白三葉雜交F1代群體的表型鑒定及SSR分析
2024-01-01 00:00:00汪陽閆三博張睿黃琳凱張新全聶剛
草地學報 2024年6期






摘要:早期快速鑒定白三葉(Trifolium repens L.)F1雜交子代,獲取真雜種對白三葉優良品系的培育和遺傳學研究具有重要的意義。本文針對兩個白三葉親本及55個雜交F1的6個表型性狀進行分析,繪制聚類熱圖,再選用3對特異性SSR引物鑒定雜交F1代的真實性。結果表明,在聚類熱圖中57個株系可以根據親本的表型性狀劃分為父本型和母本型。篩選出的19對SSR引物共擴增共得到清晰可辨條帶281條,多態性條帶137條,多態位點占比為48.41%,每個SSR位點的PIC在18.49%~43.58%之間,平均值為34.73%。其中3對特異性引物在55個雜交后代中共鑒定出53個真雜種,真雜種比率為96.36%,與UPGMA聚類分析結果基本一致。19對SSR分子標記引物具有較高的多態性,可直接用于白三葉遺傳多樣性分析、雜種鑒定等相關研究,鑒定到的白三葉F1代真雜種為后續育種工作奠定重要基礎。
關鍵詞:白三葉;雜種鑒定;SSR分子標記;遺傳多樣性;表型分析
中圖分類號:Q344+.4""" 文獻標識碼:A"""" 文章編號:1007-0435(2024)06-1657-08
Phenotypic Identification and SSR Analysis of Trifolium repens
F1 Hybrid Progeny
WANG Yang, YAN San-bo, ZHANG Rui, HUANG Lin-kai, ZHANG Xin-quan, NIE Gang*
(College of Grassland Science and Technology, Sichuan Agricultural University, Chengdu, Sichuan Province 611130, China)
Abstract:Early and rapid identification of F1 hybrid progeny and acquisition of true hybrids are of great significance to the breeding and genetic research of Trifolium repens superior strains. Six phenotypic traits of 2 Trifolium repens parent materials and 55 F1 hybrids were mapped by clustering heat map,and 3 pairs of specific SSR primers were selected to identify the authenticity of F1 hybrids. The results showed that the 57 materials in the cluster heat map could be divided into 2 classes according to the phenotypic characters of their parents. A total of 281 distinct bands and 137 polymorphic bands were obtained by amplification with 19 pairs of selected primers. The polymorphic sites accounted for 48.41%. The PIC of each SSR site ranged from 18.49% to 43.58%,with an average value of 34.73%. A total of 53 true-hybrids were identified by three pairs of specific primers,and the true-hybrid ratio was 96.36%,which was basically consistent with the results of UPGMA cluster analysis. The 19 pairs of SSR molecular marker primers have high polymorphism,which can be directly used for genetic diversity analysis of Trifolium repens and hybrid identification and other related studies. The identified F1 generation true hybrids of Trifolium repens can lay a foundation for subsequent breeding work.
