云南5個典型高原湖泊沉水植物群落組成及其與環境因子的關系
2024-01-01 00:00:00朱利明張道熙史方鄒曦張志永胡蓮
水生態學雜志 2024年5期


摘要:通過對云南中部5個典型高原湖泊沉水植物和水環境狀況進行野外調查,研究該區域主要沉水植物群落組成及其與環境因子的關系,為云南高原湖泊沉水植被保護和管理提供理論參考。綜合營養狀態指數計算表明,所選取的5個湖泊中,異龍湖、杞麓湖和星云湖屬于富營養湖泊,撫仙湖和陽宗海屬于貧-中營養湖泊。調查共發現沉水植物6科10屬15種,微齒眼子菜(Potamogeton maackianus)和穿葉眼子菜(P. perfoliatus)在貧-中營養湖泊中為主要優勢種,穗花狐尾藻(Myriophyllum spicatum)和篦齒眼子菜(P. pectinatus)在各營養類型湖泊中均能成為優勢種或亞優勢種,輪葉黑藻(Hydrilla verticillata)、菹草(P. crispus)和金魚藻(Ceratophyllum demersum)僅在富營養湖泊中成為優勢種。沉水植物多樣性分析表明,撫仙湖的香農-威納多樣性指數(H')最高,平均值達到1.63;杞麓湖的H'最低,平均值僅0.79,總體來說,貧-中營養湖泊的沉水植物多樣性要高于富營養湖泊。沉水植物群落物種與水環境因子的冗余分析(redundancy analysis, RDA)結果顯示,透明度(SD)、pH、硝氮(NO3--N)是影響這些不同營養水平湖泊沉水植物分布特征的關鍵環境因子,pH、氨氮(NH4+-N)、水溫(WT)是影響貧-中營養湖泊沉水植物分布特征的關鍵環境因子,WT、電導率(SPC)、葉綠素a(Chl-a)是影響富營養湖泊沉水植物分布特征的關鍵環境因子。研究對于恢復、保護和管理該區域湖泊沉水植物具有重要指導意義。
關鍵詞:高原湖泊;沉水植物;空間分布;環境因子;冗余分析
中圖分類號:Q178.1" " " " 文獻標志碼:A" " " " 文章編號:1674-3075(2024)05-0039-10
沉水植物是淡水湖泊生態系統主要的初級生產者,在湖泊物質循環和能量流動中發揮著重要作用,是維持湖泊清水穩態的關鍵(Wang et al,2016)。沉水植物生命活動對湖泊環境因素的變化非常敏感,水體營養鹽濃度、底泥類型、水文水動力條件、濁度、透明度、溶解氧、水深等諸多環境因子都與沉水植物的群落組成與空間分布密切相關(Nakamura et al,2008;Bornette amp; Puijalon,2011;孔祥虹等,2015)。近些年來,隨著社會經濟的快速發展,湖泊富營養化日益加重,我國多個湖泊沉水植物群落演替劇烈,群落組成和優勢物種發生了明顯的改變,最終導致生物量銳減、多樣性減少,湖泊生態系統功能受到嚴峻挑戰(Sachse et al,2014)。
湖泊富營養化會導致水生植物群落結構發生變化,這與富營養水體本身對水生植物的脅迫、富營養促進浮游植物與水生植物之間的競爭以及富營養促進單一物種的大量繁殖擴散等因素有關(Kalff,2002)。富營養化引起水生植物群落退化、生物多樣性降低的內在機理是不同水生植物對水環境的適應范圍存在差異(吳愛平等,2005;游灝,2006),耐受范圍小、生態位寬度窄的物種對水環境的變化更為敏感。有沉水植物存在的水域水質通常較好。有研究表明,沉水植物具有富集水體污染物的功能,在富營養化水體修復過程中起到凈化水質、維持水生態系統健康的作用(Carpenter amp; Lodge,1986)。當前,國內外眾多學者在沉水植被恢復重建領域開展了大量研究,光照強度(李文朝和連光華,1996)、營養鹽(Horppila amp; Nurminen,2005;金相燦等,2007)、底質(謝貽發等,2007)、濁度(王文林等,2006)、溫度(Barko amp; Smart,1981;閆志強等,2014)等因素對沉水植物的生長至關重要,共同影響沉水植物的群落組成與分布(王華等,2008)。因此,研究沉水植物群落組成及分布格局與水環境因子的關系具有重要意義。
