吲哚在生物材料表面大腸桿菌生物膜中的研究進展
2024-01-13 13:30:00蘇丹燕唐文甜楊謹旭李邦勝趙應鼎黃云超
昆明醫科大學學報 2023年12期
蘇丹燕,唐文甜,楊謹旭,劉 華,李邦勝,趙應鼎,黃云超
(昆明醫科大學第三附屬醫院/云南省腫瘤醫院胸外一科,云南 昆明 650118)
隨著生物材料被廣泛應用于臨床,各種形式的植入手術已經成為臨床治療中不可或缺的重要組成部分,雖使成千上萬的患者從中獲益,但其帶來的高植入失敗率和再次手術風險的危害同樣不可忽視[1]。一旦治療失敗,患者不得不進行完全清創或者植入物置換手術,這不僅給患者帶來嚴重的健康損害和經濟負擔,還制約著全球衛生經濟水平的發展[2-3]。生物材料作為外源性材料,細菌在其表面定植而導致的生物膜相關感染(biofilm-associated infections,BAI)是植入物手術失敗的主要原因,但目前尚無特異性的靶向藥物,尋找靶向生物膜的抗感染藥物已成為一種研究熱點[4]。
抗生物膜治療仍是一個非常具有挑戰性的難題,為此,大量研究者致力于抗生物膜的研究,構建了聚氯乙烯(polyvinyl chloride,PVC)、硅膠等生物材料表面細菌生物膜感染模型[1,5-9],發現群體感應信號可能是生物材料植入相關感染的潛在治療靶點。細菌常常黏附于生物材料表面,形成細菌生物膜(bacterial biofilms,BF),因其結構的特殊性及致密性,常常能夠有效抵御機體的防御反應及抗生素的滲透,是導致慢性持續性感染的根源之一[10-11]。有研究發現,生物膜感染占人類微生物感染的80%[12],細菌定植在尿管導管的概率是每天5%~10%[13]。大腸桿菌是引起以生物材料為中心植入感染的優勢菌種之一,在術后生物材料植入感染中的檢出率為1.6%~2.4%[14]。研究表明大腸桿菌生物膜形成過程中還受到新型群體感應信號分子吲哚的調控[15]。本文重點就大腸桿菌生物膜、吲哚的微生物代謝及其對大腸桿菌生物膜的調控進行綜述,為臨床靶向藥物的研發提供理論依據。
1 大腸桿菌生物膜概述
1.1 細菌生物膜定義
細菌生物膜是細菌為了適應環境黏附在生物或非生物表面通過分泌多種胞外聚合物(主要是胞外多糖)包裹菌體從而保護膜內細菌免受機體攻擊及抗生素清除作用的三維立體膜狀結構,是細菌在自然界中一種常見的生存狀態[16]。
1.2 大腸桿菌生物膜形成
大腸桿菌的致病力很大程度上來自于其形成細菌生物膜的能力。不同細菌形成的生物膜雖在結構上呈現多樣性,但形成過程大致相似。大腸桿菌生物膜的形成是一個動態、循環往復的連續性過程,在不同階段,其組成比例、菌體形態、菌體數量、細菌耐藥程度及清除難度都不相同[17],一般經歷4 個生理階段,見圖1:(1)細菌黏附:包括可逆附著和不可逆黏附兩相過程。