率和總增殖系數最高,其凈增殖系數也相對較高。對照表2 可知:0.8 mg·L-16-BA 和0.5 mg·L-1GA3的組合適宜應用于西南樺愈傷組織的不定芽分化和增殖。
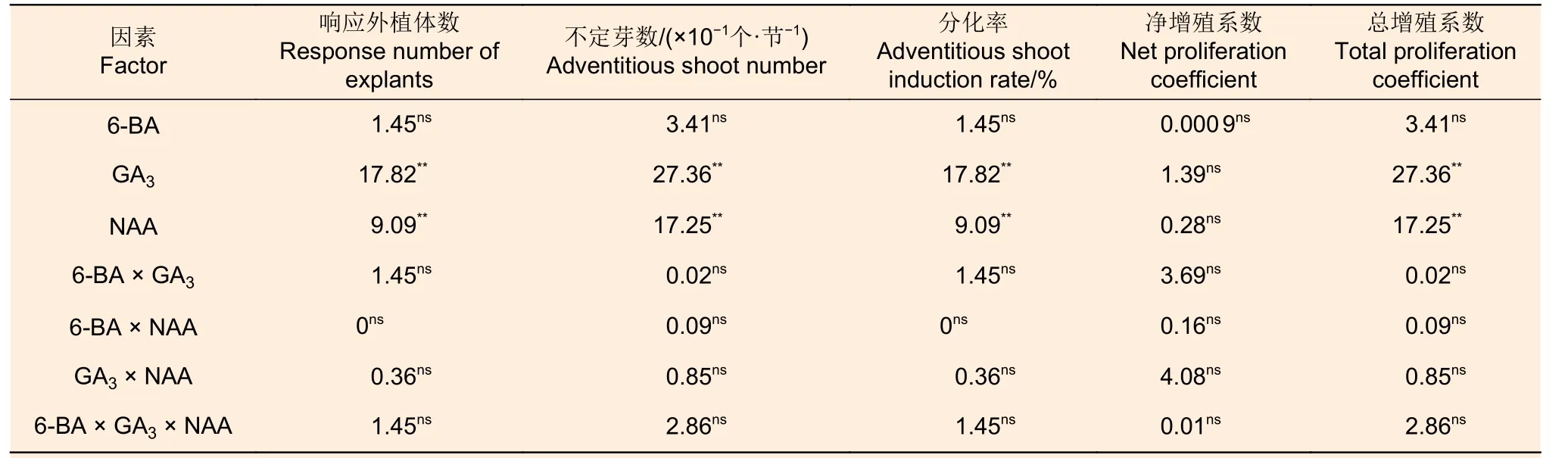
表8 西南樺不定芽分化指標的F 值和顯著性分析Table 8 F-value and statistical significance of adventitious bud differentiation of Betula alnoides
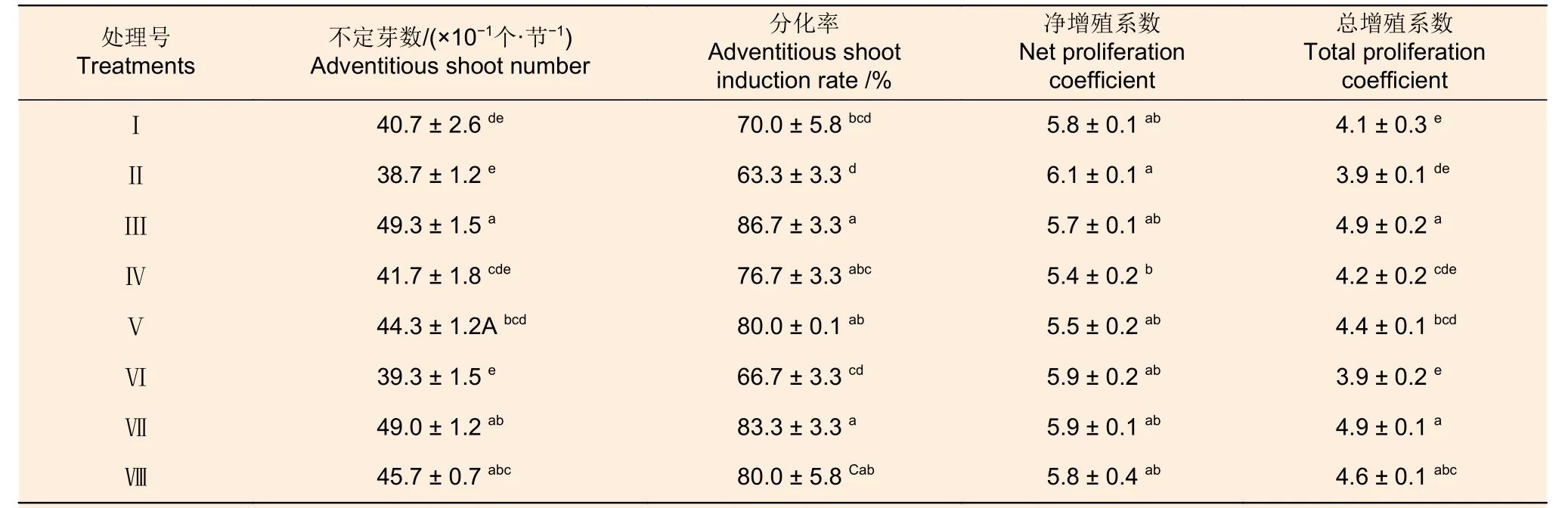
表9 不同激素組合對西南樺不定芽分化的影響Table 9 Effects of different hormone combinations on adventitious bud differentiation of Betula alnoides
2.3 不定芽生根誘導
培養12 d 和30 d 時,添加NAA 的處理(Ⅲ、Ⅳ、Ⅶ、Ⅷ)與添加IBA 的處理(Ⅰ、Ⅱ、Ⅴ、Ⅵ)間生根率和平均生根數均差異顯著(表10)。處理Ⅱ和Ⅵ,基部出現大量致密的愈傷組織,30 d 時仍未見生根;處理Ⅰ和Ⅴ,基部出現較多愈傷組織,培養12 d 時未見生根,30 d 時雖有生根但生根率低,僅為22.2%~25.0%;其它4 種處理, 12 d 時生根率為33.3%~52.9%,平均生根數為2.4~6.7,30 d 時生根率均在90%以上,平均約10 條根,平均根長4.7 cm。對照表3 可知,無論采用1/2MS 還是WPM 作為基本培養基,添加0.1 mg·L-1或0.2 mg·L-1IBA 時不定芽不生根或生根率極低,而添加0.1 或者0.2 mg·L-1NAA的生根效果均較好。相對而言,適宜西南樺不定芽生根的基本培養基為WPM,外源激素及其濃度為0.1 mg·L-1NAA,在該條件下培養30 d 生根率可達100%。

表10 不同培養基對西南樺不定芽生根的影響Table 10 Effects of different media on rooting of Betula alnoides adventitious buds
2.4 不同預培養條件對莖段待取苗生長和莖段再生的影響
