東北退化草地豬毛蒿種群繁殖分配與個體壽命可塑性適應機理
2024-01-01 00:00:00韓大勇李海燕張維楊允菲
草地學報 2024年6期




摘要:為了探討個體出生時間、采食干擾及有性生殖等過程對個體壽命的調節作用,本試驗以東北退化草地廣布種豬毛蒿(Artemisia scoparia)為研究對象,開展不同播種時間和不同去頂頻次的實驗種群,以及不同程度放牧干擾下天然種群繁殖和個體存活特征的比較研究。研究表明:播種時間處理中,早春播種種群結實后當年全部死亡,春末以后播種的結實后部分死亡;去頂處理中,無論是對照,還是1,2次去頂處理結實后均部分死亡;延遲播種時間降低了花序生物量分配和種子生物量分配比例,但去頂干擾對兩種繁殖分配均有促進作用;天然種群中,3齡級植株繁殖分配比例下降,但是種子產量增加,分別是2齡級3.6倍和1齡級3.1倍。豬毛蒿種群中個體出生時間和繁殖分配策略都會影響個體壽命,個體死亡是生殖消耗過多養分所致。
關鍵詞:種子大小;繁殖策略;冪律權衡;r-K連續統;繁殖成本
中圖分類號:Q948.1""" 文獻標識碼:A"""" 文章編號:1007-0435(2024)06-1800-10
Reproductive Allocation and Adaptation Mechanism of Individual Lifespan Plasticity
of Artemisia scoparia Population in Degraded Grassland of Northeast China
HAN Da-yong1, LI Hai-yan2, ZHANG Wei, YANG Yun-fei1,2*
(1.School of Biology and Geography Sciences, Yili Normal University, Yining, Xinjiang 835000, China; 2.Key Laboratory of
Vegetation Ecology, Ministry of Education, Northeast Normal University, Changchun, Jilin Province 130024, China)
Abstract:To explore the joint regulatory effects of birth time,feeding disturbance,and sexual reproduction of individual lifespan,Artemisia scoparia,a widely distributed species in degraded grasslands in Northeast China,was used as the study object. The reproduction and individual survival characteristics were compared among the experimental populations with different sowing times and decapitation frequencies,as well as natural population under different degrees of grazing disturbance. In the treatment of sowing time,after seeds-setting the experimental populations died entirely of those sown in the early spring and died partly of those sown in the late spring. In the treatment of decapitation,no matter the control or the experimental populations given one or two times of decapitation,all the populations died partly after seed-setting. Delayed sowing time reduced the allocation of inflorescence biomass and seed biomass,but decapitation treatments promoted two kinds of reproductive