Key words:Trifolium repens;Identification of Hybrid;SSR molecular marker;Genetic diversity;Phenotypic analysis
白三葉(Trifolium repens L.)別名白車軸草、荷蘭翹搖,是豆科車軸草屬多年生草本植物,起源于地中海地區,現廣泛分布于全世界溫帶地區,因其具有蛋白質含量高、再生能力強、適應范圍廣等特點被作為優質牧草[1-2]。相關研究表明,白三葉廣泛栽培于長江中下游平原和云南、貴州、四川等低山丘陵區[3]。然而,我國白三葉育種工作相對緩慢,到目前為止國審白三葉品種僅11個(全國草品種審定委員會審定8個,國家林業和草原局草品種審定委員會審定3個),且多數為引進品種,遠不能滿足當前畜牧業的需求。因此,培育具有自主知識產權白三葉新品種,對我國白三葉的發展和應用尤為重要。
雜交育種作為國內外應用最有效和最廣泛的育種方式,其不僅能獲得集合親本優良性狀的新類型,還能通過雜種的超親分離使雜交后代群體出現超越親本的新性狀,對創造新的種質資源和培育出優良新品種具有創造性的意義[4]。但在實際雜交過程中由于各種操作因素、自然因素等,雜交后代中易出現假雜種,很大程度上延緩了雜交育種的進程。因此,鑒定F1代真實性是后續白三葉雜交育種的前提。
簡單重復序列標記(Simple sequence repeat,SSR)分子標記可快速準確的從DNA水平上揭示親代與子代間的遺傳差異[5],被廣泛應用于遺傳分析[6]、品種鑒定[7]、分子輔助育種[8]、DNA指紋圖譜構建[9]等方面。汪陽等[10]利用SSR分子標記對多年生黑麥草(Lolium perenne)4個雜交組合的78個雜交后代進行真實性鑒定,每個雜交組合分別選擇2對特異性SSR引物共鑒定出72個真雜種。謝文剛等[11]基于SSR分子標記對140個鴨茅(Dactylis glomerata)雜交種進行鑒定,利用3對特異引物快速準確鑒定出真雜種111個。馬驄毓等[12]利用SSR分子標記構建了13個白三葉品種的指紋圖譜。馬賽男等[13]利用SSR分子標記對10個白三葉品種及其近緣系的遺傳多樣性進行了分子水平的分析,發現紫色白三葉與其它9個白三葉品種的親緣關系較遠,且具有清晰的帶譜,有效與其它品種區別鑒定,為白三葉優質種質和品種的選育奠定理論基礎。但紫色白三葉其生長性能較差,因此本研究利用紫色白三葉與1個性狀優良的白三葉雜交,并對55個雜交后代進行SSR鑒定,以期篩選出具有優良性狀的雜種后代用于后續育種工作。
1 材料與方法
1.1 試驗材料