云南是我國淡水湖泊資源最為集中的地區之一,其湖泊主要分布在斷裂帶或各大水系的分水嶺地帶,具有海拔較高、湖岸陡峻、面積小而水深的特點(曾理等,2009),兼具調節氣候、水量,提供生產生活用水、水產養殖產品,美化環境和發展旅游的功能(藍紅林,2001)。這些湖泊多為源頭型湖泊,其生態狀況同時影響著當地和下游平原地區的生態環境(楊柳和黃耀志,2002)。然而,隨著該地區工農業的發展以及長期不合理的湖泊資源開發,云南高原湖泊生態環境遭到嚴重破壞,富營養化嚴重,水體營養梯度日益明顯,生態系統日益退化,這一系列問題已引起眾多學者和社會的關注(顏昌宙等,2005)。以往的調查多由不同研究者單獨進行,對多個湖泊同時期沉水植物分布情況了解不足。為了深入了解云南高原湖泊沉水植物群落組成及分布格局,本文對云南5個典型高原湖泊沉水植物進行了系統調查,比較分析該區域沉水植物的區系特征,探明沉水植被現狀,對影響沉水植物分布的水環境因子進行解析,辨識關鍵影響因子,以期為云南高原湖泊沉水植被保護和管理提供理論參考。
1" "材料與方法
1.1" "研究區域
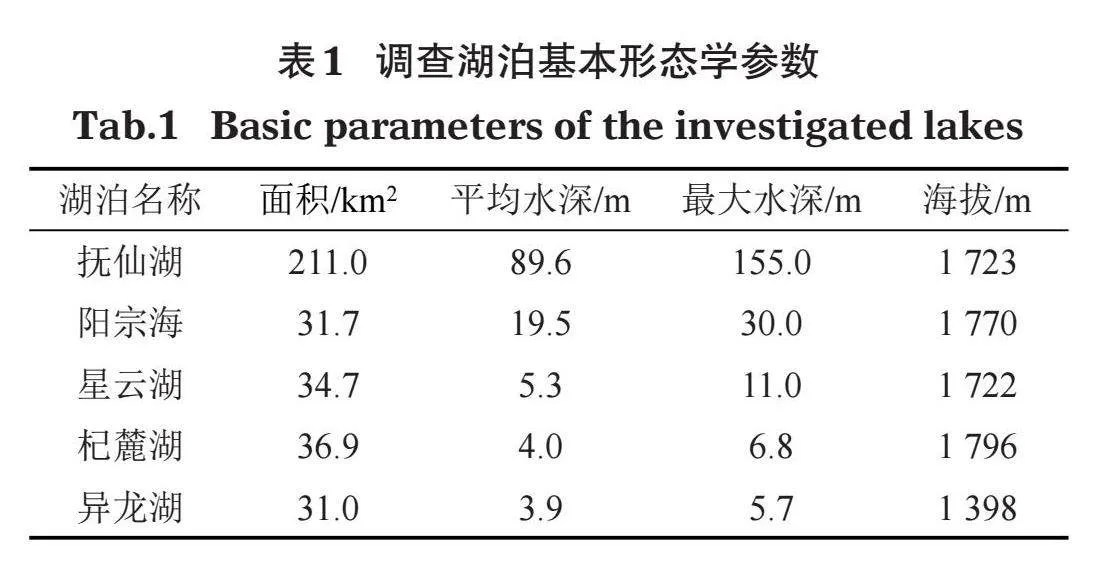
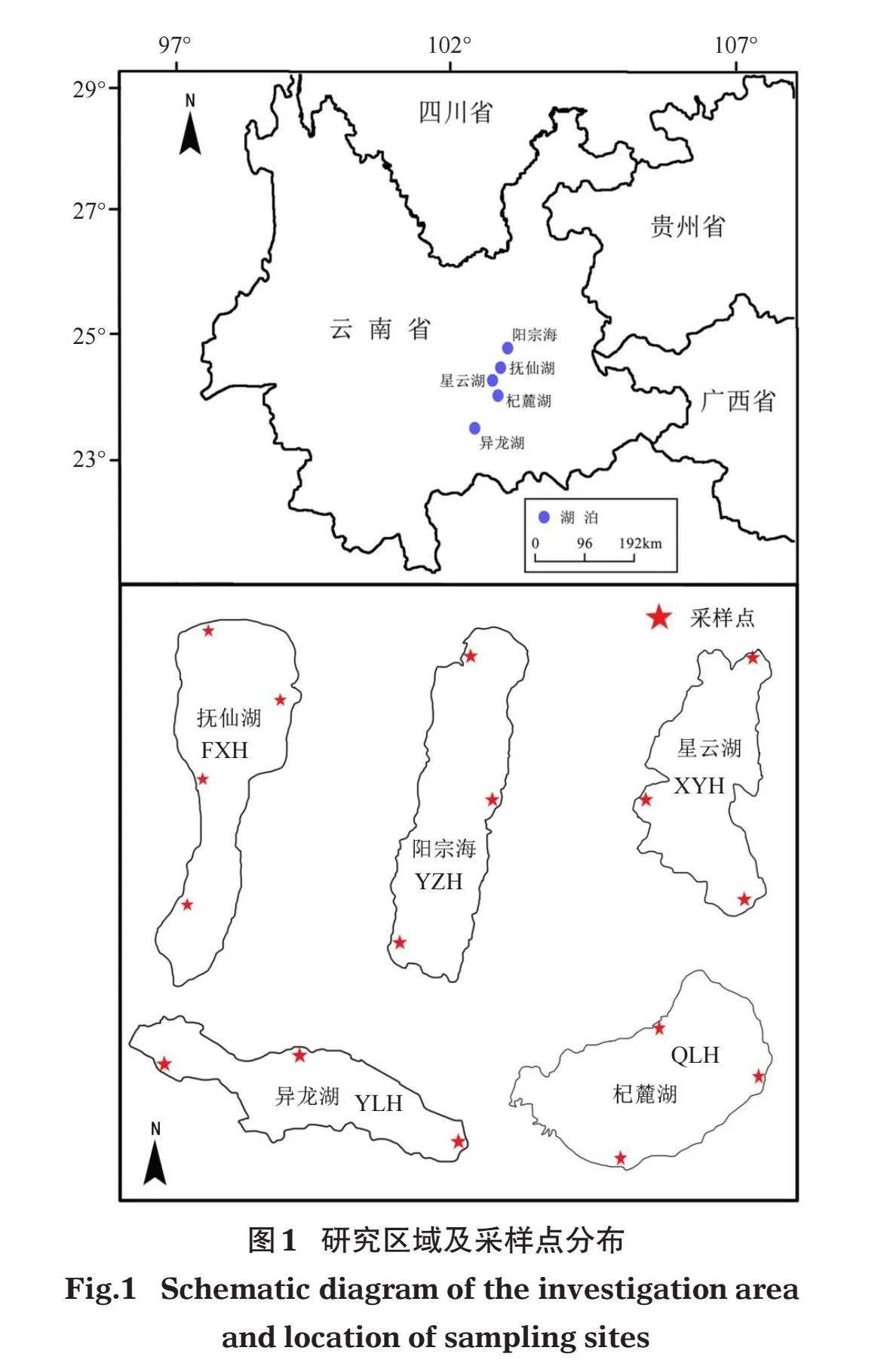
分別于2020年和2021年9-10月對云南中部5個典型高原湖泊進行2個年度的沉水植物調查(圖1)。根據所選湖泊大小、受干擾程度以及沉水植物實際分布情況設置采樣點,對沉水植物的群落組成及水環境因子進行調查。其中,撫仙湖(FXH)和陽宗海(YZH)沉水植物主要局限于環湖湖岸帶一周,因此調查時沿湖岸帶設置采樣點(沈亞強等,2010;高弋明等,2021)。星云湖(XYH)、杞麓湖(QLH)和異龍湖(YLH)由于水生態環境較差,沉水植物分布面積和生物量已大為減少,并呈現向岸邊萎縮和分布不均勻的趨勢(沈亞強等,2010),同樣采用環湖設置采樣點的方法進行調查。其中,撫仙湖設置4個采樣點,星云湖、杞麓湖、陽宗海和異龍湖各設置3個采樣點。各湖泊的基本形態學特征見表1。
1.2" "環境因子的測定
現場采用透明度盤測定透明度(SD),用YSI多參數水質監測儀測定水體水溫(WT)、pH、電導率(SPC)、氧化還原電位(ORP)、溶解氧(DO)、鹽度(Sal)等參數,采集表層水樣帶回實驗室分析總氮(TN)、總磷(TP)、氨氮(NH4+-N)、硝氮(NO3--N)、磷酸鹽(PO43-)、葉綠素a(Chl-a)和高錳酸鹽指數(CODMn),水體化學指標的測定參考《水和廢水監測分析方法》(國家環境保護總局,2002)。
1.3" "沉水植物的采集及處理
深水區使用0.5 m×0.5 cm的采草器隨機采集3次;岸邊淺水區使用面積為 0.5 m×0.5 m 的鋁制樣方框確定采樣區域,徒手采集樣方框內所有植物,采集3次。采集到的沉水植物及時沖洗干凈、分類、稱重,以濕重代表生物量。
以往研究多使用植物株數密度來計算物種多樣性指數,但沉水植物由于其特殊的生物學特性,絕大部分種類多通過分枝或分蘗的方式進行繁殖,而不是簡單地通過增加株數來擴大種群生物量。因此,本文參考Jeppesen等(2000)的方法,使用生物量來計算沉水植物的優勢度和多樣性指數,即用相對生物量(RB)與相對頻度(RF)來計算物種的優勢度(DV):
DV=(RB + RF)/2×100%" " " ①
用香農-威納多樣性指數(H')計算公式如下:
H' = -[i=1Sbib lnbib]" " " " "②
式中:bi為群落中i物種生物量;b為群落中所有沉水植物的生物量之和;S為該群落總物種數。
1.4" "湖泊營養程度評價方法
采用綜合營養指數(TLI)法評價湖泊富營養化(金相燦,1995;王明翠等,2002)。計算公式為:
TLI(Chl-a)=10(2.5+1.086 ln Chl-a)" "③
TLI(TP)=10(9.436+1.624 ln TP)" " "④
TLI(TN)=10(5.453+1.694 ln TN)" " ⑤
TLI(SD)=10(5.118-1.94 ln SD)" " ⑥
TLI(∑)=W(Chl-a)·TLI(Chl-a)+W(TP)·TLI(TP)+W(TN)·TLI(TN)+W(SD)·TLI(SD)" " " " "⑦