浮游細菌黏附在生物材料表面,隨著血流的沖擊、集體的吞噬作用以及抗生素的清除作用,細菌被迫遷徙或被殺滅。Brun 等[18]研究表明,通過倒置顯微鏡觀察附著階段的大腸桿菌在中等流動狀態和高剪切速率下,大腸桿菌極易被水流沖走,不利于細菌在生物材料表面進一步黏附。細菌通過菌毛、鞭毛、肽聚糖、莢膜、胞外黏質物、脂壁酸等具有黏附作用的聚合物[19]與機體組織、生物材料等特異性結合,使細菌不可逆黏附在生物材料表面,進行增殖和形成多細胞群落[18-20];(2)生物膜形成:細菌不斷生長繁殖,基因表達發生變化,分泌大量胞外聚合物,如多糖、蛋白質、脂質和細胞外DNA(eDNA)等大分子物質[21-22],將細菌包埋和固定,形成高度水合的極性化合物,有助于整體支架的構建和穩定的三維微型菌落形成;(3)生物膜成熟:生物膜逐漸發育成熟,此時生物膜體積達到最大,外觀似蘑菇狀或塔狀結構,生物膜功能更加完善,膜內營養物質實現有效分配,細菌之間通過信號分子通道進行信息交流[23];(4)脫落再黏附:內外因素的相互作用下,生物膜基質降解和表面活性劑的破壞,細胞團主動擴散,生物膜轉換為浮游狀態,游離細菌繼續附著并定植,發育成熟形成新的生物膜。附著是生物膜形成的開始,主動擴散并不是生物膜的結束,生物膜的不斷循環往復的動態過程使微生物得以不斷適應各種惡劣環境,也能夠不斷分散和定植新的生態位[24]。
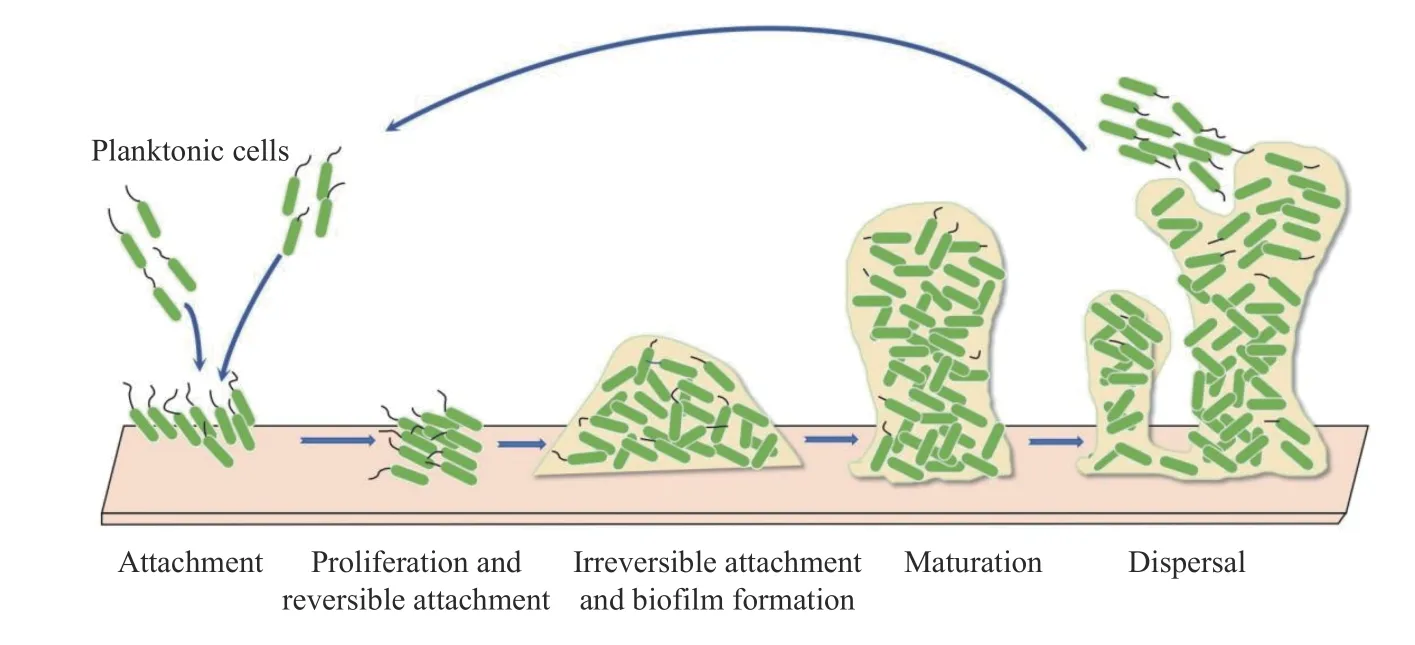
圖1 生物膜形成模式圖Fig.1 Diagram of bioflm formation
大腸桿菌作為臨床上常見的條件致病菌,在人體正常生理狀態下通常不致病,當在心臟大血管手術體外循環、骨科、泌尿外科及神經外科等術中控制性降壓,缺血再灌注損傷及臨床上失血性休克等病理情況下,人體免疫力下降,腸道內的大腸桿菌發生表型變化穿透腸壁黏膜屏障進入淋巴系統或血液系統,發生細菌移位[25],成為生物材料植入相關感染的菌源,形成細菌生物膜,導致菌血癥或敗血癥的發生,可引起遠處器官或組織的局部感染,嚴重者甚至全身感染導致MODS、SIRS 等,死亡率大大提高,甚至達37%[26]。大腸桿菌生物膜形成的調控機制目前尚未完全闡明,但近年來發現群體感應(quorum sensing,QS)對大腸桿菌生物膜形成的調控起至關重要的作用。群體感應是細菌通過產生、分泌、檢測并利用具有擴散特性的化學信號分子來感知自身及周圍環境中的細菌數量,細菌數量達到最低閾值時,細菌將會調控一系列靶基因的表達從而協調群體行為并發揮相應的生物學功能[27]。
1.3 大腸桿菌群體感應系統