選用具有較高縮合單寧含量,葉片顏色為紫色的‘Tr-292’為母本(♀)(‘紫色白三葉’由四川農業大學提供)和生長迅速、產量較高,葉片顏色為綠色的‘Tr-445’為父本(♂)(‘Luclair increase’由新西蘭皇家科學院院士John Caradus贈予)作為親本(圖1),在四川農業大學崇州試驗基地大田先單株扦插繁殖,再進行套袋不去雄雜交。把所獲種子浸泡在98%的濃硫酸中5 min,再用蒸餾水沖洗3 min。然后將種子于同一時間播種在培養皿中,置于20℃恒溫培養箱中培養,光照/黑暗時間為12 h/12 h,光照強度為2 000 lx,相對濕度為60%。最終得到55份雜交后代,其中后代葉片顏色為綠色的有11份(以G為前綴命名),為紫色的有44份(以P為前綴命名)。當白三葉幼苗形成穩定根系時移栽到營養土中,30 d后取健壯生長單株葉片進行SSR鑒定,然后在180 d時測其表型性狀。
1.2 表型測定指標與方法
株高(Plant height,PH):采用五點取樣法,從地面至植株最高部位的絕對高度為株高。
葉柄長(Petiole length,PL):采用五點取樣法,把葉柄拉伸垂直于地面再測量時的長度為葉柄長。
葉柄粗(Petioles wide,PW):采用五點取樣法,利用游標卡尺測量葉柄的直徑。
葉面積(Leaf area,LA):隨機選取9片小葉,用圖像處理技術測量葉面積。
葉長(Leaf length,LL):隨機選取9片小葉,并壓平葉片測量其長度。
葉寬(Leaf width,LW):隨機選取9片小葉,并壓平葉片測量其寬度。
1.3 DNA提取
試驗選取生長良好的白三葉幼嫩葉片0.2 g,用CTAB法提取其基因組DNA[14],采用紫外分光光度計檢測DNA純度和濃度。最后將DNA原液的一部分用ddH2O稀釋為20 ng·μL-1,DNA樣品于-20℃冰箱內保存備用,剩余的DNA原液放于-80℃冰箱。
1.4 SSR反應體系及電泳檢測
進行PCR擴增前,將引物干粉稀釋至規定濃度放于4℃冰箱保存,PCR擴增反應體系為20 μL,其中加入10.5 μL PCR-Mix(北京天根生化公司),2 μL稀釋后的DNA,SSR上、下游引物各0.7 μL,最后加入6.1 μL ddH2O補足體積,用液體石蠟密封。PCR反應參數為:94℃預變性5 min;35個降落PCR循環:94℃變性1 min,55℃退火30 s,72℃延伸40 s;最后72℃延伸5 min,擴增完成的PCR板放于4℃冰箱保存,保存時間最好不超過一周。取出PCR板,用8%聚丙烯酰胺凝膠檢測擴增產物,在緩沖體系為1×TBE的緩沖液中進行檢測。膠版制好后固定在電泳儀上,拔出梳子后開始點樣,點樣時樣孔中每樣品加樣3 μL,以D2000 Marker為對照,恒定電壓電泳約2 h,電壓為300 V。電泳完畢后,將膠板放于0.1%的AgNO3溶液銀染15 min,隨后放入NaOH液中,放置在搖床上直至顯色,最后用相機照相保存。
1.5 SSR引物的篩選
試驗所用19條白三葉SSR標記引物序列從Griffiths等[15]中篩選得出,引物信息詳見表1。篩選可重復性好、擴增條帶清晰的引物,用于下一步雜交種的分析和鑒定。
1.6 數據統計分析