式中:W(Chl-a)、W(TP)、W(TN)和W(SD)分別為0.3260、0.2300、0.2192和0.2246。采用0~100的一系列連續數字對湖泊營養狀態進行分級:TLI(∑)≤30為貧營養;30lt;TLI(∑)≤50為中營養;50lt;TLI(∑)≤60為輕度富營養;60lt;TLI(∑)≤70為中度富營養;70lt;TLI(∑)≤100為高度富營養。
1.5nbsp; "數據分析
利用One-way ANOVA對不同湖泊環境因子變量均值進行分析(Plt;0.05),在進行方差分析前,對數據進行了方差齊性分析和正態分布分析,若方差齊性,選擇LSD檢驗,反之使用Tamhane's檢驗(Plt;0.05),用SPSS19.0軟件進行分析,origin 9.0進行作圖。運用R 4.1.3中的vegan包分析沉水植物優勢度與環境因子的關系。在數據進行排序分析前,對物種數據和環境因子數據進行標準化處理(ErO\"s et al,2012),再進行去趨勢對應分析(detrended correspondence analysis, DCA)以確定排序模型(Braak amp; Smilauer,2002)。采用方差膨脹因子(variance inflation factor,VIF)分析,剔除共線性強的環境因子(VIFgt;10);Pearson相關分析進一步剔除強相關性環境因子(rgt;0.7);采用蒙特卡洛檢測方法檢驗環境因子對植物影響的顯著性,保留Plt;0.05的環境因子(Blanchet et al,2008)。
2" "結果與分析
2.1" "水環境現狀
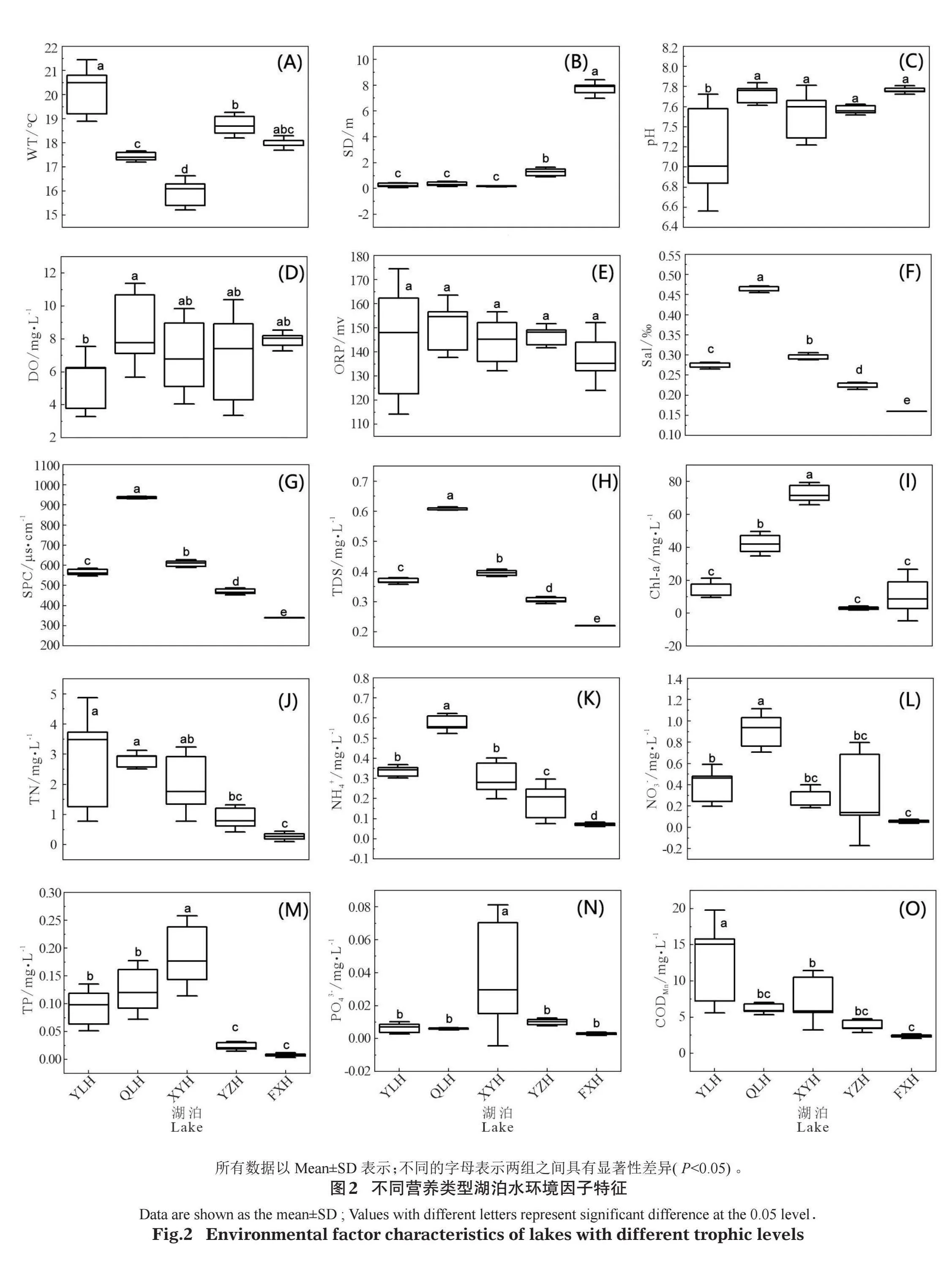
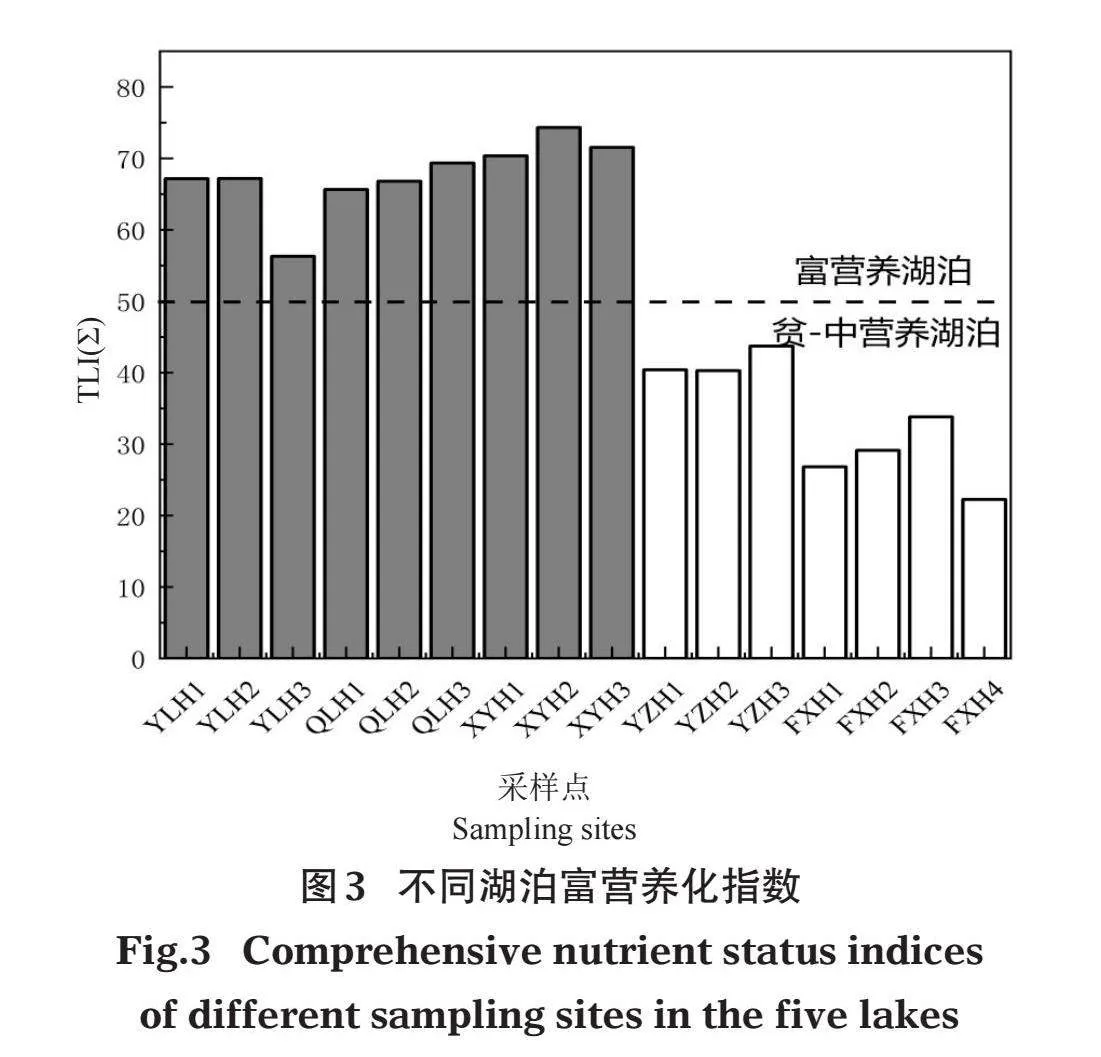
不同營養類型湖泊水環境因子特征如圖2。從中可以看出,異龍湖、杞麓湖、星云湖和陽宗海4個湖泊的WT差異顯著(Plt;0.05),異龍湖最高,星云湖最低,撫仙湖與異龍湖WT具有顯著差異(Plt;0.05),其他湖泊差異不顯著(圖2A)。Sal、SPC、TDS 3個指標在不同湖泊間具有顯著差異(Plt;0.05),表現為杞麓湖gt;星云湖gt;異龍湖gt;陽宗海gt;撫仙湖(圖2F、2G、2H)。陽宗海和撫仙湖的SD顯著大于杞麓湖、星云湖和異龍湖(P<0.05)(圖2B)。杞麓湖、星云湖和異龍湖TN、NH4+-N、TP顯著大于撫仙湖(Plt;0.05)(圖2J、2K、2M),pH、DO、ORP、PO43-、CODMn等水環境因子在各湖泊之間差異不大(圖2C、2D、2E、2N、2O)。綜合營養狀態指數法計算表明,異龍湖、杞麓湖和星云湖屬于富營養湖泊,營養指數分別為63.55±6.28、67.26±1.89和72.08±2.04;撫仙湖和陽宗海屬于貧-中營養湖泊,營養指數分別為28.01±4.82和41.47±1.96(圖3)。
2.2" "沉水植物群落現狀
本次共調查到沉水植物6科10屬15種,包括苦草(Vallisneria natans)、大茨藻(Najas marina)、篦齒眼子菜(Potamogeton pectinatus)、穿葉眼子菜(P. perfoliatus)、光葉眼子菜(P. lucens)、角果藻(Pteridophyta)、金魚藻(Ceratophyllum demersum)、麗藻屬(Nitella)、輪葉黑藻(Hydrilla verticillata)、輪藻屬(Chara)、馬來眼子菜(P. wrightii)、穗花狐尾藻(Myriophyllum spicatum)、微齒眼子菜(P. maackianus)、伊樂藻(Elodea nuttallii)和菹草(P. crispus)。
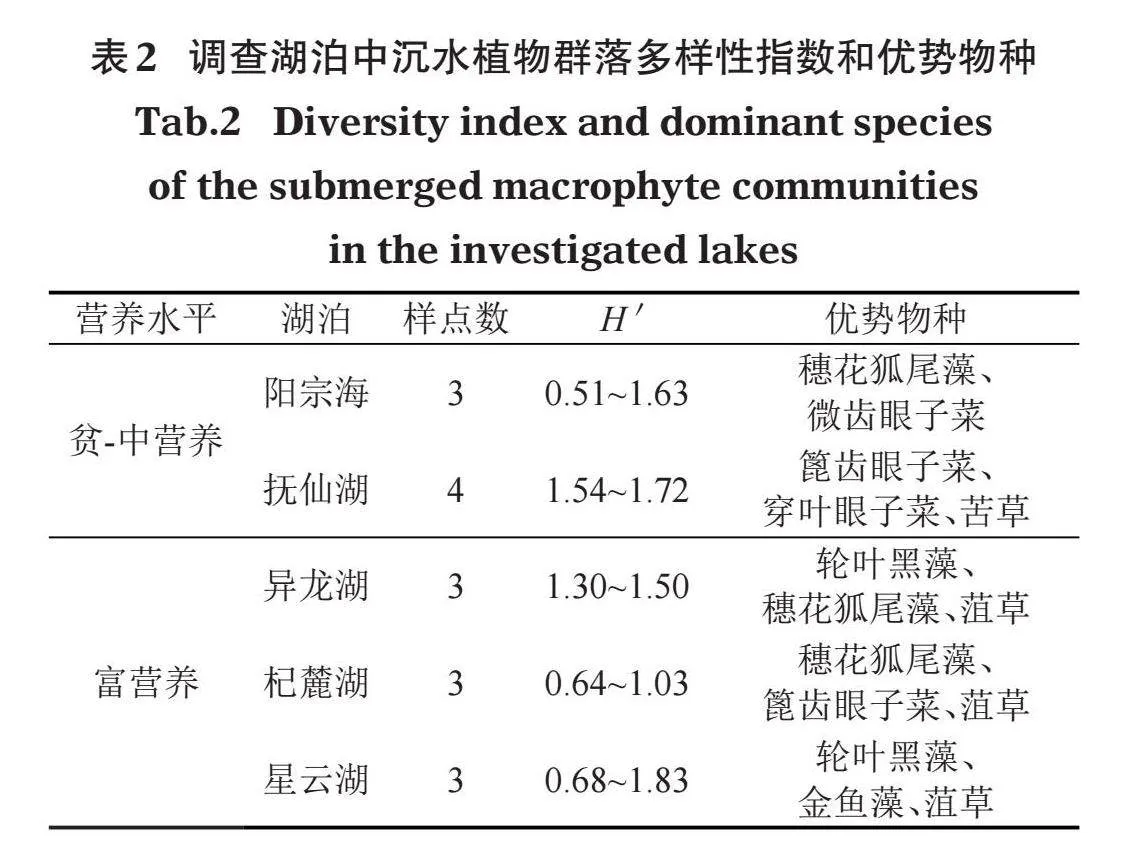
云南高原湖泊中沉水植物的主要優勢種為穗花狐尾藻、微齒眼子菜、輪葉黑藻、穿葉眼子菜、菹草、篦齒眼子菜和苦草。不同營養類型湖泊沉水植物優勢種有所不同,微齒眼子菜和穿葉眼子菜為貧-中營養湖泊的優勢種,穗花狐尾藻和篦齒眼子菜在各營養類型湖泊中均能成為優勢種或亞優勢種,輪葉黑藻、菹草和金魚藻僅在富營養湖泊中成為優勢種(表2)。沉水植物多樣性分析表明,撫仙湖的香農-威納多樣性H'最高,平均值達到1.63;杞麓湖的H'最低,平均值僅0.79。總體來說,貧-中營養湖泊的沉水植物多樣性要大于富營養湖泊。在16個樣點中,出現頻率最高的是穗花狐尾藻,為87.5%;最低是大茨藻、角果藻和麗藻屬,為6.25%(圖4)。