群體感應系統是大腸桿菌1 種重要的信號傳遞機制。大腸桿菌通過群體感應系統接收復雜的外界環境信號并感知環境中自身的密度,從而調控胞外基質多糖的特定化學信號使細菌之間相互聚集和粘連,這對生物膜的形成和維持具有重要的意義。根據信號分子的不同,將大腸桿菌群體感應系統分為5 種:(1)由N-酰基高絲氨酸內脂(acyl-homoserine lactones,AHLs)信號分子,AHLQS 系統普遍存在于革蘭氏陰性菌中,與胞內DNA受體蛋白LuxR 結合,調控目的基因的表達[28];(2)由自身誘導肽(autoinducing peptides,AIP)介導,調控目的基因表達,參與種內細菌信號交流[29];(3)由自體誘導劑(autoinducer-2,AI-2)介導[30],調控細菌多種生物學功能,其中包括生物膜的形成[31];(4)由AI-3/腎上腺素/去甲腎上腺素介導,負責宿主-細菌間的相互交流[32-33];(5)細菌自身產生的吲哚效應分子介導的QS 系統,見圖2。吲哚通常作為大腸桿菌的生理生化鑒定標記之一,直到近年來,吲哚作為新型QS 信號分子參與細菌多種生理活動,如孢子形成[34]、質粒穩定性[35]、耐藥性[36]、吲哚產生菌的毒力[37]以及生物膜形成[38-40]等的調控作用才逐漸被揭示并重視。
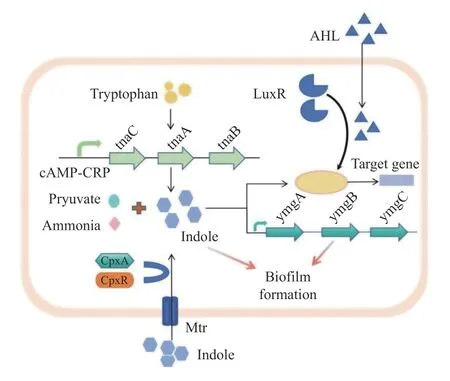
圖2 參與大腸桿菌界間通訊過程的吲哚系統示意圖Fig.2 Schematic representation of the indole system that participates in inter-kingdom communication processes in E. coli
2 吲哚的微生物代謝
吲哚是1 種典型的相對穩定的氮雜環芳烴有機化合物,在自然界中廣泛存在。微生物和動植物在進行生理生化活動過程中通常會伴隨著吲哚的產生,動物腸道中吲哚的濃度可高達1.0 mmol/L[41]。色氨酸(tryptophan,Trp)在色氨酸酶(tryptophanase,TnaA)以5′-磷酸吡哆醛為輔酶的作用下催化L-色氨酸(pyridoxal 5′-phosphate,PLP)生成吲哚,同時產物還有丙酮酸和氨[37]。大腸桿菌色氨酸酶的表達受操縱子(trpEDCBA)調控[42]。色氨酸有3 條代謝途徑,分別是犬尿氨酸途徑、5-TH 途徑、微生物代謝途徑[43]。研究發現大腸桿菌的細胞膜上存在3 種分別由Arop、TnaB 和Mtr 編碼的轉運蛋白可將胞內色氨酸轉運至胞外進行色氨酸代謝[38]。犬尿氨酸途徑承擔約95%的色氨酸代謝,是色氨酸代謝的主要途徑[44],腸道內微生物可直接或間接調節色氨酸代謝,經tnaA 編碼的酶將色氨酸轉化為吲哚,最終經尿液排出[45]。大部分生物體內均存在色氨酸合成及代謝通路,但吲哚只有在編碼tnaA 的細菌中產生[46],在低水平色氨酸的培養基條件下,Trp 操縱子的表達升高,而tna 操縱子的表達下降,吲哚生成受到抑制,與此相反,當在富含色氨酸的培養基條件下時,消除轉錄終止 Rho 依賴性,激活tnaA 基因表達,產生大量的吲哚[47]。