不同培養基、生長調節劑濃度對4 個尾巨桉無性系組培生根的影響
2024-01-20 10:00:08黃安瀛陳銘秋盧萬鴻王楚彪羅建中
林業科學研究 2023年6期
黃安瀛,陳銘秋,林 彥,盧萬鴻,王楚彪,燕 青,羅建中*
(1.中國林業科學研究院速生樹木研究所,廣東 湛江 524300;2.西南林業大學,云南 昆明 650224;3.中國熱帶農業科學院湛江實驗站,廣東 湛江 524013)
隨著木材需求的日益增加、生態文明建設推進及國際木材保護性貿易政策實施力度的不斷加強,木材供應與需求間矛盾日趨嚴峻,可持續經營已成為我國林業發展的重要要求[1-2]。尾巨桉(Eucalyptus urophylla×E.grandis),是尾葉桉(Eucalyptus urophyllaS.T.Blake) 與巨桉(Eucalyptus grandisW.Mill ex Maiden)的雜交種,融合了親本速生豐產、樹干通直等特點,還具有適應性好、抗逆性強、病蟲害少等優點,在我國南方已廣泛作為紙漿、造紙、纖維板等生產原材料[3]。良種尾巨桉的大面積種植,可以降低本土木材在市場供應上的壓力,減少我國對進口木材的依賴性,對我國森林資源的培育、木材供應的安全均具有保障作用[4]。
我國桉樹新品種的開發主要采用“有性改良,無性利用”的模式,即在獲得優異的有性改良個體后,通過組織培養來擴繁、利用。組織培養(組培)具有諸多優點,但作為決定其成敗的分化、增殖及生根等過程均受多種因素影響,由各種因素導致的無法分化出根或生根質量差等,直接影響后續移栽存活率[5-7]。植物生根過程中細胞需經歷脫分化、再分化等階段,部分植物由于細胞內信號傳遞限制,相關轉錄因子基因表達受阻,次生代謝物質抑制等問題導致生根率低、生根質量差、移栽難以成活等問題[8]。因此,提升組培苗生根率,提高不定根質量在植株無性繁殖方面具有十分重要的意義。目前對尾巨桉生根已有較多研究,主要是通過改變外源生長調節劑來促進其脫分化與再分化,常用到的生長調節劑有吲哚乙酸(IBA)、萘乙酸(NAA)及ABT1。研究表明:DH32-26、DH32-28、DH32-29 等尾巨桉的最適生根培養基中添加的生長素以IBA 與ABT1 為主,當IBA 的濃度在0.2~0.3 mg·L-1,ABT1 濃度為0.3~1.5 mg·L-1時,誘導獲得的最佳生根率均在95% 以上[9-11]。