數據使用Excel 2019 進行統計整理,使用SPSS 28.0進行單因素方差分析,并采用Origin 2021繪制聚類熱圖,然后利用NTSYS PC 2.10軟件計算株系間的遺傳系數,用UPGMA法進行聚類分析,并繪制樹狀聚類圖。
2 結果與分析
2.1 白三葉雜交F1代的表型變異分析
通過SPSS 28.0對白三葉親本及雜交F1代共57個株系的株高、葉柄長、葉柄寬、葉面積、葉長和葉寬6個表型性狀進行單因素方差分析,在方差齊性檢驗顯示各樣本所在總體方差的前提下,F檢驗表明測定的6個性狀個體間差異都達到極顯著水平(P≤0.01)。此外,白三葉親本及雜交F1代的6個表型性狀的變異系數分別為17.86%,15.54%,25.54%,34.36%,17.59%和17.11%。同時‘Tr-292’的表型性狀為219.40 mm,224.40 mm,2.22 mm,834.86 mm2,38.33 mm,30.22 mm,‘Tr-445’的表型性狀為287.80 mm,241.60 mm,2.80 mm,928.93 mm2,38.00 mm,35.00 mm,雜交后代中6個表型性狀均有超親優勢的出現。表明白三葉親本及雜交F1代不同性狀在個體間具有較為豐富的變異為后續白三葉的育種工作提供了良好的基礎(表2)。
對白三葉親本及雜交F1表型進行聚類熱圖分析,結果如圖2所示:57個株系可以明顯的劃分為2類,第Ⅰ類27個雜交后代與‘Tr-445’(♂)聚在一起,雜交后代占比49.1%;第Ⅱ類28個雜交后代與‘Tr-292’(♀)聚在一起,雜交后代占比50.9%。此外,與‘Tr-445’聚在一起的雜交后代在6個表型性狀的平均值分別為291.30 mm,275.61 mm,2.56 mm,1 080.12 mm2,38.12 mm,35.93 mm,與‘Tr-292’聚在一起的雜交后代在6個表型性狀的平均值分別為227.10 mm,223.92 mm,1.78 mm,623.10 mm2,28.91 mm,27.83 mm,表明與‘Tr-445’聚在一起的雜交后代其6個表型性狀明顯優于與‘Tr-292’聚在一起的雜交后代。并且與‘Tr-445’聚在一起的雜交后代中大部分的葉色為紫色,表明其雜交后代成功獲得父母本的優良性狀,為后續白三葉育種工作提供了良好的材料。
2.2 白三葉親本及雜交種SSR擴增產物的多態性分析
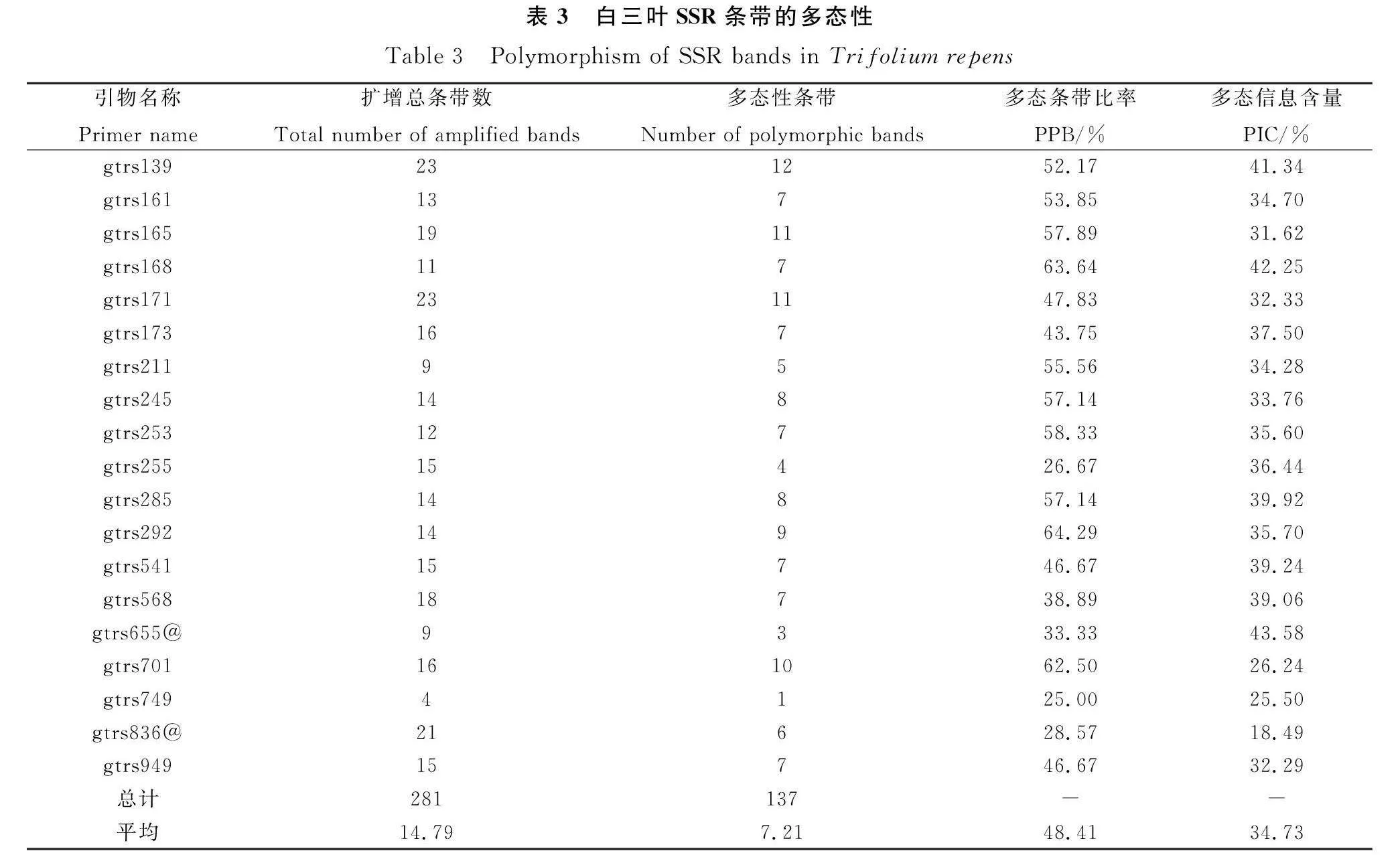
用篩選出的19對引物對白三葉2個親本及55個F1代進行擴增,共得到清晰可辨條帶281條。其中,多態性條帶137條,多態位點占比為48.41%;平均每對引物擴增出14.79條帶,多態性條帶7.21條;每個SSR位點的多態性信息含量(Polymorphism Information Content,PIC)在18.49%~43.58%之間,平均值為34.73%(表3)。
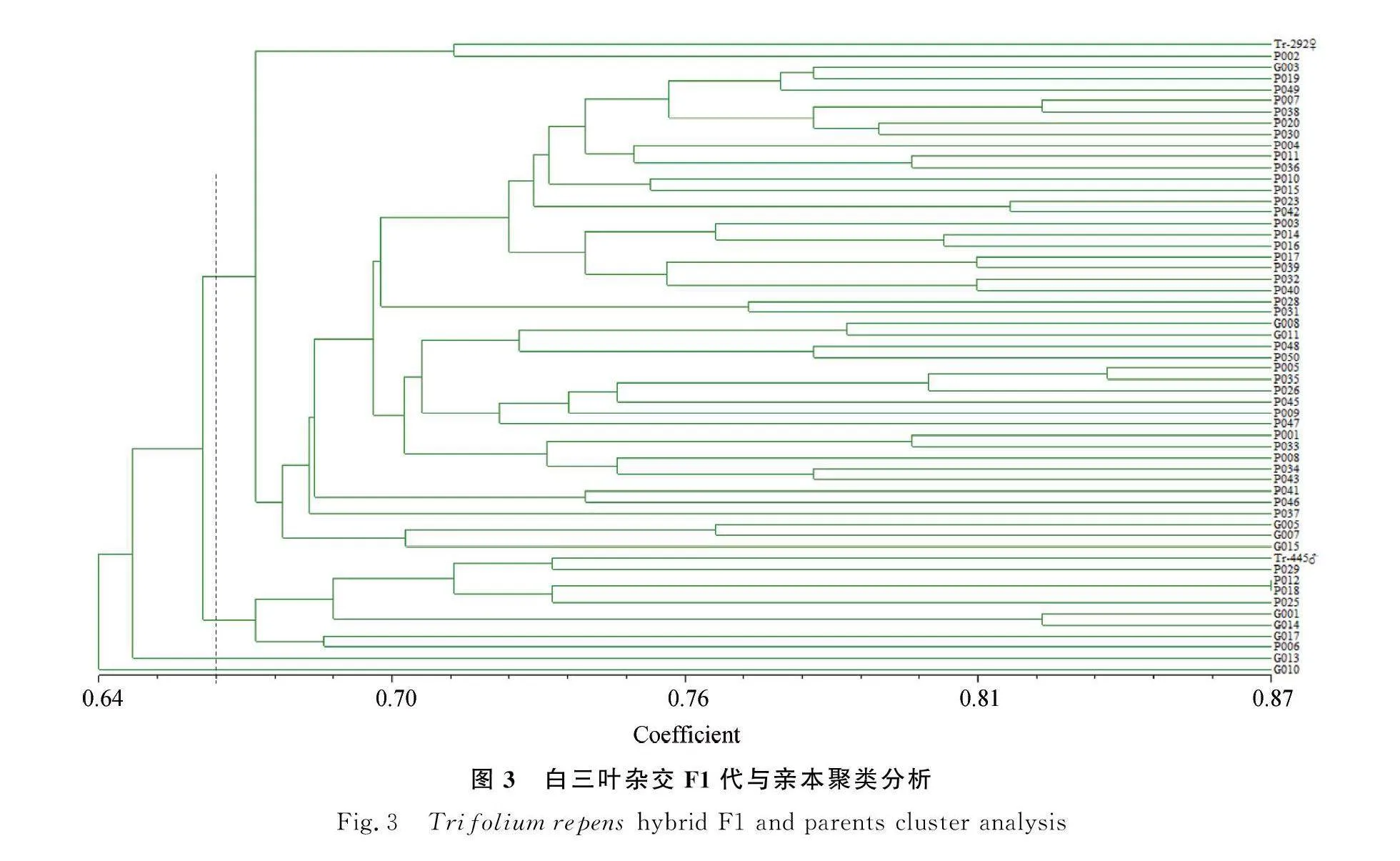