吲哚的生成量還受到環境和生物等多種因素的影響,如溫度、pH、細胞密度、碳源及抗生素等,其中外源性色氨酸量的多少很大程度上直接影響吲哚的生成量。據報道,當提供一定濃度范圍內的色氨酸量時,進入穩定期的大腸桿菌可以把這些色氨酸轉化為等量的吲哚,最高達5.0 mmo/L[48],然而繼續加入色氨酸,生成吲哚的量卻不再增加,可能是過量的吲哚抑制TnaA 的活性以及色氨酸的轉運。
3 吲哚—信號分子新成員
細菌通過產生信號分子用于接收細胞內和細胞間的通訊信息,進行種內和種間的信息交流。目前已發現有超過85 種細菌能夠產生吲哚[12],吲哚調控細菌群感效應功能逐漸被研究者重視。盡管有研究者對吲哚是否為細胞間信號分子存在一定爭議,但相關證據表明吲哚具備成為細胞間信號分子的條件。吲哚主要產生于大腸桿菌生長的指數期晚期和穩定期的過渡期,在細菌生長遲緩期和指數期早期以非常低的水平產生[49]。研究發現[50],在小鼠腸道中吲哚水平的高低可以通過細菌膜結合組氨酸傳感器激酶(histidine sensor kinases,HK)CpxA-CpxR 雙組分系統調節腸出血性大腸桿菌(enterohemorrhagic Escherichia coli)毒力基因的表達,隨著吲哚濃度的升高,毒力基因表達下降。外源吲哚還可以誘導不產生吲哚的惡臭假單胞菌和銅綠假單胞菌中編碼兩個RND 型多藥外排操縱子基因的ttgAB 和一個編碼β-內酰胺酶ampC 的表達增加,上調CpxA-CpxR 雙組分系統,導致與抗微生物耐藥相關的基因轉錄激活,增強細菌耐藥性[51]。大腸桿菌SdiA 蛋白屬于LuxR 家族,是一種參與細胞分裂的轉錄調節因子,可以與AHL、AI-2 等信號分子相互作用[52]。有研究表明,吲哚介導的信號轉導與AHL 介導的信號轉導之間可能存在一定的聯系,高濃度吲哚抑制大腸桿菌的AHL 活性,AHL 與SdiA 結合可以免受吲哚的影響[40,53]。目前尚不清楚吲哚是如何對SdiA 同源物做出反應并發揮相應的生物學功能。此外,吲哚還通過下調噬菌體受體IV 型菌毛(T4P)活性和組裝相關基因的表達,減少T4P 介導的噬菌體吸附保護銅綠假單胞菌免受噬菌體的感染,在細菌行為調節中發揮重要的作用[54]。吲哚還可以調控非吲哚產生菌的生理活動,非吲哚產生菌通過編碼萘雙加氧酶、P540 單加氧酶和苯酚羥化酶等加氧酶對吲哚進行轉化或降解為吲哚衍生物[55]。目前,吲哚信號傳導已被證實在細菌生理生化等各個方面發揮作用,其生物學功能也不斷被揭示。
4 吲哚介導的生物膜調節作用