方差分析結果(表11)顯示:8 種預培養條件下,處理Ⅲ和Ⅵ的株高及每株可取平均莖段數顯著高于其它處理,處理Ⅶ和Ⅷ的凈增殖系數顯著高于其它處理,8 個處理間分化率差異不顯著。西南樺株高、節間長度以及每株可取莖段數均以處理Ⅵ表現最佳,其次為處理Ⅲ,兩者的株高、節間長度以及每株可取莖段數分別在8 cm、3.5 cm 和12 以上,顯著大于其它處理。分別采用各處理的莖段誘導愈傷組織并進一步分化培養,得到的分化率亦以處理Ⅵ為最高,但各處理間差異并不顯著,均在86%以上;處理Ⅶ和Ⅷ的凈增殖系數和總增殖系數均最高,凈增殖系數在6.2 以上,總增殖系數約5.5,顯著高于絕大多數其它處理。總而言之,處理Ⅶ的再生效率最高,平均每個預培養植株可產生56.8 個不定芽,整個再生過程見圖1。因此,適宜西南樺莖段再生的預培養條件為處理Ⅶ,即弱光培養(1 000 lx)15 d 后轉至暗培養7 d,再正常光照(2 000 lx)培養8 d。

圖1 西南樺莖段再生各階段的生長狀態Fig.1 Growth status of each stage of regeneration from internodes of Betula alnoides
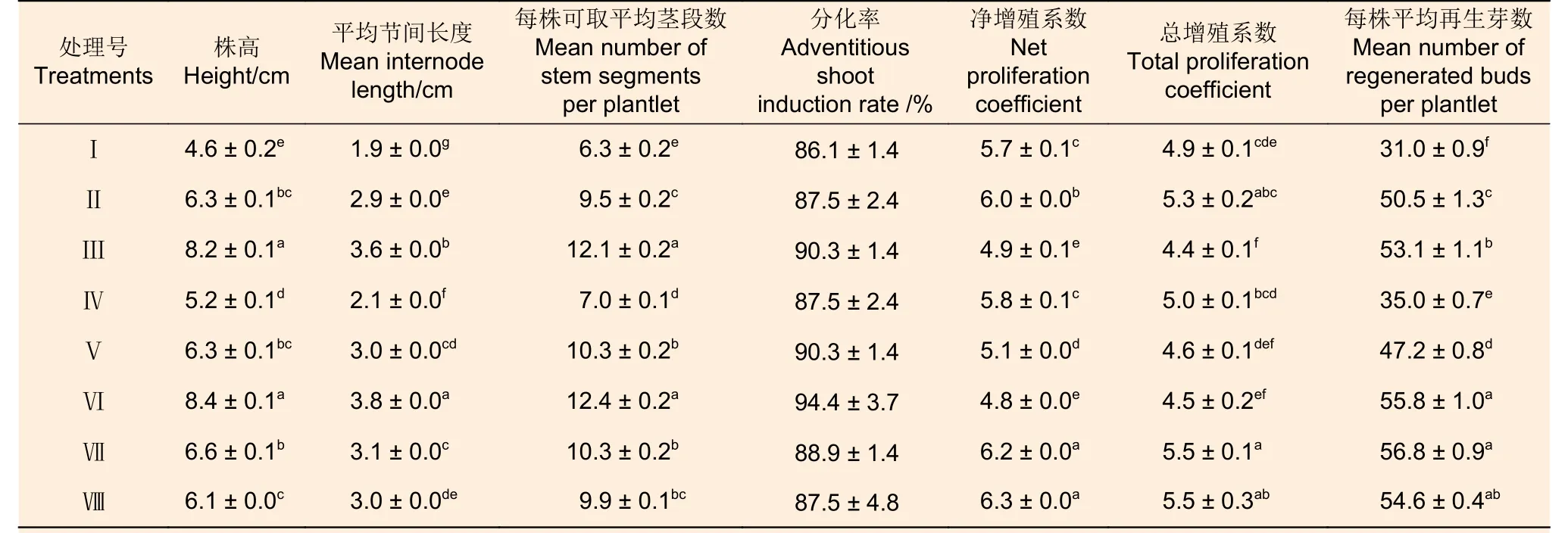
表11 不同預培養條件下西南樺植株生長表現和后續不定芽分化效果差異Table 11 Differences in growth performance and subsequent adventitious bud differentiation of Betula alnoides plantlets under various preculture conditions
3 討論
外植體的選擇是決定再生成效的第一步。幼態組織具有更高的形態發生能力,傳統培養選取靠近形態學上端的材料更易成功[23-26],但每個植株可有效利用的材料較少,從而增加了取材時間和培養成本。對于節間莖段的取材而言,適當的黑暗培養可使試管苗生長加快、節間伸長、幼嫩程度增加[27],相對于傳統培養僅能利用較短的頂端節間而言,提升其操作性的同時增加其莖段利用率。本研究通過設置8 種培養條件,篩選出再生效率和取材利用率均較高的組合,即弱光培養(1,000 lx)15 d 后,轉至暗培養7 d,接著正常光照(2,000 lx))培養8 d。與常規培養(處理Ⅰ)相比,總增殖系數和凈增殖系數分別提高8.8%和12.2%,每株平均再生芽數提高83.2%。究其原因,由于植物在黑暗中節間伸長加快,而隨著光照時間的減少,光合作用變弱,光合產物也較少,相應地維管束和機械組織較不發達,更容易脫分化形成愈傷組織[28]。亦有研究表明,一段時間黑暗培養后的黃化苗,其生長素、可溶性糖、氨基酸和蛋白質等含量均比光培養苗高,即黃化培養后的幼莖生理代謝更活躍[29],更有助于脫分化形成胚性愈傷組織,從而提高其再生效率。