allocations. In the natural population,the proportion of reproduction allocation of 3-year-old individuals decreased,but the seed production increased,which have a seed quantity 3.6 times that of 2-year-old individuals and 3.1 times that of 1-year-old individuals,respectively. Both the birth time and reproductive allocation strategy of individuals in A. scoparia population affect individual lifespan,and individual death is caused by excessive nutrient consumption during reproduction.
Key words:Seed size;Reproductive strategy;Power-law trade-off;r-K continuum;Reproduction cost
適應與進化是生物學基礎學科共同的核心科學問題,其永恒性并不隨著科學的發展和研究技術的提高而改變。適應與進化是一種因果關系,植物的許多特征和特性都是進化的結果,都有其適應的生物學與生態學原理[1]。植物的適應有兩個方式,一個是趨同適應,結果是不同植物由于長期對相同環境適應而在外貌上表現出相同的生活型;另一個是趨異適應,結果是同種植物由于長期對不同環境適應而發生生長型分化,有的形成若干生態型,再進一步分化為新種[2]。所以,適應是物種性狀多樣性和物種多樣性的根源[3]。但是,天然植物無論在特征、特性上如何變化,一般生活型是穩定的,一種植物具有生活型多樣性的現象極為稀有。
豬毛蒿(Artemisia scoparia)是菊科蒿屬植物,其生活型具有多樣化特征。在《東北植物檢索表》第一版[4]描述為2年生草本植物;在第二版[5]描述為1,2年生草本植物;在《中國植物志》描述為多年生草本或近1,2年生草本植物[6]。國外網絡數據庫如俄羅斯的阿爾泰-薩彥嶺生態區生物多樣性網[7]描述為1,2年生。以上列舉表明國內外學術界長期以來對豬毛蒿生活型的界定一直未達成共識。根據本課題組對東北草原植物生活型和生活史特性觀測,發現在頻繁擾動生境,豬毛蒿種群由1年生、2年生和多年生的不同年齡個體組成,在放牧生境,大多數個體為多年生,在棄耕撂荒地,大多數個體為1年生和2年生,表明豬毛蒿生活型混亂的根源是個體壽命可塑性問題。
繁殖和死亡是植物生活史中的重要事件[8-10],也是種群更新和能否持續發展的基礎[11-12]。根據繁殖成本理論,對于多年生草本植物,當前的繁殖過程總是要以未來的生存、繁殖為代價,這是兩個共同進化的性狀,這一點目前已取得共識[13-14]。從理論上來說,繁殖時間和繁殖投入都存在繁殖成本,前者引出延遲繁殖和提前繁殖的問題,后者引出繁殖分配多少的問題,從而導致繁殖成本發生相應的變化[15-16]。考慮到豬毛蒿天然種群是由不同時間出生的個體組成,既存在年齡結構,也存在不同齡級個體生產力與繁殖力的差異。由此,個體壽命受個體定居時間、采食去頂干擾及有性生殖生長的分配消耗等過程的共同調節,是繁殖成本大小的反映。以往對豬毛蒿的種子重量與萌發特性[17]、根系生長[18]、葉片形態與解剖結構[19]、性別表達與繁殖輸出[20]、種內、種間競爭[21]、化感作用[22]、青蒿素生物合成的基因表達[23]等方面已有報道,而對于豬毛蒿個體存活和死亡及衰老過程與有性生殖和營養更新相聯系的系統研究未見報道。
本研究針對東北退化草地廣布種豬毛蒿的生活型多樣性和個體壽命可塑性問題,采用天然種群和實驗種群互相佐證的方法,以不同放牧干擾強度下天然種群以及不同播種時間和不同去頂頻次的實驗種群為對象,對各種群的繁殖、存活和衰老數量特征進行相應測定與定量比較分析,藉以厘清豬毛蒿個體壽命問題及其適應機理。本研究可拓展對廣布種豬毛蒿多樣化適應途徑及其進化意義的新認知,具有重要的理論意義。
1 材料與方法
1.1 研究地點