吲哚通過干擾大腸桿菌營養獲取和黏附定植,減弱細菌毒力,調控BF 形成相關基因的表達及群體感應系統等途徑,調控大腸桿菌的生長和BF 的形成。Wang 等[56]指出,在大腸桿菌S17-1菌株中,TnaA 缺失是抑制生物膜形成能力的主要原因,添加吲哚后,其生物膜形成能力得到恢復,證實吲哚參與調控并促進大腸桿菌生物膜的形成。Liu 等[57]實驗通過利用蛋白質的細菌通訊譜表達式(CAPPEX)來分析細菌間信息交流響應方式時發現,大腸桿菌與鼠傷寒沙門氏菌競爭Trp 以增加吲哚的產生,吲哚抑制鼠傷寒沙門氏菌HilA、SipB 和SopB 等毒力因子的表達。Sun 等[58-59]研究團隊報道指出,亞最低抑制濃度頭孢他啶(亞MIC CAZ)抑制大腸桿菌的生物膜形成能力,可能是通過增加TnaA 基因的表達和胞外吲哚濃度來實現的。Lee 等[60]研究發現,吲哚通過與基因調控蛋白YmgB 相互作用降低大腸桿菌運動性從而抑制生物膜形成。Domka 等[61]研究報道,大腸桿菌生物膜形成調控基因YliH(BssR)和YceP(BssS)通過cAMP-CRP 介導的分解代謝抑制途徑參與吲哚的調節,YliH、YceP 突變體抑制吲哚的生成,從而提高大腸桿菌運動和生物膜的形成,外源吲哚的加入使YliH、YceP 突變體大腸桿菌生物膜的形成受到抑制。
雖然目前研究者對吲哚介導BF 形成的觀點表示贊同,但因研究者所研究的實驗菌株、實驗條件的不同和難以排除內生吲哚對實驗結果的干擾,以及通過何種機制來發揮作用還是未知的,吲哚對BF 的形成起促進或是抑制作用暫無定論,還需要進一步的實驗來驗證。
5 小結
目前,隨著全球預期壽命的增長,生物材料植入人體內以替換受損的細胞、組織及器官的需求不斷增加,醫藥衛生領域以生物材料為中心植入感染問題愈加嚴峻。越來越多研究表明吲哚對于細菌生物膜的形成起著重要的調控作用,但相關機制研究仍較少,未來可能需要從以下方面進一步探索:(1)吲哚調控網絡仍需進一步完善,未來將需要更多的體內體外研究來揭示和闡明吲哚調控大腸桿菌生物膜形成的具體機制;(2)吲哚及其衍生物在抗感染治療中的應用前景及可能的不良反應;(3)吲哚信號通路可能是藥物研發的重要潛在靶點。
猜你喜歡
天天愛科學(2022年9期)2022-09-15 01:12:54
天天愛科學(2022年4期)2022-05-23 12:41:48
當代水產(2022年3期)2022-04-26 14:26:56
科學大眾(2021年9期)2021-07-16 07:02:54
鴨綠江(2021年35期)2021-04-19 12:24:18
軍事文摘(2020年20期)2020-11-28 11:42:50
考試與評價·高一版(2020年6期)2020-11-02 02:45:24
航空世界(2020年10期)2020-01-19 14:36:20
中國生殖健康(2019年3期)2019-02-01 06:12:26
鑿巖機械氣動工具(2016年3期)2016-03-01 04:00:25