2.3 親本與其雜交F1代的聚類分析
基于遺傳相似性指數,運用NTSYS PC 2.10對2個白三葉親本與雜交F1代進行非加權組平均法(Unweighted Pair-group Method with Arithmetic mean,UPGMA)聚類分析,如圖3所示:在相似系數為0.664處可將55個F1劃分為四類,第一類是與母本‘Tr-292’聚在一起的45個雜交F1代,占81.82%;第二類是與父本‘Tr-445’聚在一起的8個雜交F1代,占14.55%;第三類是G013單獨聚在一起,占1.82%;第四類是G010單獨聚在一起,占1.82%。大部分雜交后代都與母本聚在一起,表明‘Tr-292’(♀)與‘Tr-445’(♂)雜交中,雜交后代多為偏母本遺傳。此外,單獨聚成一類的G013和G010可能是發生較大的重組變異,有利于下一步的育種工作。
2.4 雜交F1代真實性鑒定
從篩選出來的19對SSR引物中選擴增帶型好,且父本出現明顯特征條帶的引物進行F1代真假雜種的分子鑒定,選擇3對能產生互補特異位點的引物以確保鑒定的準確性。將F1代SSR引物擴增的條帶帶型與其親本比對,只要擴增出父本特異性位點的判斷為真雜種,沒有擴增出父本特異性位點為假雜種[16],引物的擴增條帶見圖4。
鑒定結果如表4所示:白三葉的55個雜交后代中真雜種53個,假雜種2個(分別為G013和G017),真雜種比率為96.36%,表明白三葉雜交獲得的真雜種比率較高。此外,引物gtrs165鑒定出真雜種29個,鑒定比率為52.73%;引物gtrs173鑒定出真雜種23個,鑒定比率為41.82%;引物gtrs245鑒定出真雜種51個,鑒定比率為92.73%。表明引物gtrs245鑒定真雜種的比率高、效果好,為加速白三葉育種工作奠定基礎。
3 討論
雜交育種作為最有效與最廣泛的育種方式,是擴大遺傳變異和獲得新類型材料的一個有效辦法[17]。但大多數異花授粉植物在雜交育種過程中后代易出現假雜種,極大地影響了其育種進程[10]。此外,白三葉屬于異源四倍體,雜交后代遺傳基礎復雜,性狀分離廣,難以從形態學上確定其真實性。SSR分子標記為共顯性標記,具有重復性好、穩定性強、操作簡單,鑒定周期短等特點,目前已成為用于鑒定雜交后代真實性的最有效方法之一[18]。李文秀等[19]篩選出3對多態性引物對橡膠樹(Hevea brasiliensis)優良品系‘PR107’和‘93-114’的35株雜交后代進行真實性鑒定,鑒定后代真雜種為100%,并驗證了其1對引物鑒定真偽雜種的可靠性。曹奕鴦等[20]應用SSR分子標記技術鑒定非洲菊(Gerbera jamesonii)F1代雜種的真實性,選用4對引物將65個雜交后代鑒定為真雜種,真雜種率達100%。陳慶明等[21]利用2對多態性引物對‘綠優’冬瓜(Benincasa hispida)和‘梅花8號’節瓜雜種種子純度進行鑒定,分子檢測平均純度均為98%,田間純度鑒定分別為98%和99%,結果基本一致。