本項研究在東北松嫩平原南部、吉林省長嶺種馬場、東北師范大學吉林松嫩草地生態系統國家野外科學觀測研究站的天然草地和栽培實驗地進行。地理位置為北緯44°38′,東經123°41′附近,海拔約160 m。該區屬于溫帶半干旱半濕潤區,具有典型的季風氣候特征,年平均溫度 4.6~6.4℃,1月份的平均溫度-20.6℃,7月份的平均溫度 28.2℃,無霜期120~140 d;年平均降水量 400~500 mm,主要集中在6-8月份。年蒸發量 1 200~1 400 mm,是降水量的3倍[24]。
1.2 研究物種生活史基本特性和植株齡級劃分
豬毛蒿種子通常春、秋兩季萌發,春萌的植株開花結實后秋季死亡,在生活型上表現為1年生植物特性;秋季萌發則不能進入生殖生長階段,以幼苗越冬翌年開花結實和死亡,在生活型上表現為2年生植物特性。經觀察和實驗證明,只要有生長空間和適宜條件,豬毛蒿種子在整個生長季均可以萌發和生長,即使當年株開花結實了秋季也不全部死亡,在根頸處萌生若干芽越冬,并且這些2年生植株再次開花結實后秋季仍不全部死亡,在生活型上表現為多年生植物或地面芽植物特性。
豬毛蒿植株的年齡劃分標準是:豬毛蒿種子一般不存在休眠,當年秋季成熟、散布后萌發為實生苗,實生苗以蓮座叢越冬,翌年春季繼續生長后實生苗成長為1a株,相對應的根頸和主根由于實際上已經存活了2個生長季,其年齡為2a。秋季,在2a根頸處繼續形成萌芽,并在第3年生長成為2a株,根頸和主根為3a[25-26]。因此,按照軸根型植物的根頸營養繁殖特性來劃分豬毛蒿種群中不同植株的年齡,植株的齡級數是根頸的營養繁殖世代數。
1.3 野外取樣和處理方法
本研究同時開展不同放牧干擾強度草地的天然種群和人工小區實驗種群的調查研究。所有實驗小區播種的豬毛蒿種子均為上一年采集于棄耕撂荒地同一種群的個體,以保證基因型相對穩定。
(1)在天然草甸上,分別選擇圍欄封育未放牧(2013年開始圍封)、中度放牧(以雜類草為主)和重度放牧(以一年生鹽生植物為主)3個放牧干擾梯度的地段,于2017年6,7,8,9月份各取樣1次,調查種群組成的年齡。由于豬毛蒿在不同地段中分布不均勻,難以用單位面積作為數量衡量標準,所以采用了不固定面積的五點取樣法。具體是根據豬毛蒿個體的分布先確定5 m×5 m的面積,然后以4個角和中央為取樣點,當1 m×1 m樣方內植株數量不足50株時再擴展面積挖取,作為5次重復。取樣時保證豬毛蒿地上植株與地下主根和側根的完整性,帶回實驗室,逐株鑒別年齡[28],按齡級分置,并記錄和標記。在豬毛蒿的種子完熟期的最后一次取樣,逐齡級逐株計數根頸生芽和未生芽的株數,烘干測定株生物量、花序生物量、種子生物量和千粒重。
(2)人工建植實驗小區做兩種處理。2017年4月27日開始在東北師范大學野外生態站專用試驗田提前整地,播種地為多年翻耕草地,地勢平整,土壤質地均勻。總共建立了24塊面積為4 m×6 m的實驗小區,總面積576 m2,各小區間隔1 m。其中12塊小區用于播種時間處理,另外12塊小區用于去頂刈割處理。1)播種時間處理:分別于5月1日,5月15日,6月1日和6月15日將豬毛蒿種子撒播于紙質育苗筒中,覆土1 cm,適時澆水,再于播后1個月時將幼苗移栽至4 m×6 m的實驗小區內,重復3次。株行距為30 cm,并定期澆水,及時除雜草和防治蟲害。該處理種群為當年生,無種內和種間競爭過程。2)去頂刈割頻次處理:2017年7月15日進行移栽,2018年開始取樣。栽植密度為100株·m-2,株行距均為10 cm,移栽后做好小區澆水、除草和防蟲等管理工作。該處理種群為越年生,有種內競爭但是沒有種間競爭過程。刈割處理設置4個水平,依次為對照(不進行去頂刈割,讓其自然生長);刈割1次(于6月1日進行);刈割2次(分別于6月1日和7月1日進行);刈割3次(分別于6月1日,7月1日和8月1日進行),每個水平重復3次。每次刈割留茬高度5 cm,保留根頸萌蘗能力。在兩個實驗種群的種子完熟期,每個處理隨機取樣10株,整株挖出,分別編上序號。逐株計數根頸生芽和未生芽的株數,在80℃下烘至恒重測定株生物量、花序生物量、種子生物量、種子數量和千粒重。在生長季末期統計以上各處理下個體的存活和死亡情況。
1.4 數據統計分析
用SPSS 17.0 和Origin 2022統計軟件進行數據分析和制圖,對天然草地不同放牧干擾強度生境中豬毛蒿種群植株進行年齡譜統計。由于不同齡級之間數據非獨立,采用重復測量方差分析Duncan’s多重比較分析方法。預先對數據的球形度檢驗結果表明數據不存在自相關性。采用Bonferroni法對P值進行事后校正。
年齡譜的計算公式為:
Ri=Ni/Nt×100%
式中R為比率;i為齡級,i=1,2,3,4;Ni為樣本中某齡級植株的數量,Nt為樣本植株總數量。
生殖分配的計算公式為:
RABi=Bi/Bt×100%
RABs =Bs/Bt×100%