以IBA 及NAA 為主的常規生長調節劑誘導生根研究中,當IBA 與NAA 的濃度分別為1 mg·L-1與0.2 mg·L-1時生根率最高可達82.5%[12],然而,盡管許多研究中試驗材料及生長調節劑種類相同,最佳配方的濃度及成分卻不盡相同。此外,ABT1生根粉作為復合配方,目前僅清楚NAA 含量20%,IBA 含量30%,其余成分未知,難以定量分析各成分對生根影響。
植物難以生根應主要受自身基因與代謝物影響,不同濃度的生長調節劑及培養基也會對其造成不同程度的影響。以幾種表型較優但生根能力不同的尾巨桉無性系為材料,通過研究不同生長調節劑種類、濃度、無性系、培養基類型對4 個尾巨桉無性系組培苗生根的影響程度,判斷這幾個因素對其不同生根階段的影響大小,分析其中的主效因子,為難生根木本植物的根系發育調控研究提供理論和實踐參考。
1 材料與方法
1.1 試驗材料
試驗的尾巨桉無性系均由中國林科院速生樹木所遺傳育種團隊自主研發,為對5 年生雜交子代測定林中長勢優良的個體環割、組培而成。取來自同一雜交組合的4 個無性系(EC262、EC264、EC269和EC272)繼代增殖瓶苗,進行不同培養基、生長調節劑處理,以研究其生根差異。
1.2 試驗方法
選取生長調節劑種類、濃度、培養基及無性系為4 個因素,設計4 因素4 水平正交試驗(表1)。預實驗中IBA 與NAA 濃度超過5 mg·L-1時4 個無性系均無法生根,故設濃度范圍1~4 mg·L-1。其中試驗組合中的A、B、C、D 分別代表培養基類型、IBA 濃度、NAA 濃度、無性系4 個因素,1、2、3、4 代表各因素不同處理水平。
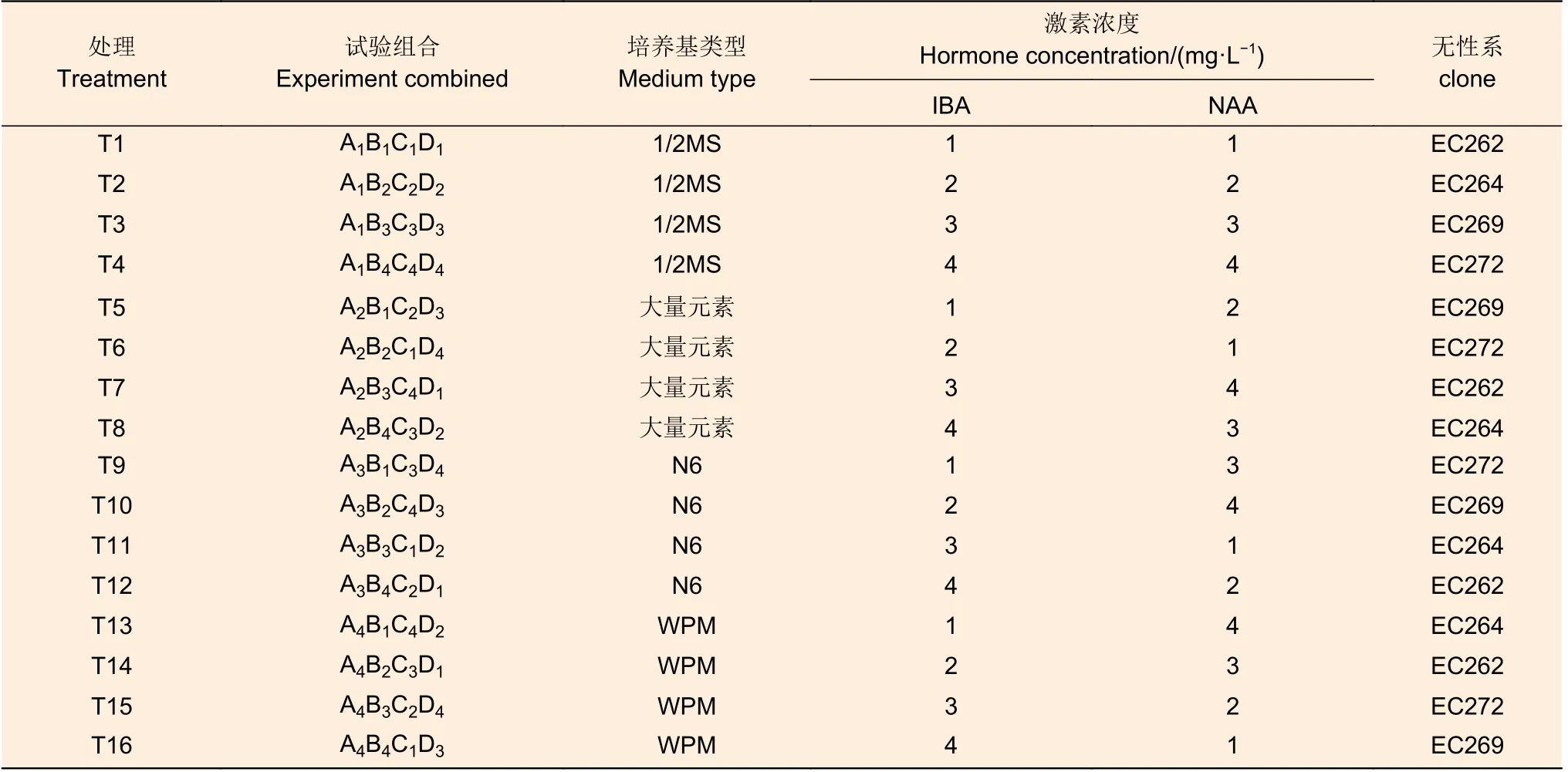
表1 正交試驗設計的因素與水平Table 1 Factors and levels for orthogonal design
1.3 培養基配置
選取1/2 MS 培養基、大量元素培養基、WPM、N6 培養基為基礎培養基,大量元素培養基配方參照王楚彪[13]提出的根誘導最適配方。培養基中添加15 g·L-1蔗糖、0.5 mg·L-1的半胱氨酸及7 g·L-1的卡拉膠,pH 值為5.8,121 ℃、20 min 高壓滅菌后取出加濾膜過濾后的植物生長調節劑。
1.4 接種與培養