在本研究中55份雜交后代,其后代與父本顏色一致為綠色的僅有11個雜交后代,與母本顏色一致為紫色的有44個雜交后代。此外,在親本及雜交F1代的株高、葉柄長、葉柄粗、葉面積、葉片長和葉片寬6個表型性狀的聚類熱圖中,與父本表型相似的有27個雜交后代,與母本表型相似的有28個雜交后代。但在SSR分子標記鑒定中顯示,其鑒定出的假雜種僅有G013和G017這兩個雜交后代,且都在聚類熱圖中與父本表型類似。表明白三葉雜交后代遺傳復雜,僅通過表型不能完成其真實性的鑒定,SSR分子標記結果更為準確。
此外,對于SSR分子標記而言,理論上1對多態性較好的引物便可以對雜交后代完成鑒定[22]。但在實際操作中,僅用1對引物難以鑒定出全部的真雜種,其鑒定結果可能存在一定的誤差[23-24]。朱志成[25]在使用2對SSR引物鑒定玉米(Zea mays)的種子純度時發現,每個引物的鑒別效果有差異。因此為使SSR分子標記結果更加可靠,選用2對及2對以上SSR引物較為合理[10]。有研究認為,大田試驗的外界條件復雜,易出現用多對引物鑒定同一批雜交種,引物檢測結果不一致的現象[26]。因此本研究采用3對多態性引物,進一步提高SSR分子標記鑒定結果的可靠性。
雜交后代群體的遺傳變異水平與雜交的成功與否密切相關。雷雨等[18]在茶樹(Camellia sinensis)真假雜種的SSR鑒定及遺傳多樣性分析顯示,其遺傳相似系數在0.44~0.95之間,有較豐富的遺傳變異。本研究中白三葉雜交后代的遺傳相似系數為0.52~0.84,表明其雜交后代群體具有較為豐富的遺傳變異。特別是G010雖鑒定為真雜種,但在UPGMA聚類分析中單獨聚成一類,離親本的關系較遠,表明其具有較大的遺傳變異。此外,在聚類熱圖和UPGMA聚類分析中,雜交后代多數與母本聚在一起,且雜交后代在葉色上大都與母本一致,雜交子代獲得父本優良性狀較少。因此在后續雜交育種中將考慮以‘Tr-445’為母本,‘Tr-292’為父本,以期獲得更多具有父本優良性狀的雜交后代。綜上所述,白三葉雜交后代群體具有較為豐富的遺傳變異,為后續育種工作提供了良好的條件。
4 結論
對2個白三葉親本及55個雜交F1的6個表型性狀進行聚類熱圖分析,55個雜交后代可以明顯的劃分為2類,第Ⅰ類27個雜交后代與‘Tr-445’(♂)聚在一起,雜交后代占比49.1%;第Ⅱ類28份雜交后代與‘Tr-292’(♀)聚在一起,雜交后代占比50.9%。選用19對SSR引物對2白三葉親本‘Tr-292’和‘Tr-445’以及55個雜交后代進行分析,共得到清晰可辨條帶281條,多態性條帶137條,多態位點占比為48.41%,每個SSR位點的PIC在18.49%~43.58%之間,平均值為34.73%,具有較高的多態性,可直接用于白三葉遺傳多樣性分析、雜種鑒定等相關研究。3對能產生互補特異位點的引物在55個雜交后代中共鑒定出53個真雜種,真雜種比率為96.36%,與UPGMA聚類分析結果基本一致。