式中RA為生殖分配,Bi為花序生物量,Bs為種子生物量,Bt為植株地上和地下總生物量。
將兩個實驗種群的不同處理和天然種群的不同齡級分別合并為大樣本(N=30),再將其進一步合并(N=90),利用線性、冪、指數和對數函數,分別做種子千粒重與種子數量之間的相關性分析和顯著性檢驗,選用確定系數最大的方程作為種子大小與數量權衡模型。
2 結果與分析
2.1 年齡結構的變化
在天然草地生境中,各月份豬毛蒿種群植株的年齡結構隨放牧干擾強度的不同而表現出差異。總體上,圍欄無放牧和中度放牧干擾的年齡譜均以2齡級最高,均顯著高于1齡級(Plt;0.05),1齡級又均顯著高于3齡級;重度干擾的年齡譜多以2齡級高于1齡級,但兩個齡級之間差異均不顯著,而1,2齡級均顯著高于3齡級(表1)。由此表明,在圍欄無放牧地段和中度放牧干擾地段的豬毛蒿種群植株均呈穩定型年齡結構,重度放牧干擾地段為增長型年齡結構。
2.2 有性生殖數量特征的變化
經方差分析和多重比較,在4個播種時間的實驗處理中,最后1次播種處理未進入生殖生長,在其他3次播種時間處理中,豬毛蒿的花序生物量和種子生物量均隨著播種時間的延遲依次顯著下降(Plt;0.05)(圖1A1)。在4個去頂刈割的實驗處理中,3次去頂的未進入生殖生長,與對照相比,1次去頂的略有促進作用,但花序生物量和種子生物量均沒有顯著差異,2次去頂的則顯著下降(圖1B1)。由此表明,出苗時間越晚或去頂頻次越高,對花序和種子生物量的不利影響就越大,甚至不能進入開花階段。
而在相對數量特征花序(RABi)和種子(RABs)的生殖分配上,播種時間處理之間的變化與生物量特征大體相同,即隨著播種時間的延遲依次下降,RABi在3個處理之間的差異均達到顯著水平,但RABs在第1次播種和第2次播種兩個處理之間差異尚未達到顯著水平,第3次播種顯著下降(圖1A2)。去頂處理之間的變化與生物量特征完全相反,RABi隨著去頂頻次增加顯著增加,RABs在1次去頂與對照間差異不顯著,2次去頂顯著高于對照(圖1B2)。由此表明,出苗時間越晚,對RABi和RABs的不利影響也越大,但去頂干擾對兩種生殖分配均具有促進作用。
在天然種群中,在圍欄無放牧干擾草地的3個齡級中,花序生物量和種子生物量均以3齡級最大并顯著高于1,2齡級,而1,2齡級之間均無顯著差異,顯示3齡級株在花序生長和種子生物量上尚未表現衰老跡象(圖1C1)。3齡級株在兩種生殖分配上均顯著減少,其中花序生物量分配RABi在3齡級中顯著低于1,2齡級,種子生物量分配RABs在3齡級中顯著低于1齡級(圖1C2)。
2.3 種子數量和質量的變化
千粒重是評估種子質量和大小的重要指標。經方差分析和多重比較,在播種時間的實驗處理中,豬毛蒿種子數量隨著播種時間延遲依次顯著減少,以第1次播種的最多,平均達到5.2萬粒·株-1,第3次播種的為0.5萬粒·株-1,僅為第1次播種的1/10(圖2A1);而種子千粒重則相反,第1次播種的最低,平均為30 mg,并隨著播種時間延遲而顯著增加,至第3次播種的平均為61 mg,為第1次播種的2倍(圖2A2);在去頂處理中,由于個體之間的變異大,在種子數量上,1,2次去頂處理與對照均無顯著差異(圖2B1),種子千粒重只有2次去頂顯著低于對照(圖2B2)。
在天然種群中,種子數量以3齡級最多,平均達到19.8萬粒·株-1,分別是1齡級6.4萬粒·株-1的3.1倍,2齡級5.5萬粒·株-1的3.6倍(圖2C1);相反,種子千粒重以1齡級最高,平均為93 mg,是2齡級56 mg的1.7倍和3齡級38 mg的2.4倍(圖2C2)。
以上結果表明,2次去頂處理對豬毛蒿個體種子生產的數量及其質量均產生不利影響,播種時間處理的實驗種群和3個齡級的圍封無放牧干擾天然種群均具有種子數量和質量權衡趨勢。
經相關性分析和顯著性檢驗,隨著種子數量的增加,不同播種時間的實驗種群(圖3A)和天然種群(圖3B)的種子千粒重均為冪函數形式下降的確定系數最大,均達到了極顯著水平(Plt;0.01)。而不同去頂頻次的實驗種群千粒重則沒有明顯的相關關系,各處理間均在37.2 mg±6.57 mg范圍內波動(圖3C)。3個樣本再合并后擬合仍最符合冪函數形式下降,表現出權衡規律(圖3D)。
2.4 個體存活與死亡的變化
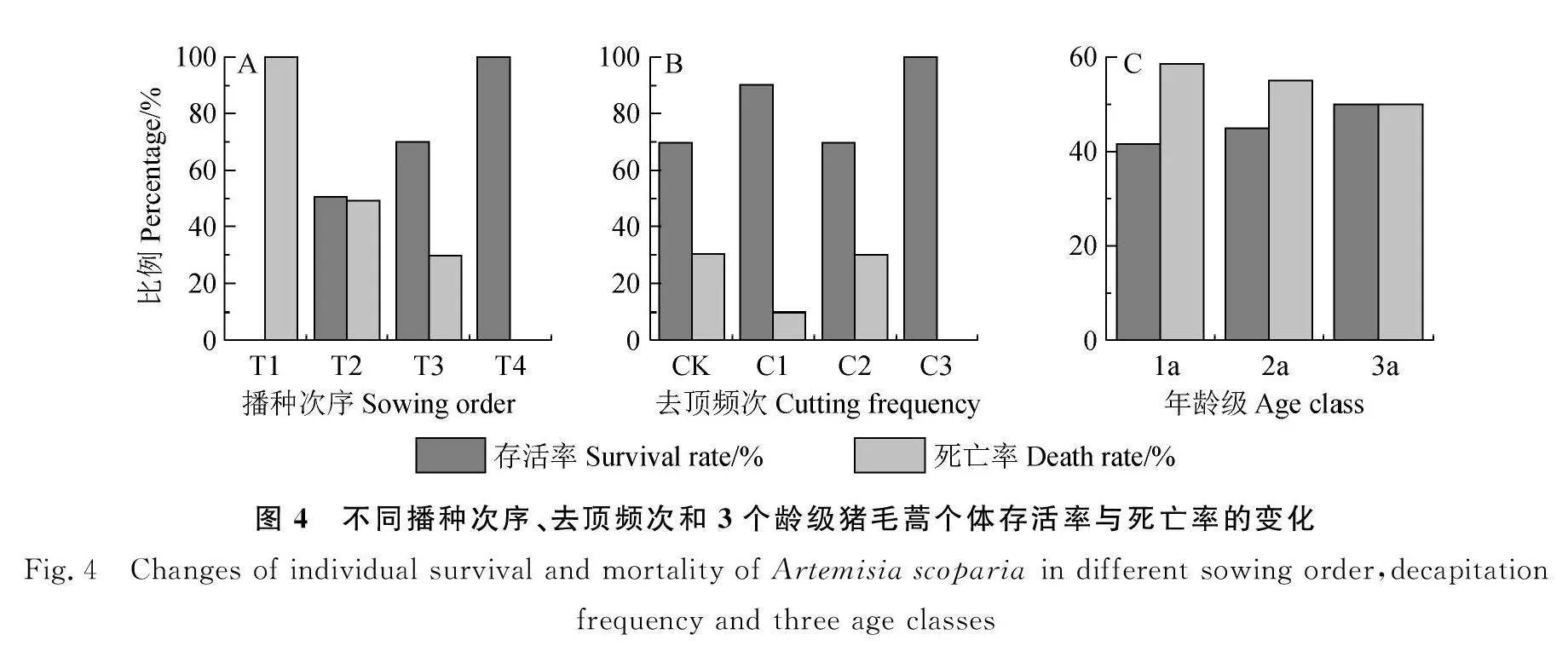