2.3" "沉水植物群落物種與環境因子的關系
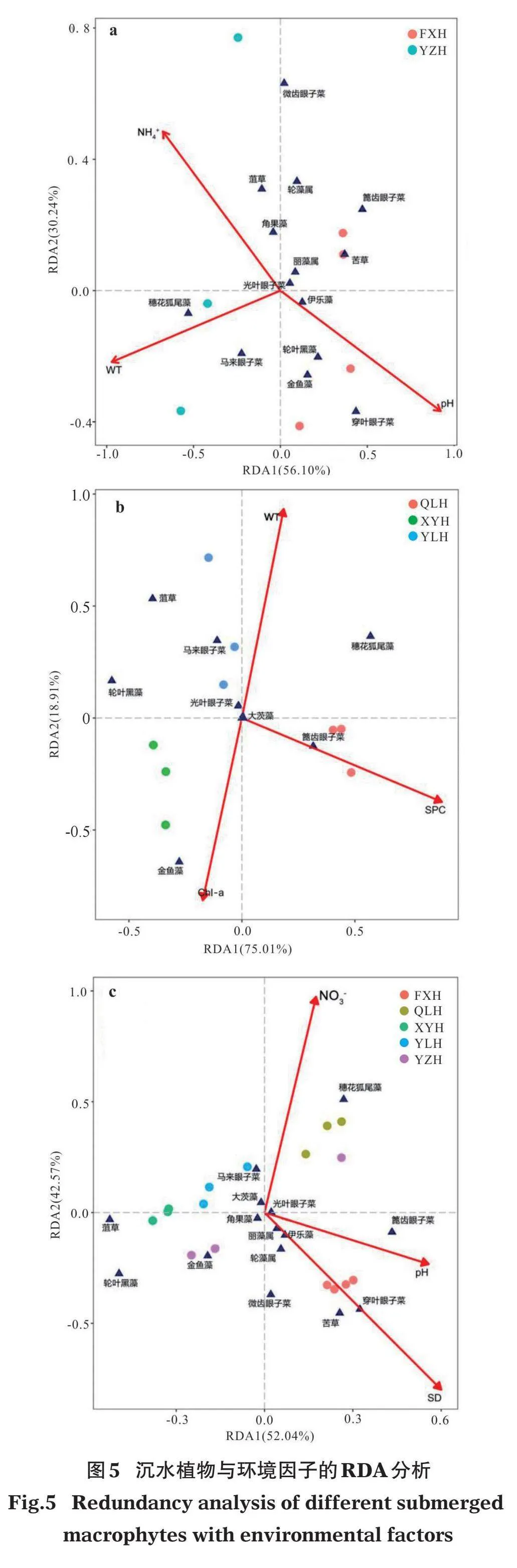
DCA結果表明,各排序軸的最大梯度長度(LOG)為:貧-中營養湖泊1.87、富營養湖泊3.67、整體湖泊2.99,都lt;4,符合線性模型,因此本研究統一選用RDA分析方法。分析顯示,貧-中營養湖泊、富營養湖泊、整體湖泊的第1、2軸共解釋物種與環境因子之間累計方差為86.34%、93.92%、94.61%,說明排序結果可以很好地解釋沉水植物群落組成與環境因子的關系(圖5)。
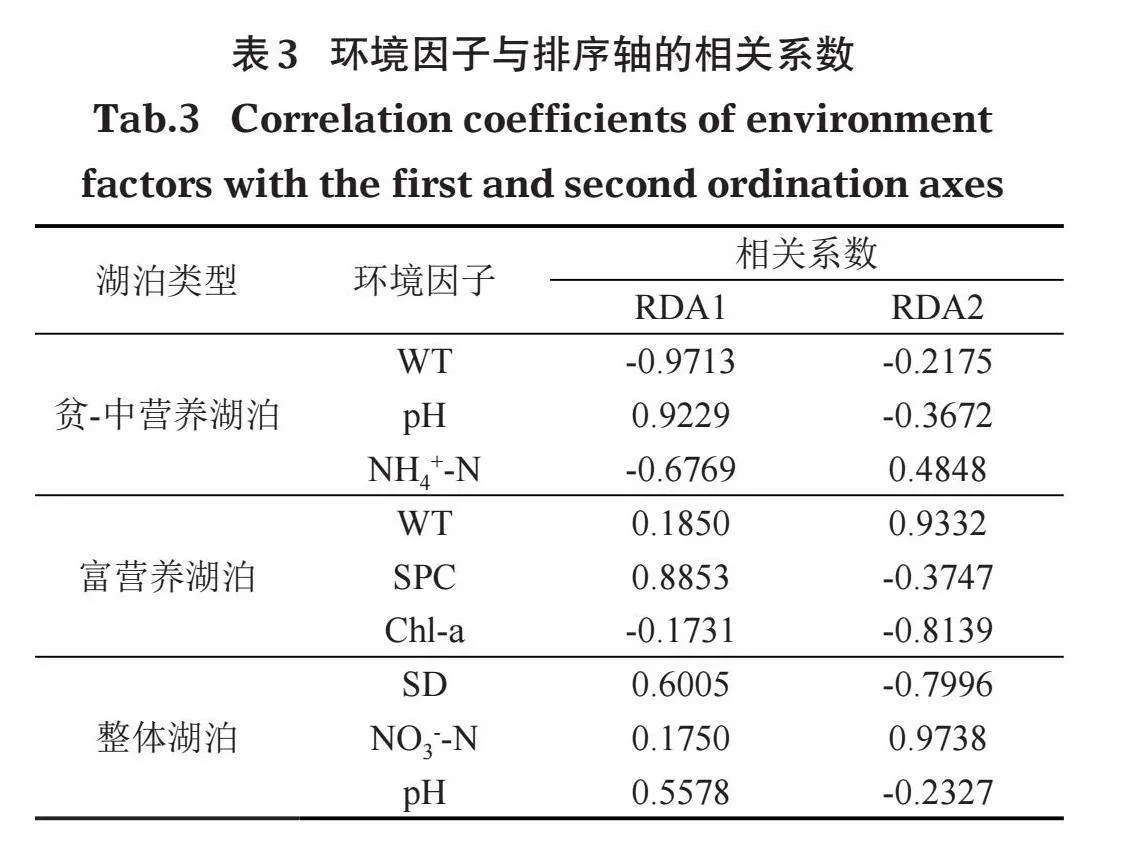
蒙特卡洛置換檢驗分析表明,貧-中營養湖泊中15個環境變量中只有3個變量對解釋沉水植物群落組成與環境因子的關系做出了顯著貢獻(即Plt; 0.05)。其中,穿葉眼子菜、輪葉黑藻、金魚藻與pH呈顯著正相關,與NH4+-N呈顯著負相關;菹草、角果藻、輪藻屬和微齒眼子菜與NH4+-N呈顯著正相關,與pH呈顯著負相關;馬來眼子菜和穗花狐尾藻與WT呈顯著正相關,篦齒眼子菜、苦草和輪藻屬與WT呈顯著負相關(圖5a)。環境因子與坐標軸的相關系數越高則表明環境因子對物種的相關性越強,影響越大,由表3可以看出,pH、NH4+-N、WT是影響貧-中營養湖泊沉水植物分布特征的主要因子。
富營養湖泊RDA分析中,通過蒙特卡洛置換檢驗預選出符合條件的環境因子為WT、SPC、Chl-a,其中,輪葉黑藻、菹草和馬來眼子菜與SPC呈顯著負相關,穗花狐尾藻和篦齒眼子菜與SPC呈顯著正相關;金魚藻與Chl-a呈顯著正相關,與WT呈顯著負相關;穗花狐尾藻、馬來眼子菜、菹草與WT呈顯著正相關,與Chl-a呈顯著負相關(圖5b)。環境因子與坐標軸的相關系數表明,WT、SPC、Chl-a是影響富營養湖泊沉水植物分布特征的主要因子(表3)。
整體湖泊RDA分析表明,SD、NO3--N、pH為蒙特卡洛置換檢驗預選出的環境因子。其中,穿葉眼子菜、苦草、篦齒眼子菜、微齒眼子菜、輪藻屬、伊樂藻和麗藻屬分別與pH和SD呈顯著正相關;菹草、輪葉黑藻、金魚藻以及馬來眼子菜分別與pH和SD呈顯著負相關;穗花狐尾藻和馬來眼子菜與NO3--N呈顯著正相關;穿葉眼子菜、苦草、篦齒眼子菜、微齒眼子菜、輪藻屬、伊樂藻、麗藻屬、金魚藻和輪葉黑藻都與NO3--N呈顯著負相關(圖5c)。環境因子與坐標軸的相關系數表明,SD、NO3--N、pH是影響整體湖泊沉水植物分布特征的主要因子(表3)。
3" "討論
3.1" "沉水植物群落組成變化
本研究通過綜合營養狀態指數法計算表明,異龍湖、杞麓湖和星云湖屬于富營養湖泊,撫仙湖和陽宗海屬于貧-中營養湖泊(圖3)。調查共發現沉水植物6科10屬15種,其中優勢種包括穗花狐尾藻、微齒眼子菜、篦齒眼子菜、穿葉眼子菜、輪葉黑藻、菹草和苦草。微齒眼子菜和穿葉眼子菜在貧-中營養湖泊中可以成為優勢種,穗花狐尾藻和篦齒眼子菜在各營養湖泊中均能成為優勢種或亞優勢種,輪葉黑藻、菹草和金魚藻僅能在富營養湖泊中成為優勢種(表2)。導致這種分布現象的原因可能包括:(1)環境因素。已有研究表明水生植物的生長受水深、水溫、光照強度、營養鹽、水文水動力條件、底質類型等眾多環境因子共同作用,水生生境條件對水生植物的物種組成及分布具有顯著的影響(秦伯強,2002)。(2)人為因素。社會、經濟、工農業的發展以及長期以來對湖泊資源的不合理開發,造成云南地區湖泊生態環境嚴重破壞,湖泊富營養化嚴重,湖泊生態系統日益退化(顏昌宙等,2005)。