參考文獻
[1] ELLISON N W,LISTON A,STEINER J J,et al. Molecular phylogenetics of the clover genus (Trifolium-Leguminosae)[J]. Molecular Phylogenetics and Evolution,2006,39(3):688-705
[2] 陳菲,趙思怡,張鶴山,等. 103份白三葉種質資源萌發期耐銅性評價[J]. 分子植物育種, 2020,18(14):4829-4839
[3] 肖斌. 白三葉牧草資源研究以及豬糞尿施肥試驗報告[J]. 中國豬業,2023,18(5):122-124,129
[4] 云錦風. 牧草及飼料作物育種學[M]. 北京:中國農業出版社,2016:72-83
[5] 朱駿馳,郭印山,劉鎮東,等. 利用SSR分子標記鑒定葡萄F1代雜種[J]. 沈陽農業大學學報,2016,47(2):148-152
[6] XIE W G,ZHANG X Q,MA X,et al. Genetic variation and rela tionship in orchardgrass(Dactylis glomerata L.) germplasm detected by SSR markers[J]. Hereditas(Beijing),2009,31(6):654-662
[7] 尚維,趙申清玉,黨江波,等. 基于SSR分子標記的Nicotiana tobacum-N.plumbaginifolia異源染色體植株的鑒定與篩選[J]. 作物學報,2018,44(11):1640-1649
[8] 陳彪,陳銘,李洋洋,等. 煙草糖酯SSR分子標記篩選及輔助育種應用[J]. 中國煙草科學,2019,40(3):8-15
[9] 張迪,任立飛,張璐璐,等.基于熒光毛細管電泳技術的苜蓿SSR指紋圖譜構建[J]. 中國草地學報,2020,42(6):10-14
[10]汪陽,易莉美,班晚婷,等. 基于SSR分子標記的多年生黑麥草雜交F1代雜種鑒定[J]. 中國草地學報,2021,43(11):10-17
[11]謝文剛,張新全,陳永霞. 鴨茅雜交種的SSR分子標記鑒定及其遺傳變異分析[J]. 草業學報,2010,19(2):212-217
[12]馬驄毓,韓重陽,馬賽男,等. 基于SSR分子標記的13個白三葉(Trifolium repens L.)品種指紋圖譜構建[J]. 草地學報,2021,29 (9):1892-1899
[13]MA S N,HAN C Y,ZHOU J,et al. Fingerprint identification of white clover cultivars based on SSR molecular markers[J]. Molecular Biology Reports,2020,47(11):8513-8521