選取繼代培養20~25 d,高度在4 ± 0.5 cm,生長健壯的叢生苗,剪取2 cm 以上帶頂芽部分進行生根誘導試驗[14]。暗培養7 d 后轉光照培養,培養環境為光照強度為2 000~2 500 lx,光照時長12 h,溫度28 ℃,濕度80%~90%。
1.5 測定方法
每周對植物的生長情況進行一次檢測,以評估根系的生長狀態。培育30 d 后,參考張沛建[15]的分析方法,統計各個試驗組生根情況,以生根率、平均根長、最長根長與根數等作為評價標準。生根性狀評分參考王藝[16]等方法評估,基部只有一級根及二級根,根系生長局限明顯的(圖1A),評定為1 分;基部一級根數少于3,存在二級及以上根的(圖1B),評定為2 分;基部一級根數大于3,有二級根及以上根,根系發育較好的(圖1C),評定為3 分;基部根系發育較為完善,存在三級以上側根,扎入培養基基質較為扎實的(圖1D),評定為4 分。公式計算獲得最終分數,
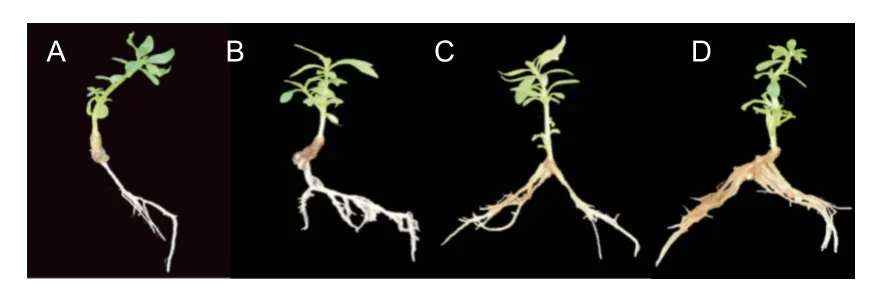
圖1 生根性狀評分示例Fig.1 The example of rooting trait scoring
其中:N1、N2、N3、N4 均為相應評分的苗數。
1.6 數據統計與分析
利用Microsoft Excel 2016 對獲得的不同試驗數據進行匯總,并利用SPSS 21.0 進行方差分析,進行多重對比研究(Duncan 法)[17]。
2 實驗結果
2.1 尾巨桉生根情況
2.1.1 生根過程 4 個尾巨桉無性系組培苗均屬于綜合生根類型,總生根數為158 株,其中皮質生根數為114(72.2%),愈傷生根數為44(27.8%)。大量元素培養基中皮質生根植株比例最高,達到86%,相對而言,皮質生根的幼苗葉片及植株生長更健壯(圖1C),而愈傷生根的根系通常側根發育更加健全,根系的擴展能力強,與培養基基質結合更加緊密,但與莖段的連接性較差,葉片及植株生長受限(圖1D)。
尾巨桉無性系生根大致分為4 個階段:(1)0~7 d,莖末端出現小型愈傷團(圖2A);(2)7~10 d,愈傷組織陸續出現小根點(圖2B );(3)10~15 d,小根點不斷膨大且數量不斷增加(圖2C);(4)15~30 d ,不定根長度快速抽長,出現二級及以上側根(圖2D);(5)30 d以上,不定根數量趨于穩定,根長持續伸長,側根持續生長 。
圖2 皮質生根不定根形成過程Fig.2 The formation of adventitious roots of E.urophylla x grandis