經統計,在播種時間的實驗處理中,第1次播種的豬毛蒿結實后100%死亡;第2次播種的存活與死亡各占50%;第3次播種的死亡30%;第4次播種的存活率100%(圖4A)。在去頂處理中,1次去頂的死亡率10%;2次去頂的死亡率30%;3次去頂的死亡率0%,而對照的死亡率40%(圖4B)。天然種群3個齡級的存活率和死亡率在齡級間和齡級內均變化不大,波動在42%~58%之間(圖4C)。由此表明,早春萌發且有充分生長空間的當年結實后死亡,表現為1年生植物的生活型特征,晚春以后萌發或各去頂處理,包括對照存活率均高于死亡率,即多數表現為多年生植物的生活型特征,而天然種群各齡級的存活率與死亡率接近,也表現為多年生植物的生活型特征。
3 討論
3.1 天然豬毛蒿種群個體最大壽命與年齡結構預示的種群動態
雙子葉軸根型多年生草本植物主要依靠根頸的營養繁殖維持多年生性[25]。與其他頂端生長優勢的植物一樣,豬毛蒿個體在遭到去頂干擾時主干將出現更多的分枝,只有在生長季末地上部分全部衰老死亡時,根頸的營養繁殖才得以發揮出來。因此,豬毛蒿根頸一年僅營養繁殖1個世代。本研究圍封無放牧干擾樣地的個別月份出現了4a株,其根頸的存活時間為4~5年。由此推斷在東北草原區,豬毛蒿種群中個體壽命最長為5年。天然草地不同擾動地段的豬毛蒿種群最多由4個齡級植物組成,如果把1a級視為幼齡,2a級視為壯齡,3a和4a級視為老齡,則圍欄無放牧干擾地段和中度放牧干擾地段的豬毛蒿種群植株均呈穩定型年齡結構,標志著這兩個地段是由壯齡級植株占據優勢地位,下一年的種群將穩定地繼續占據生境;在重度放牧干擾地段為1a和2a植株所占比率相近的增長型年齡結構,標志著該地段的幼齡級與壯齡級植株占據同樣的優勢地位,下一年的種群在該生境有擴展趨勢。盡管未做秋季出苗狀況的調查,但結合其根頸存活與死亡的統計,圍欄無放牧干擾地段3個齡級的存活率保持在42%~50%相同的水平,也可以反映天然種群在下一年仍然要繼續占據生境,并且即使存在越冬率,下一年種群將仍有4a級的組份。
3.2 豬毛蒿實驗種群個體的生殖分配與壽命關系
植物的繁殖和生存是適應的兩個基本量度。植物的適應性隨其生產的種子數量成比例地增加,但是繁殖要以生長和存活付出成本,需要營養代價,在個體較小時就開始繁殖的有機體,其死亡的危險性較大[14,16]。因為在生活史中的各個生命環節,如維持生命、生長和繁殖,乃至各種競爭,都要分攤有限資源。如果增加某一生命環節的能量分配,就必然要以減少其它環節能量分配為代價,這就是所稱謂的“分配原理”[27]。依此原理綜合評判在具有充分生長空間(株、行距均為30 cm)和定期清除其他雜草,既沒有種內競爭,也沒有種間競爭的當年生實驗種群中,4次播種時間處理中最后一次播種的未進入生殖生長階段。其他在生殖分配上,均隨著播種時間的延遲依次下降,并且死亡率也依次下降,最后一次播種的無個體死亡。在春季播種生長空間縮小(株、行距均為10 cm)但定期清除其他雜草僅具有種內競爭的越年生實驗種群的4個去頂頻次處理中,與對照相比,去頂處理的死亡率均有所下降,其中2次去頂處理下植株花序生物量和死亡率均是最高的,而3次去頂處理的植株未進入生殖生長階段,死亡率為0,即100%存活。天然種群3個齡級的死亡率1a為58%,2a為55%,3a為50%。以上實驗結果可以得出:1)天然種群最多由4個齡級組成,種子成熟后3個齡級均有近一半存活率,由此足以認證豬毛蒿是多年生草本;2)實驗種群在沒有種內種間競爭的早春播種的結實后當年全部死亡,春末以后播種的結實后部分死亡,證實出苗時間對個體壽命影響更大;3)在有種內競爭而沒有種間競爭的去頂處理中,無論是對照處理,還是1或2次去頂處理,結實后均有部分植株死亡,一并認證這些死亡都是有性生殖消耗營養過多而減少了個體的壽命,屬于個體壽命可塑性現象,并不是種群在特定條件下發生了1年生或2年生的生活型變異。根據以上分析,可知豬毛蒿種群中個體壽命可塑性與繁殖有關,無論是出苗時間還是去頂采食,都會降低繁殖構件生物量和分配比例,當前繁殖越少,越有利于后期個體存活,二者之間具有權衡關系,提前繁殖所需要付出的成本更高。
3.3 豬毛蒿個體壽命可塑性的適應機理
本研究獲得的結果表明,兩個實驗豬毛蒿種群的死亡均與生殖生長狀況有關。結合天然的多年生種群在生殖生長量度上,花序生物量和種子生物量均以3齡級顯著高于1或2齡級,而1,2齡級之間均無顯著差異,3齡級株不僅尚未表現衰老跡象,反而其種子數量平均達到19.8萬粒·株-1,分別是2齡級的3.6倍,1齡級的3.1倍;但秋末在根的養分貯備上卻是可溶性碳水化合物含量隨著齡級增加而下降[26],并且3齡級顯著低于1齡級而表現出衰老跡象,顯然養分貯備的減少是過多的生殖投入所致。對于多年生植物來說,個體出生當年結實后死亡,以及越年生一次結實后死亡均屬于縮短了壽命,而二次以上結實的存活者屬于延長了壽命。