[14]劉亞令,耿雅萍,解瀟冬,等. 基于SSR分子標記的藥用黃芪遺傳多樣性與遺傳結構分析[J]. 草地學報,2019,27(5):1154-1162
[15]GRIFFITHS A G,BARRETT B A,SIMON D,et al. An integrated genetic linkage map for white clover (Trifolium repens L.) with alignment to Medicago[J]. BMC Genomics,2013,14(1):388
[16]韓國輝,向素瓊,汪衛星,等. 沙田柚雜交后代群體的SSR鑒定與遺傳多樣性分析[J]. 中國農業科學,2010,43(22):4678-4686
[17]EECKHAUT T,KEYSER E D,HUYLENBROECK J V,et al. Application of embryo rescue after interspecific crosses in the genus Rhododendron[J]. Plant Cell Tiss.Organ Cult,2007,89(1):29-35
[18]雷雨,段繼華,黃飛毅,等. 茶樹雜交F1代真假雜種的SSR鑒定及遺傳多樣性分析[J]. 植物遺傳資源學報,2021,22(3):748-757
[19]李文秀,賀軍軍,張華林,等. SSR分子標記鑒定橡膠樹F1真偽雜種[J]. 熱帶作物學報,2021,42(5):1305-1309
[20]曹奕鴦,夏朝水,陳瑋婷,等. 應用SSR分子標記鑒定非洲菊F1代雜種的真實性[J]. 福建農業學報,2023,38(7):842-850
[21]陳慶明,劉政國,凌志陽,等. 冬瓜雜種種子純度鑒定的SSR分析[J]. 分子植物育種,2020,18(8):2613-2618
[22]賈慶麟,張瑞花,王文強,等. 柱花草EST-SSR標記的鑒定及應用[J]. 分子植物育種,2016,14(5):1195-1202
[23]鄧衍明,賈新平,梁麗建. 觀賞植物遠緣雜交后代的鑒定方法[J]. 核農學報,2016,30(7):1308-1315
[24]GIANFRANCESCHI L,SEGLIAS N,TARCHINI R,et al. Simple sequence repeats for the genetic analysis of apple[J]. Theoretical and Applied Genetics,1998,96(8):1069-1076
[25]朱志成. SSR分子標記在玉米品種良玉88純度鑒定上的應用[J]. 農業與技術,2021,41(2):54-55
[26]何俊平,張書芬,王建平,等. 甘藍型雙低油菜雜交種豐油10號純度的SSR鑒定[J]. 作物雜志,2019(1):75-80
(責任編輯 劉婷婷)