愈傷生根過程的主要差異體現于7~15 d,愈傷組織不出現小根點,而是持續膨大約3~5 倍,形成愈傷球。在此階段,從愈傷球中抽出根的組培苗最終會形成愈傷生根,剩余停留在愈傷階段的植株則會緩慢至停止生長。
2.1.2 不同處理對尾巨桉組培苗生根效果的影響
4 個尾巨桉無性系的根系發育情況如下表(表2),不同處理組合對無性系的生根情況影響不同,方差分析結果表明:植物生長調節劑種類和濃度、培養基類型及無性系均對植株生根的效果均具有極顯著影響。4 個無性系在不同處理下均可生根,生根率在7.5%~55%之間,其中,T2 及T6 的生根率顯著高于其它處理,其根數也同樣高于大部分處理,然而根長與生根性狀卻與這兩種生根情況的關聯性不大。各處理下4 個尾巨桉的平均根數介于1~3.27 之間,平均根長在1.95~5.02 cm 之間,最長根長在2.17~7.03 cm 之間, 生根性狀評分介于27.78%~75.75%之間。
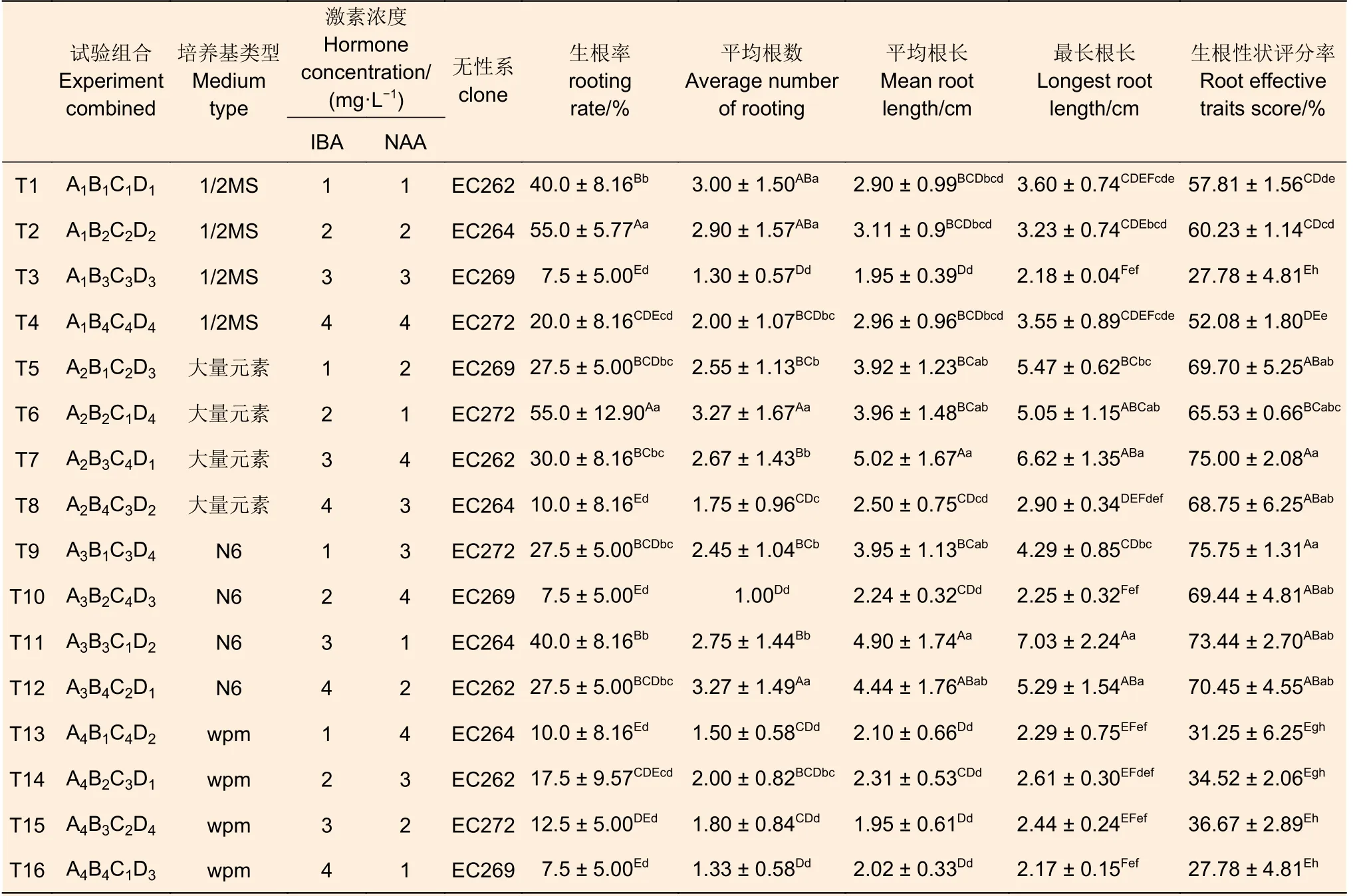
表2 各因素對尾巨桉生根的影響Table 2 Effects of different treatments on rooting of E.urophylla×E.grandis
2.2 不同處理對尾巨桉生根效果的極差分析
2.2.1 尾巨桉組培苗生根率及平均根數極差分析
結合4 個尾巨桉無性系的生根率(表3)及平均根數極差分析(表4)表明,無性系間差異是這兩種生根效果的最主要差異來源。其中,通過生根率的極差分析結果可見,無性系對生根的影響遠大于其余3 種因素,其極差達到24.5,而其余3 種因素均在7~8 之間;此外,這4 種因素在處理水平間均存在極顯著差異,其中,無性系的4 個處理中的K3(EC269)的生根率(3.5)在極顯著水平低于其余無性系(11.5~28.0)。
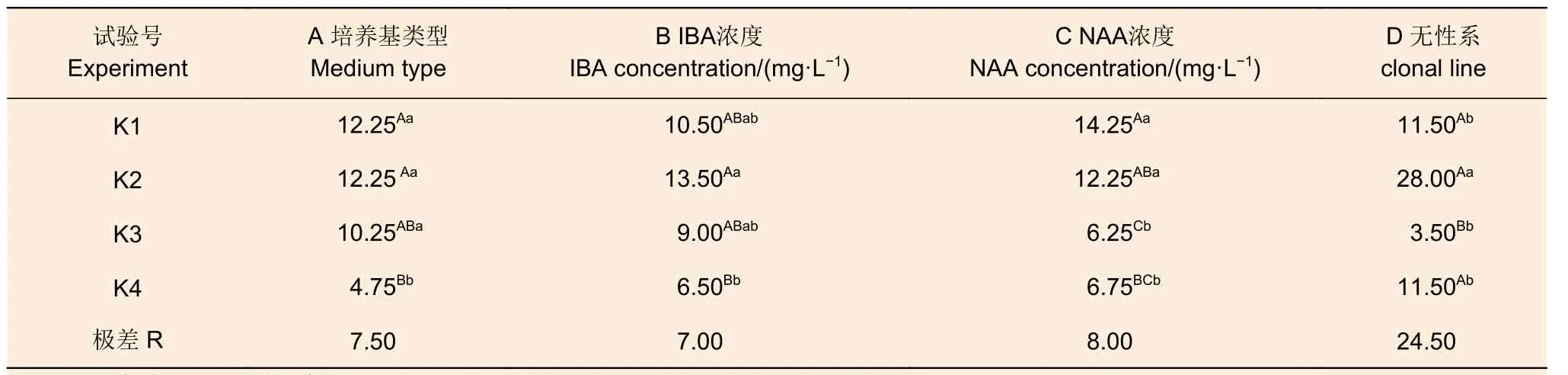
表3 尾巨桉生根率極差分析Table 3 Range analysis of the rooting rate of rooting for E.urophylla×E.grandis
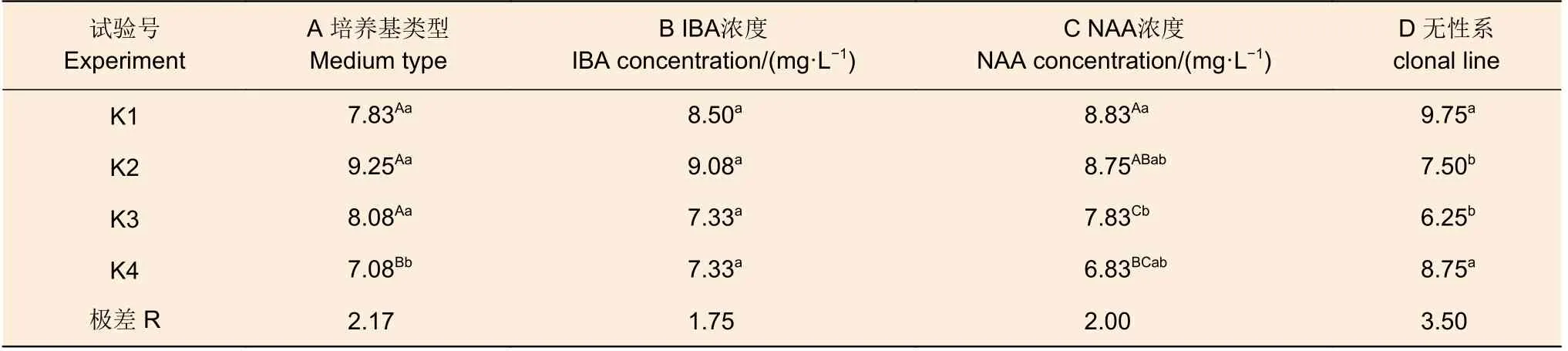