縮短和延長壽命均有其生存與發展的適應意義,當生長空間和資源充分時,當年生和越年生個體不惜以犧牲生命為代價,將養分資源最大化投入生殖,生產更多的種子,為種群的擴散做貢獻[28-29];當生長空間受限或遭到采食等不利干擾時便將養分資源分配到貯藏器官維持生理活動,一方面可以穩定地占據現有生境使種群繼續存在,另一方面等待時機,如本研究的3齡級直到生產更多的種子時死亡。這也意味著豬毛蒿是一種生殖目標極強的物種,即使是延長壽命的個體的死亡,也不是生理活性和生產力下降的自然衰老死亡,仍然是為生殖投入過多而犧牲奉獻。這也是在長期進化過程中豬毛蒿所形成的特有的生態適應。事實上,豬毛蒿經常成為退化草地恢復演替的先鋒種,或者是棄耕地新生境的先鋒與優勢種[30],其種群更新主要依靠種子繁殖,雖然是多年生,但個體壽命的可塑性大,在群落景觀上經常出現1年生或越年生植物普遍的“突然爆發”和“突然消失”現象,是個體壽命可塑性在生活型多樣性上的具體表現。
3.4 豬毛蒿生殖策略調節及進化意義
在植物生理學中,植物一旦花器官形成,便形成頂端生長優勢,得到養分的優先供應,并且在資源不足時停止相對的弱勢供給[31]。種子千粒重既可以代表種子的質量,也可以代表種子大小。種內的個體產籽數量和種子大小的差異均屬于因環境條件不同的可塑性響應[32-33]。如果把實驗種群的不同播種時間作為環境條件漸變系列,把去頂頻次作為外部擾動漸變系列,把天然種群的不同齡級作為個體從幼齡到老齡的漸變系列,以及把這三個系列再合并為一個綜合條件漸變系列,相關性統計的結果是,盡管在局部環節如去頂頻次干擾尚未顯露出規律性,但放在大的整體中,其種子大小與數量均遵循種群固有的冪律權衡。意味著盡管豬毛蒿在種群水平上可歸類于r策略者,但隨著生長條件的改變種群內部的個體水平上也經常進行或產生大量的小種子的r策略,或產生少量的大種子的K策略等生殖策略的調節。如果把冪律的截距附近的一些個體視為K策略者,把接近擬合曲線尾部的視為r策略者,則此冪律把r,K兩個極端類型及其過渡類型全部囊括其中而成為r-K連續統。此結果可以獲得如下認知:1)包括種子大小和產籽數量等植物數量性狀的可塑性是對環境變化適應的體現,從可塑性中尋找規律可以定量解析種群調節。所以,有序的種群策略變化是種群調節的結果。2)隸屬于r策略的豬毛蒿在生殖再分配上普遍存在r-K連續統的種群調節,具有重要適應與多維向進化意義,即在資源充分的條件下豬毛蒿采用生產更多小種子的r策略,實現將種子散布到更遠的地方而使種群向更大范圍擴張;在資源受限的條件下則采用適時停止弱勢種子生長消耗來保證生產少而更大種子的K策略,實現在種子更新中提高幼苗的存活率和在同生群中的競爭力。以往研究也一致證明在親本受資源限制時所產生的種子大小和數量之間存在權衡關系[34-35]。3)豬毛蒿作為新生境的開拓者經常生活在高度不穩定的環境中,在生存競爭和自然選擇中進化為r-K連續統,這可能也是該物種快速擴散并成為廣布種的生殖策略所致。
4 結論
在當年生實驗種群中,早春播種的豬毛蒿在結實后當年全部死亡,春末以后播種的結實后部分死亡。在越年生實驗種群中,無去頂處理和1,2次去頂處理中均出現結實后部分個體死亡現象。在天然種群中,3齡級老齡株在生殖上不僅沒有衰老跡象,反而其種子產量遠高于幼齡株,表明老齡個體也是因為生殖消耗過多養分而死亡。天然種群和實驗種群相互認證的結果是:豬毛蒿1年生和越年生植株結實后死亡均屬于個體壽命可塑性,都是有性生殖消耗過多營養而減少了個體的壽命,符合繁殖成本理論的基本假設,提前繁殖需要付出的成本更高。
參考文獻
[1] HARPER J L. Population Biology of Plants[M]. New York:Academic Press,1977:5-30
[2] SILVERTOWN J W. Introduction to Plant Population Ecology[M]. London and New York:Longman Press,1982:3-20
[3] 沈銀柱,黃占景,葛榮朝. 進化生物學[M]. 第4版. 北京:高等教育出版社,2020:127-128
[4] 劉慎諤. 東北植物檢索表[M]. 北京:科學出版社,1959:393-394
[5] 傅沛云. 東北植物檢索表[M]. 第2版.北京:科學出版社,1995:674-675
[6] 林有潤. 中國植物志第七十六卷第二分冊[M]. 北京:科學出版社,1991:220-224