已有研究發現,異龍湖水環境在21世紀90年代后逐漸惡化,2009年異龍湖水質類別甚至達到劣Ⅴ類,水生生態系統發生劇烈變化,平均透明度為0.22 m,綜合營養狀態指數為78.13,同年進行的該湖泊水生植物調查顯示沉水植物已完全消失(魏翔和唐光明,2014)。本研究調查異龍湖的綜合營養狀態指數為63.55±6.28,仍屬于中度富營養狀態,沉水植物雖有所恢復,但多以耐污種(穗花狐尾藻和篦齒眼子菜)為主,這與方馨等(2021)的研究結果一致。1970年代以來,受周邊圍湖造田以及農業面源污染的長期影響,杞麓湖水生生態系統遭到嚴重破壞,對水質要求非常嚴格的海菜花(Ottelia acuminata)、水鱉(Hydrocharis dubia)群落、荇菜(Nymphoides peltatum)群落等已相繼消失,對水質要求較高的竹葉眼子菜群落分布面積巨減,穗花狐尾藻、菹草等耐污種群落逐漸增多,成為優勢類群(沈亞強等,2010),本研究調查也發現穗花狐尾藻、篦齒眼子菜、菹草為杞麓湖的主要優勢種。已有資料顯示,隨著星云湖周邊畜禽養殖業生產規模的擴大,畜禽糞便污染已成為星云湖水環境的重要的污染源(邱成和葉春麗,2015)。另有研究認為,星云湖周邊農田施用的化學用劑污染及大量食草魚類放養等因素影響其水質質量,導致湖內植物群落類型單一,僅有少數對環境要求較低的植物得以生存(李恒,1980)。本次調查發現星云湖綜合營養狀態指數為72.08±2.04,屬于高度富營養狀態,間接導致該湖泊沉水植物生物多樣性較低,輪葉黑藻、金魚藻、菹草等耐污種為主要優勢種(表2),這與楊加林等(2012)的研究結果一致。
3.2" "沉水植物與環境因子的關系
本研究結果發現,在水體營養鹽水平差異不大的情況下(中營養湖泊和富營養湖泊),WT成為影響沉水植物分布的主要因子(圖5a和圖5b),且在富營養湖泊中,沉水植物物種多樣性隨著溫度的升高而增加(表2)。已有研究表明,水溫和沉積物溫度都會影響水生植物分布及其群落結構特征(Rooney amp; Kalff,2000;Spencer et al,2000;Heino,2002)。溫度可對水體鹽度、營養鹽含量和氣體含量產生明顯的影響,在水生態系統的物理、化學和生物過程中發揮著重要作用(Welch,1952)。較高的水溫會增加水體的蒸發量,降低水體溶解氧,從而增加許多水生物種的含鹽量,增加其生存壓力(Al-Thahaibawi et al,2021)。本研究發現富營養湖泊異龍湖,其較高的水溫導致了較低的水體溶解氧和鹽度(圖2),這也是導致該湖泊沉水植物物種多以耐污種為主的原因。
SPC是影響富營養湖泊沉水植物分布的另一主要環境因子(圖5b)。水體SPC的變化與水位、蒸發量都有顯著的關系(Al-Kenzawi et al,2011),此外,農業活動面源污染也可能導致夏季水體SPC過高(Al-Shawi,2006)。已有研究表明,自1970年代以來,杞麓湖一直受周邊圍湖造田和農業面源污染的影響,使得竹葉眼子菜群落分布面積銳減,穗花狐尾藻、菹草群落等耐污種逐漸蔓延(沈亞強等,2010;邱成和葉春麗,2015)。本研究調查發現,杞麓湖水體SPC顯著大于其他4個湖泊(Plt;0.05)(圖2),耐鹽能力較強的穗花狐尾藻(劉萌萌等,2019)、篦齒眼子菜(王衛紅和季民,2007)為主要優勢物種。
pH是沉水植物生存與繁衍的重要環境因子,其通過改變水體溶解無機碳的平衡關系,進而影響水生植物的生長和分布(任南等,1996;何俊等,2008)。已有研究表明,穗花狐尾藻、篦齒眼子菜在pH值為7.5~8.8時對HCO3-形式的碳源都表現出較強的利用能力,從而能適應高pH的水環境(蘇睿麗和李偉,2005;劉玲玲,2011)。本次調查的5個云南高原湖泊的pH值平均范圍為7.14~7.77,穗花狐尾藻和篦齒眼子菜為主要優勢種和亞優勢種,也證實了這一結論。以往研究還發現,當pH為7~9時,輪葉黑藻更易在富營養狀態水體中生長(張瀏等,2007),本次調查結果與其一致,即在富營養湖泊(異龍湖、星云湖)中輪葉黑藻更易成為優勢類群。
由于沉水植物整個生活史大部分位于水中,底質和水體中氮元素可直接被其根系和葉片吸收,其中NH4+-N和NO3--N兩類基礎無機氮源最易被吸收利用(段娜等,2015)。本研究中湖泊NH4+-N和NO3-的平均濃度為0.28 mg/L和0.37 mg/L,其中富營養湖泊NH4+-N為0.40 mg/L、NO3--N為0.53 mg/L,貧-中營養湖泊NH4+-N為0.12 mg/L、NO3--N為0.17 mg/L。已有研究表明,在一定范圍內,較高的氮素濃度有利于馬來眼子菜的生長(蘇睿麗和李偉,2005)。本研究發現馬來眼子菜與NO3--N濃度呈顯著正相關(圖5c),其在富營養湖泊異龍湖中更容易成為優勢物種。