表4 尾巨桉平均根數極差分析Table 4 Range analysis of the mean number of rooting for E.urophylla × E.grandis
平均根數中幾種因素的極差較為接近,極差的范圍在1.75~3.50 間。從結果而言無性系的影響最大(3.50),其次是培養基類型(2.17)及NAA濃度(2.00)。此外,培養基類型、NAA 濃度的不同處理水平間存在0.01 水平的極顯著差異,無性系間存在0.05 水平的顯著差異,IBA 各處理間則無顯著差異。
2.2.2 尾巨桉組培苗平均根長、最長根長及生根性狀評分率極差分析 對4 個尾巨桉無性系平均根長(表5)、最長根長(表6)及生根性狀評分率(表7)的極差分析結果表明,所有因素的影響都達到顯著或極顯著水平,但各因素的重要性與生根率、平均根數中不盡相同。平均根長的極差結果顯示,培養基類型對平均根長的影響最大,極差為7.15,且處理K2(大量元素)及K3(N6)在極顯著水平上均高于其余處理。而無性系、NAA 濃度、IBA 濃度的極差分別為4.55、3.07 和2.20,重要性依次遞減;同時,在平均根長中,除無性系間存在顯著差異外,其余均為極顯著差異。
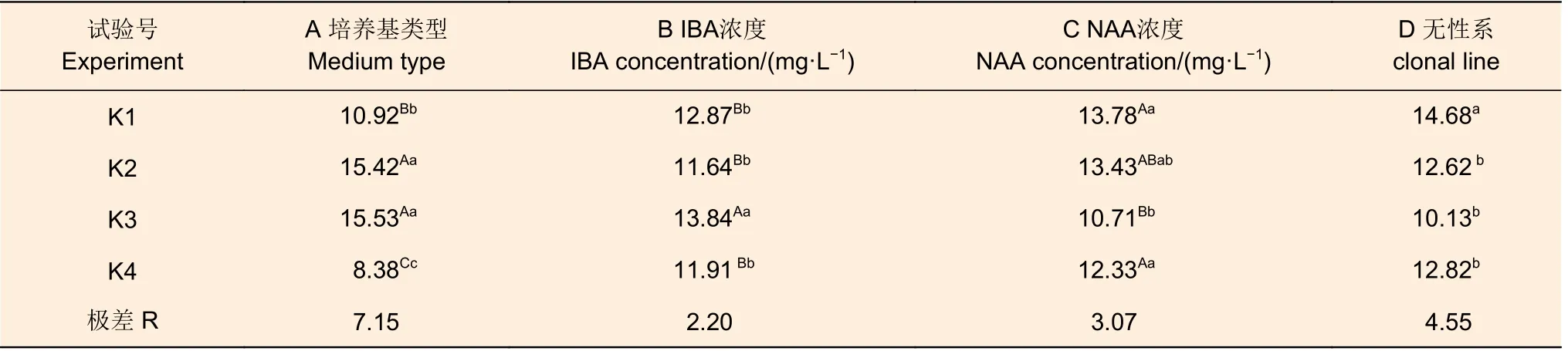
表5 尾巨桉平均根長極差分析Table 5 Range analysis of the longest root length of rooting for E.urophylla × E.grandis
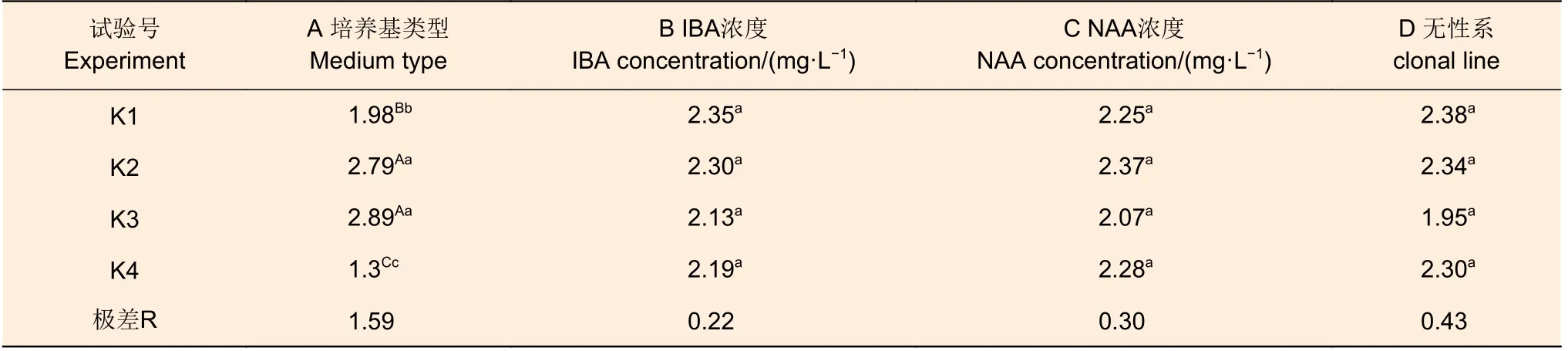
表7 尾巨桉生根性狀評分率極差分析Table 7 Range analysis of the root effective traits rate of rooting for E.urophylla × E.grandis
對于最長根長,各因素的影響大小排序與平均根長的相同(表6),也是培養基類型影響最大,極差為9.06 , 且處理K2 ( 大量元素)及K3(N6)的平均值分別為18.48 和18.75,極顯著地高于其余處理。而無性系、NAA 濃度、IBA 濃度的極差分別為7.26、5.09 和3.17,重要性依次遞減。
生根性狀評分率(表7)也是判定植株根系發育的重要指標。極差分析結果表明,培養基類型是在這個方面影響最大的因素,其極差為1.59,顯著大于無性系的0.43、NAA 濃度的0.30、IBA 濃度的0.22。4 個因素中,也僅培養基類型間的差異達到極顯著水平,其他因素內的差異均不顯著。