[7] ЮЖНО-СИБИРСКИЙ БОТАНИЧЕСКИЙ САД АЛТГУ. БиоразнообразиеАлтае-Саянскогоэкорегиона[DB/OL]. http://bioaltai-sayan.ru/regnum/species_all.php?right=box-spec-p/artemisia.phpamp;left=go.phpamp;species=artemisia_scoparia,2018-08-21/2023-06-24
[8] HUFFAKER R C. Proteolytic activity during senescence of plants[J]. New Phytologist,1990,116(2):199-231
[9] 黃學林. 植物發育生物學[M]. 北京:科學出版社,2012:2-13
[10]THOMAS H,HUANG L,YOUNG M,et al. Evolution of plant senescence[J]. BMC evolutionary biology,2009,9:163
[11]KLIMESˇOV J,NOBIS M P,HERBEN T. Senescence,ageing and death of the whole plant:morphological prerequisites and constraints of plant immortality[J]. New Phytologist,2015,206:14-18
[12]NIKLAS K J,COBB E D. The evolutionary ecology (evo-eco) of plant asexual reproduction[J]. Evolutionary Ecology,2017,31(3):317-332
[13]OBESO J R. The costs of reproduction in plants[J]. New Phytologist,2002,155(3):321-348
[14]SLETVOLD N,GREN J. Among-population variation in costs of reproduction in the long-lived orchid Gymnadenia conopsea:an experimental study[J]. Oecologia,2011,167(2):461-468
[15]EHRLN J,VAN GROENENDAEL J. Storage and the delayed costs of reproduction in the understorey perennial Lathyrus vernus[J]. Journal of Ecology,2001,89:237-246
[16]SLETVOLD N,GREN J. Nonlinear costs of reproduction in a long-lived plant[J]. Journal of Ecology,2015,103(5):1205-1213
[17]LI X H,JIANG D M,ALANNUSA,et al. Comparison of seed germination of four Artemisia species (Asteraceae) in northeastern Inner Mongolia,China[J]. Journal of Arid Land,2012,4(1):36-42
[18]黃剛,趙學勇,蘇延桂. 科爾沁沙地3種草本植物根系生長動態[J]. 植物生態學報,2006,31(6):1161-1167
[19]杜華棟,焦菊英,寇萌,等. 黃土高原先鋒種豬毛蒿葉片形態解剖與生理特征對立地的適應性[J]. 生態學報,2016,36(10):2914-2925
[20]SHARMA I,BHARTI U,PARIHAR J,et al. Sex-expression and reproductive output in three species of Artemisia L. Abounding Jammu province (Jamp;K),India[J]. National Academy Science Letters,2014,37(3):285-288
[21]杜峰,梁宗鎖,山侖,等. 黃土丘陵區不同立地條件下豬毛蒿種內、種間競爭[J]. 植物生態學報,2006,30(4),601-609