愈傷組織的誘導與其基本培養基、外源激素濃度、培養時間長短等因素密切關系。木本植物通常使用MS 或WPM 培養基作為基本培養基。本研究發現西南樺莖段在MS 培養基上不定芽誘導率極低,在WPM 培養基上也并不高,而在WPB5 培養基上表現較好。與WPM 培養基相比,WPB5 換用了B5 培養基的有機物,其有機物含量更高;相比MS 培養基,WPB5 替換了有機成分的同時還降低了氮和鉀含量。由此可見,無機鹽濃度低且有機物濃度高的基本培養基更適合西南樺莖段愈傷組織誘導。分析其原因,西南樺屬于酚類物質較多的物種,培養過程中容易氧化形成醌類物質而產生褐變,從而影響外植體的脫分化和器官分化。而低鹽培養基可以適當減輕此類褐變[30],從而誘導出愈傷組織。此外,在愈傷組織形成的起動期與分裂期,外植體細胞的合成代謝活動加強,需要迅速且大量地進行蛋白質和核酸的合成,適當添加有機物質可有效促進愈傷組織誘導[31]。
細胞分裂素和生長素是西南樺莖段器官發生途徑愈傷組織誘導的必需因子,也是提高再生效率的重要因素。TDZ 具有生長素和細胞分裂素的雙重功能,可使許多難再生的植物種實現高頻分化[32-34]。如蔣淑磊等[34]對照山白杜鵑的莖段進行愈傷組織誘導研究發現,最佳TDZ 濃度為1.0 mg·L-1,愈傷組織誘導率可達96.3%,增殖系數為4.9。本研究通過設置0.1~2.0 mg·L-1的TDZ 濃度梯度開展試驗,發現隨著TDZ 濃度的增加,西南樺莖段的愈傷組織分化率和總增殖系數呈現先上升再降低的趨勢,亦以1.0 mg·L-1濃度處理表現最好,過高則會導致愈傷組織逐漸變得疏松綿軟,不利于不定芽分化。NAA 同樣對愈傷組織的誘導至關重要[35],本研究發現,NAA 濃度為0.2 mg·L-1時,其愈傷組織分化出不定芽的概率與數量均比其它濃度高;濃度為0.5 mg·L-1時,其再生效果明顯下降。關于暗培養時間,本研究結果顯示,西南樺莖段愈傷組織誘導的最佳時間為15 d,培養20 d 效果僅次于15 d,但培養10 d 效果則明顯降低。這可能與暗培養期間從培養基中吸收的TDZ 量有關,TDZ 具有快速誘導愈傷組織的效果,同時也具有劑量效應,因此外植體不宜長時間培養在該培養基上,但時間過短亦達不到相應效果。