3 討論
3.1 不同因素對尾巨桉生根效果的影響
影響桉樹生根的原因有很多,無性系、生長調節劑濃度及培養基等均會對生根造成影響。本研究綜合考慮了這幾種因素對4 個尾巨桉無性系組培苗生根效果的影響,結果表明,植株生根不同階段的主要影響因子不同,在生根初始階段,主要是遺傳因素(無性系間差異)決定其生根差異,而在根系發育的后續階段,環境因素(培養基類型)的影響超過了遺傳因素。有研究表明,生長素與細胞分裂素是植株脫分化的必備條件,在前期預實驗中,無生長素、細胞分裂素或極高濃度(5 mg·L-1)的生長素和細胞分裂素均導致植株難以生根,說明一定濃度的生長素及細胞分裂素確是植物生根不可或缺的因素[18]。在本研究中,生長素及細胞分裂素雖是生根必須但其對植株生根的影響小于無性系或培養基類型。研究還發現,4 個尾巨桉無性系生根生長調節劑濃度及培養基適應范圍較大,1~4 mg·L-1生長調節劑濃度及4 種培養基培育下均有生根現象,生根植株均從類根愈傷組織開始生長,皮質生根在10 d 左右從莖側端生長出不定根幼根,剩余則發展為愈傷團,部分可形成愈傷生根,剩余則會逐漸褐化后死亡。
3.2 不定根形成過程影響因素
植物的基因控制著植物內源激素、信號傳導,次生代謝產物的水平,而這些產物又反過來控制基因重編程,進而控制如細胞分裂,擴大和分化等細胞過程,以此來影響植物根系的生長[19]。有研究表明,生長素IAA(Indole acetic acid)與其受體蛋白TIR1/AFB ( Transport Inhibitor Response 1/Auxin Signaling F-Box)及ARF(Auxin Response Factor)互作,在調節植物根部分生區大小及早期胚胎根形成階段,發揮了極為重要的作用[19-22]。ARF 還參與控制生長素外排因子PIN(PINFORMED)的活性[23],該蛋白通過極性運輸改變細胞間生長素流動的方向[24-26],參與介導生長素轉運的其它3 種轉錄因子PLT(PLETHORA),SHR(Short-Root)及SCR(SCARECROW),同樣被證實在影響細胞分裂,控制細胞增殖及大小等方面發揮重要作用[27-30]。