[22]SINGH H P,KAUR S,MITTAL S,et al. Essential oil of Artemisia scoparia inhibits plant growth by generating reactive oxygen species and causing oxidative damage[J]. Journal of Chemical Ecology,2009,35(2):154-162
[23]RANJBAR M,NAGHAVI M R,ALIZADEH H,et al. Expression of artemisinin biosynthesis genes in eight Artemisia species at three developmental stages[J]. Industrial Crops and Products,2015,76:836-843
[24]鄭慧瑩,李建東. 松嫩平原的草地植被及其利用保護[M]. 北京:科學出版社,1993:45-108
[25]YANG Y F,WANG S Z,LI J D. Development and age structure of ramets of Kalimeris integrifolia populations in the Songnen Plains,Northeast China[J]. Acta Botanica Sinica,2003,45(2):158-163
[26]WANG S Y,YANG Y F,ZHI H. Water-soluble carbohydrates of root components and activity rhythms at vegetative growth stage of Artemisia scoparia in northeastern grassland of China[J]. Plos One,2017,12(5):e0176667
[27]楊持. 生態學(3版)[M]. 北京:高等教育出版社,2015:52-63
[28]武瑞鑫,鐘夢瑩,潘多,等.青藏高原東緣不同恢復措施對披針葉黃華繁殖構件及資源分配的影響[J]. 草地學報,2015,23(1):34-40
[29]左晨,王夢亭,王禮欣,等. 不同生境大托葉豬屎豆構件生物量分配的權衡特征[J]. 草地學報,2020,28(2):454-460
[30]SHI L X,GUO J X. Changes in photosynthetic and growth characteristics of Leymus chinensis community along the retrogression on the Songnen grassland in northeastern China[J]. Photosynthetica,2006,44(4):542-547
[31]潘瑞熾. 植物生理學(5版)[M]. 北京:高等教育出版社,2004:142-145
[32]GNAN S,PRIEST A,KOVER P X. The genetic basis of natural variation in seed size and seed number and their trade-off using Arabidopsis thaliana MAGIC lines[J]. Genetics,2014,198(4):1751-1758
[33]PAUL-VICTOR C,TURNBULL L A. The effect of growth conditions on the seed size/number trade-off[J]. Plos One,2009,4(9):e6917
[34]LEISHMAN M R. Does the seed size/number trade-off model determine plant community structure? An assessment of the model mechanisms and their generality[J]. Oikos,2001,93(2):294-302
[35]DEMALACH N,KADMON R. Seed mass diversity along resource gradients:the role of allometric growth rate and size-asymmetric competition[J]. Ecology,2018,99(10):2196-2206
(責任編輯 閔芝智)