水下光照條件是沉水植物進行正常光合作用的重要前提,是影響沉水植物在天然水體中生長的主要因素(王華等,2008)。已有研究表明,沉水植物可正常生長的水深通常為水體SD的2~2.5倍(邱成和葉春麗,2015)。本研究發現,水體SD是影響云南高原湖泊沉水植物分布最關鍵的環境因子(圖5c);貧-中營養湖泊(撫仙湖和陽宗海)的SD要顯著大于富營養湖泊(Plt;0.05),其中撫仙湖的SD為(7.77±0.03)m(圖2)。由于沉水植物對光和空間的競爭能力不同,可粗分為草甸型植物和冠層型植物2類(James et al,2004)。其中,草甸型植物具有莖或分枝短、植冠低矮、生長較慢的生物學特征,絕大部分生活在沉積物附近,對光和生存空間具有相對較弱的競爭能力,如輪藻類、苦草、篦齒眼子菜和微齒眼子菜等物種;相反,冠層型植物具有較長的莖或分枝,其植冠高大、生長迅速,可生長到水面附近,對光和空間的具有較強的競爭能力,如穗花狐尾藻、黑藻和金魚藻等物種(Gopal amp; Usha,1993)。由于撫仙湖、陽宗海具有較高的SD,故苦草、篦齒眼子菜、穿葉眼子菜和微齒眼子菜等草甸型沉水植物可以在該類型湖泊中成為優勢種。
沉水植物的生長分布與水環境因子是相互影響的。草甸型植物比冠層型植物更有利于維持湖泊的清水穩態(Carpenter amp; Lodge,1986;Rehman et al,2016)。草甸型沉水植物多具有相對稠密的須根,且絕大部分生活在沉積物附近,能有效阻止沉積物向水體釋放營養鹽,顯著提升沉積物滯留營養鹽的能力(Kufel amp; Kufel,2002;James et al,2004)。冠層型沉水植物生長較快,其衰敗腐爛時會釋放大量腐殖質和營養鹽進入水體,導致浮游植物快速繁殖,引起透明度下降(Engelhardt amp; Ritchie,2001;Hilt et al,2006)。因此,當沉水植物類型的轉變,也預示著湖泊營養水平的轉變(da Silva et al,2014)。另外,水環境因子之間也是相互影響、相互制約的,單個環境因子可以通過影響其他環境因子進而影響沉水植物的生長和分布。因此,在研究影響水生植物生長分布的環境因子時,應當綜合考慮各環境因子之間的內在聯系,全面系統分析水生植物與環境因子的關系。
綜合本次調查研究結果,提出以下建議:(1)針對類似本研究的貧-中營養類型湖泊,要遵循沉水植物的生物學特征和生長規律,及時收割清除高大的冠層型沉水植物,創造草甸型沉水植物的生長條件,促進沉水植物群落向貧營養化湖泊的群落結構方向發展;(2)針對類似本研究的富營養類型湖泊,其沉水植物恢復的關鍵要控制水體營養鹽濃度,抑制藻類的生長,采用不同的恢復方法,選取不同的植物種類來制定沉水植物的恢復方案。
參考文獻
段娜, 章堯想, 劉芳, 等, 2015. 植物氮素吸收及其轉運蛋白研究進展[J].分子植物育種, 13(2): 461-468.
方馨, 趙風斌, 柴曉利, 等, 2021. 異龍湖沉水植物分布格局與水環境因子相關性研究[J].長江流域資源與環境," 30(3): 636-643.
國家環境保護總局," 2002. 水和廢水監測分析方法[M].4版. 北京: 中國環境科學出版社.
高弋明, 殷春雨, 劉霞, 等, 2021. 撫仙湖近60年來沉水植物群落變化趨勢分析[J].湖泊科學, 33(4): 1209-1219.
何俊, 谷孝鴻, 劉國鋒, 2008. 東太湖水生植物及其與環境的相互作用[J]. 湖泊科學, 20(6): 47-49.
金相燦, 1995. 中國湖泊環境[M]. 北京: 海洋出版社.
金相燦, 楚建周, 王圣瑞, 2007. 水體氮濃度、形態對黑藻和狐尾藻光合特征的影響[J].應用與環境生物學報, 13(2): 200-204.
孔祥虹, 肖蘭蘭, 蘇豪杰, 等, 2015. 長江下游湖泊水生植物現狀及與水環境因子的關系[J].湖泊科學, 27(3): 385-391.