在本研究中,生根率及根數受到遺傳因素的影響最大,因此,無法生根的植株可能在基因調控的某個模塊存在問題影響植株的脫分化或再分化過程,從而阻礙后續胚胎根的形成。此外,NAA 在生根階段的影響大于除無性系外的其余因素,研究表明:細胞分裂素通過表達水平和信號通路的修飾導致PIN 家族中幾種生長素外排載體基因的轉錄發生特異性變化,從而影響生長素外排和生長素在根尖的分布以此來調節生長素極性轉運控制根分生組織的活性和大小[27-28]。以此推測,生根受阻可能與生長素的轉運或與生長素轉運相關的幾種轉錄因子PIN、PLT、SHR、SCR等有關,外源添加細胞分裂素在一定程度上彌補遺傳因素缺陷,但補救能力有限。
3.3 不定根發育過程影響因素
在后續根伸長及生根性狀方面培養基占據了更為重要的地位,大量元素培養基及N6 培養基更適合尾巨桉生長,對比4 種培養基配方后發現,大量元素培養基及N6 培養基內的大量元素KH2PO4高于其它兩種培養基約3 倍,微量元素MnSO4·4H2O及ZnSO4·7H2O 含量低于其它兩種培養基約4 倍。
磷和鉀是植物生長過程中不可缺少的重要物質[29],有研究表明:磷缺乏對擬南芥(Arabidopsis thaliana(L.) Heynh.)初生根的生長具有抑制作用,而對水稻(Oryza sativaL.)及番茄(Solanum lycopersicumMill)的初生根生長卻有促進作用。GRAS 家族成員DELLA 及SCARECROW-LIKE 3(SCL3)均被證實在下胚軸和根伸長過程中發揮重要作用[30-31]。DELLA 可通過抑制光敏色素互作因子(Phytochrome-Interacting Factors,PIF)家族豐度阻止PIF 靶基因的表達來抑制下胚軸伸長[32],赤霉素通過參與的GA-GID1-DELLA 途徑參與控制根系細胞及組織伸長[31],磷缺乏導致擬南芥中赤霉素(Gibberellin,GA) 水平下降,GADELLA 信號通路的核心DELLA 蛋白累積,進一步導致編碼其代謝酶的基因轉錄物水平發生相關變化,抑制擬南芥初生根生長[33]。而在番茄中,赤霉素在其根系處于低磷環境中時起積極作用,推測GA - PRO(PRO 為番茄中DELLA 家族成員)系統可能在響應番茄磷缺乏時發揮重要作用[33]。而磷元素缺乏會導致蘋果分生細胞外質體中鐵離子積累,導致鐵毒性從而抑制初生根生長[34-35]。