分化率和增殖系數是決定不定芽分化效果的重要指標。本文對比研究了6-BA、GA3、NAA 3 種激素對西南樺不定芽分化的影響,發現GA3的影響最大,而且不同濃度對應的分化率和總增殖系數差異較大,濃度為0.1 和0.5 mg·L-1時,其分化率相差10%以上,總增殖系數相差0.5 以上。究其原因,GA3的使用有助于細胞伸長,加快不定芽的出芽速度,使得在統計時間段內出現較多的不定芽[36]。與之不同的是,低濃度6-BA 和NAA 更有利于不定芽分化。進一步分析發現,愈傷組織誘導階段TDZ 的添加可能影響了分化階段激素的添加。如前所述,TDZ 具有生長素和細胞分裂素的雙重功能,既可以作為一種潛在的細胞分裂素物質,又可以誘導形成部分生長素類物質。愈傷組織誘導階段高濃度TDZ 的添加使莖段愈傷組織積累了部分生長素和細胞分裂素類物質,從而較低濃度的添加即可滿足愈傷組織分化的需求[37]。就6-BA 而言,雖然濃度越高,愈傷組織表面芽點越多,但過高的細胞分裂素/生長素比值容易導致這些芽點難以成芽,最終影響再生效率。本研究還發現,不添加NAA 時不定芽分化效果更好,亦可能與愈傷組織誘導階段TDZ 和NAA 的添加積累了較多的生長素類物質有關,若在分化階段繼續添加生長素可能會導致激素配比不當,從而影響不定芽的分化。因此,綜合考慮愈傷組織誘導和不定芽分化兩階段的激素使用對再生體系的建立至關重要。
4 結論
綜上所述,本研究圍繞西南樺節間莖段再生各階段展開試驗,建立了完整的西南樺間接器官發生途徑再生體系。具體而言,增殖苗經弱光培養(1 000 lx) 15 d 后轉至暗培養7 d,再正常光照(2 000 lx) 培養8 d 可獲得適度黃化的植株,其平均株高和節間長度分別為6.6 cm 和3.1 cm;以其節間莖段為外植體,接種于添加1.0 mg·L-1TDZ和0.2 mg·L-1NAA 的WPB5 培養基上,暗培養15 d 后轉接至含0.8 mg·L-16-BA 和 0.5 mg·L-1GA3的WPM 培養基上,正常光照培養2 個月,期間繼代1 次,則可實現愈傷組織分化率和凈增殖系數達88.9%和6.2 以上,平均每個植株可產生56.8個不定芽;不定芽接種于添加0.1 mg·L-1NAA 的WPM 培養基上,培養30 d 后生根率可達100%。本研究為西南樺的遺傳改良提供了理論依據和技術支持,亦為其組培快繁開辟了一條行之有效的新途徑。