藍紅林, 2001. 云南高原湖泊的治理與保護初探[J].云南環境科學, 20(4): 26-28.
李恒, 1980. 云南高原湖泊水生植被的研究[J].云南植物研究, 2(2): 113-141.
李文朝, 連光華, 1996. 幾種沉水植物營養繁殖體萌發的光需求研究[J]. 湖泊科學," 8(增刊1): 25-29.
劉玲玲, 2011. 三種沉水植物無機碳利用機制研究[D].武漢:華中師范大學.
劉萌萌, 劉巧," 楊娜," 等, 2019. 沉水植物穗花狐尾藻耐鹽性與生長[J].生態學雜志, 38(3): 778-784.
秦伯強, 2002. 長江中下游淺水湖泊富營養化發生機制與控制途徑初探[J].湖泊科學, 14(3): 193-202.
邱成, 葉春麗, 2015. 淺析江川縣發展生態畜牧業與星云湖水環境保護[C]. 中國環境科學學會環境經濟學分會2015年學術年會.
任南, 嚴國安, 馬劍敏, 等, 1996. 環境因子對東湖幾種沉水植物生理的影響研究[J].武漢大學學報(自然科學版), 42(2): 213-218.
沈亞強, 王海軍, 劉學勤, 2010. 滇中五湖水生植物區系及沉水植物群落特征[J]. 長江流域資源與環境, 19(增刊1): 111-119.
蘇睿麗, 李偉, 2005. 沉水植物光合作用的特點與研究進展[J].植物學通報, 22(增刊1): 128-138.
王華, 逄勇, 劉申寶, 等, 2008. 沉水植物生長影響因子研究進展[J].生態學報," 28(8): 3958-3968.
王明翠, 劉雪芹, 張建輝, 2002. 湖泊富營養化評價方法及分級標準[J].中國環境監測, 18(5): 47-49.
王衛紅, 季民, 2007. 9種沉水植物的耐鹽性比較[J].農業環境科學學報, 26(4): 1259-1263.
王文林, 王國祥, 李強, 等, 2006. 水體濁度對菹草(Potamogeton cripus)幼苗生長發育的影響[J].生態學報, 26(11): 3586-3593.
魏翔, 唐光明, 2014. 異龍湖近20年來營養鹽與水生生態系統變化[J].環境科學導刊, 33(2): 9-14.
吳愛平, 吳世凱, 倪樂意, 2005. 長江中游淺水湖泊水生植物氮磷含量與水柱營養的關系[J].水生生物學報," 29(4): 406-412.
謝貽發, 李傳紅, 劉正文, 等, 2007. 基質條件對苦草(Vallisneria natans)生長和形態特征的影響[J].農業環境科學學報, 26(4): 1269-1272.
楊加林, 李杰, 李經緯, 等, 2012. 撫仙湖-星云湖水生生物與水環境研究綜述[J].云南地理環境研究, 24(2): 98-102.
楊柳, 黃耀志, 2002. 云南高原湖區綜合發展研究[J].重慶大學學報(社會科學版)," 8(3): 19-21.
顏昌宙, 金相燦, 趙景柱, 等, 2005. 云南洱海的生態保護及可持續利用對策[J]. 環境科學, 26(5): 38-42.
閆志強, 劉黽, 吳小業, 等, 2014. 溫度對五種沉水植物生長和營養去除效果的影響[J].生態科學, 33(5): 839-844.
游灝, 2006. 五種沉水植物對富營養化水體的生態適應性研究[D].南京:南京農業大學.
張瀏, 陳燦, 高倩, 等," 2007. 兩種營養狀態下pH對輪葉黑藻(Hydrilla verticillata)生長和抗氧化酶活性的影響[J].生態環境, 16(3): 748-752.
曾理, 吳豐昌, 萬國江, 等, 2009. 中國地區湖泊沉積物中137Cs分布特征和環境意義[J].湖泊科學, 21(1): 1-9.
Al-Kenzawi M," Al-Haidary M," Talib A H," et al," 2011. Environmental study of some water characteristics at Um-Al-Naaj Marsh," south of Iraq[J]. Baghdad Science Journal," 8(1): 531-538.