此外,赤霉素信號調節在擬南芥側根分生組織的側根萌發率,密度及初生根生長等方面也同樣發揮重要作用,外源施用GA 可挽救側根(LR)短表型,K+缺乏同樣通過調控GA 信號和DELLAs相關轉錄因子調控LR 生長[36-37]。本研究推測,大量培養基及N6 培養基中充足的鉀離子及磷酸鹽可能通過赤霉素所介導的GA-DELLA 途徑及鐵離子累積方面,影響尾巨桉根的生長。
研究表明:在硫缺乏的情況下,水稻根能夠識別并誘導根產生獨腳金內脂,而在其它情況則不會[38]。獨腳金內脂被證實通過降低PIN 蛋白在木質部薄壁組織細胞質膜上的積累從而阻礙生長素的運輸,并以此影響初生根的伸長及細胞增殖[39-40]。獨腳金內脂的缺失促進了側根的形成,說明獨腳金內脂對根毛伸長也有一定影響[41]。這或許為本研究中,硫酸根缺乏的培養基反而在根系的生長上更具優勢提供一定理論參考。綜上所述,環境因素在4 個尾巨桉無性系的發育過程中發揮極為重要的作用,高濃度的磷酸根離子和鉀離子,低濃度的硫酸根離子環境更適合4 個尾巨桉無性系組培苗不定根的產生,這或許與赤霉素及獨腳金內脂兩種生長調節劑的功能相關。
4 結論
尾巨桉的生根大致分為4 個階段:0~7 d,莖末端出現小型愈傷團;7~10 d,愈傷組織陸續出現小根點;10~15 d,小根點不斷膨大且數量不斷增加;15~30 d ,不定根長度快速抽長,出現二級及以上側根;30 d 以上,不定根數量趨于穩定,根長持續伸長,側根持續生長。不同激素及培養基組合對不同無性系的生根情況影響不同,方差分析結果表明:植物生長調節劑種類和濃度、培養基類型及無性系均對植株生根的效果均具有極顯著影響。此外,極差分析結果表明:無性系是造成4 個尾巨桉的生根率及平均根數這兩種生根效果差異的最主要來源,而平均根長、最長根長及生根性狀等生根效果則受到培養基類型的影響更大。
猜你喜歡
中學生數理化·八年級物理人教版(2022年3期)2022-03-16 05:55:08
當代陜西(2021年2期)2021-03-29 07:41:24
小讀者(2021年2期)2021-03-29 05:03:48
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
媽媽寶寶(2017年3期)2017-02-21 01:22:28
紅領巾·萌芽(2016年1期)2016-09-10 07:22:44
中國塑料(2016年3期)2016-06-15 20:30:00