Al-Thahaibawi B," Al-Abbawy D," Al-Mayaly I," et al," 2021. The effect of some environmental variables on the abundance and distribution of aquatic macrophytes in Al-Hawizeh marsh," Iraq[J]. IOP Conference Series: Earth and Environmental Science," 722(1): 012001.
Al-Shawi I," 2006. Comparative study of some physico-chemical characteristics for Northern Al-Hammar marsh waters before destroyed and after Rehabilitation 2004[J]. Marsh Bulletin," 2:127-133.
Barko J W," Smart R M," 1981. Comparative influences of light and temperature on the growth and metabolism of selected submersed freshwater macrophytes[J]. Ecological Monographs," 51(2): 219-235.
Blanchet F G," Legendre P," Borcard D," 2008. Forward selection of explanatory variables[J]. Ecology," 89(9): 2623-2632.
Bornette G," Puijalon S," 2011. Response of aquatic plants to abiotic factors: a review[J]. Aquatic Sciences," 73(1): 1-14.
Carpenter S R," Lodge D M," 1986. Effects of submersed macrophytes on ecosystem processes[J]. Aquatic Botany," 26: 341-370.
da Silva S C A D," Cervi A C," Bona C," et al," 2014. Aquatic macrophyte community varies in urban reservoirs with different degrees of eutrophication[J]. Acta Limnol Bras," 26(2): 129-142.
Engelhardt K," Ritchie M E," 2001. Effects of macrophyte species richness on wetland ecosystem functioning and services[J]. Nature," 411(6838): 687-689.
EeO\"s T," S[a][']ly P," Tak[a][']cs P," et al," 2012. Temporal variability in the spatial and environmental determinants of functional metacommunity organization-stream fish in a human-modified landscape[J]. Freshwater Biology," 57(9): 1914-1928.
Gopal B," Usha G," 1993. Competition and allelopathy in aquatic plant communities[J]. The Botanical Review," 59(3): 155-210.
Heino J," 2002. Concordance of species richness patterns among multiple freshwater taxa: a regional perspective[J]. Biodiversity amp; Conservation," 11(1):137-147.
Hilt S," Gross E," Hupfer M," et al," 2006. Restoration of submerged vegetation in shallow eutrophic lakes: a guideline and state of the art in Germany[J]. Limnologica- Ecology and Management of Inland Waters," 36(3): 155-171.
Horppila J," Nurminen L," 2005. Effects of different macrophyte growth forms on sediment and P resuspension in a shallow lake[J]. Hydrobiologia," 545(1): 167-175.
James W F," Barko J W," Butler M G," 2004. Shear stress and sediment resuspension in relation to submersed macrophyte biomass[J]. Hydrobiologia," 515(1/2/3): 181-191.
Jeppesen E," Jensen J P," M S?ndergaard," et al," 2000. Trophic structure," species richness and biodiversity in Danish lakes: changes along a phosphorus gradient[J]. Freshwater Biology," 45(2): 201-218.
Kalff Jacob," 2002. Limnology: Inland Water Ecosystems[M].Prentice Hall," NJ: Benjamin Cummings.
Kufel L," Kufel I," 2002. Chara beds acting as nutrient sinks in shallow lakes: a review[J]. Aquatic Botany," 72(3/4): 249-260.
Wang H," Wang Q," Bowler P A," et al," 2016. Invasive aquatic plants in China[J]. Aquatic Invasions," 11(1): 1-9.
Nakamura K," Kayaba Y," Nishihiro J," et al," 2008. Effects of submerged plants on water quality and biota in large-scale experimental ponds[J]. Landscape amp; Ecological Engineering," 4(1): 1-9.
Rehman F," Pervez A," Khattak B N," et al," 2016. Constructed wetlands: perspectives of the oxygen released in the rhizosphere of macrophytes[J]. Clean-Soil," Air," Water," 45(1).DOI:10.1002/clen.201600054.
Rooney N," Kalff J," 2000. Inter-annual variation in submerged macrophyte community biomass and distribution: the influence of temperature and lake morphometry[J]. Aquatic Botany," 68(4): 321-335.
Sachse R," Petzoldt T," Blumstock M," et al," 2014. Extending one-dimensional models for deep lakes to simulate the impact of submerged macrophytes on water quality[J]. Environmental Modelling amp; Software," 61: 410-423.
Spencer D F," Ksander G G," Madsen J D," et al," 2000. Emergence of vegetative propagules of Potamogeton nodosus," Potamogeton pectinatus," Vallisneria americana," and Hydrilla verticillata based on accumulated degree-days[J]. Aquatic Botany," 67(3): 237-249.
Welch P S," 1952. Limnology[M]. New York: McGraw Hill Book Company.
(責任編輯" "鄭金秀)
Community Composition of Submerged Macrophytes and Relationship
with Environmental Factors in Five Yunnan Plateau Lakes
ZHU Li‐ming, ZHANG Dao‐xi, SHI Fang, ZOU Xi, ZHANG Zhi‐yong, HU Lian
(Key Laboratory of Ecological Impacts of Hydraulic-Projects and Restoration of Aquatic Ecosystem
of Ministry of Water Resources, Institute of Hydroecology, Ministry of Water Resources amp; Chinese
Academy of Sciences, Wuhan" "430079, P.R. China)
Abstract:Many studies have suggested that restoration of the submerged plant community is an important means for controlling lake eutrophication. Therefore, it is important to study the composition and distribution of the submerged plant communities in lakes and the relationship with the water environment factors. In this study, five typical plateau lakes in central Yunnan Province were selected for research, including Yilong Lake (YLH), Qilu Lake (QLH), Xingyun Lake (XYH), Fuxian Lake (FXH) and Yangzonghai lake (YZH). We investigated the composition and distribution of submerged macrophytes communities in the five lakes, and the relationship between the distribution of submerged macrophytes and water environmental factors. A two-year investigation of submerged plants and water environment factors were conducted in the five lakes during September and October of 2020 and 2021. The comprehensive nutrient status indices of Yilong Lake, Qilu Lake and Xingyun Lake were 63.55±6.28, 67.26±1.89 and 72.08±2.04, respectively, indicating an eutrophic state, and the comprehensive nutrient status indices of Fuxian Lake (FXH) and Yangzonghai Lake (YZH) were 28.01±4.82 and 41.47±1.96, respectively, indicating an oligotrophic-mesotrophic state. A total of 15 submerged macrophyte species belonging to 6 families and 10 genera were identified during the investigation. Myriophyllum spicatum, P. pectinatus were dominant species in all lakes, Potamogeton maackianus and P. perfoliatus were dominant in the oligotrophic-mesotrophic lakes, and Hydrilla verticillata, P. crispus and Ceratophyllum demersum were dominant in the eutrophic lakes. The Shannon-Weiner diversity (H') index of the submerged macrophytes in FXH was highest, with an average value of 1.63, while the diversity index of QLH was lowest, with an average value of only 0.79. In general, submerged macrophyte diversity was higher in the oligotrophic-mesotrophic lakes than in the eutrophic lakes. Redundancy analysis of the submerged plants and the environmental factors shows that transparency, pH and NO3--N were the key factors affecting the distribution of submerged macrophytes in lakes with different nutrient levels; pH, NH4+-N and WT were the key factors in oligotrophic-mesotrophic lakes; and WT, conductivity and Chl-a were the key factors in eutrophic lakes. In conclusion, we identified the key water environmental factors affecting the distribution of submerged macrophytes in lakes on the Yunnan Plateau, which will provide guidance for the restoration, protection and management of submerged macrophytes in lakes in Yunnan plateau lakes.
Key words: plateau lakes; submerged macrophytes; spatial distribution; environmental factors; redundancy analysis
