寧德三都澳海島馬尾松林群落特征及物種多樣性
2023-12-26 02:44:56肖集泓賴敏英王春曉鐘址非溫清燕錢志軍王迎雪鄧清雅鄧傳遠
生態學報 2023年23期
肖集泓,賴敏英,李 媚,王春曉,鐘址非,溫清燕,錢志軍,羅 真,王迎雪,鄧清雅,鄧傳遠
福建農林大學風景園林與藝術學院,福州 350002
因地理位置特殊,海島常年風速較大,受臺風浪潮侵襲,土壤瘠薄鹽分含量高,且因規模有限和空間隔離,海島生態系統具有明顯的獨立性與資源短缺性,屬于典型的脆弱型生態系統,在獨特的自身環境與復雜的干擾下,表現出易受損性和難恢復性[1]。森林生態系統作為組成結構復雜且物種資源最為豐富的生態系統類型,不僅提供了重要的自然資源,而且對保護脆弱的生態系統具有重要的意義。大量的原始天然林被破壞后,許多海島通過人工林種植和生態保護政策管理等人為調控方式進行植被重建和修復,并借此減弱和消除海島生態脆弱性[1—3]。海島人工林多以單一樹種進行種植建設。單一樹種種植的人工林群落結構簡單,抵抗病蟲害和自然災害的能力較弱,林下植被和其他地被植物及微生物等相對缺乏,因此常表現出林分物種多樣性極低,森林群落生態服務功能性差,地被植物及地表枯落物少,土壤養分等肥力較低,保持水土及通養水源等方面能力極弱等缺點[2—4]。且目前仍然缺乏相關技術和研究資料以支撐我國海島植被修復,導致野生植物資源衰退嚴重,修復選用的物種也因不適應海島或群落的環境而被淘汰[5]。因此,對相對穩定的海島半自然林進行相關研究,不僅能夠豐富海島植物資料,了解人工林向自然林演替過程中植物組成和變動,同時能夠了解演替過程、植被穩定性、群落多樣性等特點,及了解限制群落演替進程的各種影響因子,這對海島生態和植被的修復等具有十分重要的指導意義。
群落的結構特征和物種多樣性是生態學研究的熱點之一[6]。目前,國內外學者多集中于對內陸和外海島嶼的植被進行研究[7—10],對內海島嶼的相關研究較為缺乏。三都澳海島屬內海島嶼,該區域也是南亞熱帶和中亞熱帶的過渡區域,地理位置十分重要,但目前對于該區域植物研究僅見對苔蘚植物多樣性的調查[11]。馬尾松(Pinusmassoniana)是松科(Pinaceae)松屬常綠針葉大喬木,適應性強,耐干旱瘠薄,是恢復退化土地的優秀先鋒樹種[12]。馬尾松因適應性強,在三都澳地區,特別是環境較內陸惡劣的海島上廣泛分布,占據著當地最大的森林面積,是當地最具代表性的森林類型,具有不可替代的生態價值。三都澳地區的馬尾松林為上個世紀飛機撒播造林或人工造林形成,受當地政策的制約,人為影響相對較小,屬于半自然群落。自然林的保護和恢復是目前我國實施的一項龐大而復雜的社會工程,自然林資源的合理開發和研究逐漸成為社會關注的熱點問題[4]。自然或半自然林林下植物豐富多樣,伴隨著群落的演替更新和植物的優勝劣汰,適合在群落中生存的優勢物種逐漸體現,共同構成了相對穩定的群落結構。因此,對三都澳半自然馬尾松林進行群落特征和物種多樣性研究,有利于了解其群落中的主要物種組成及相對穩定的群落結構特點,這對當前海島植被修復及海島人工林建設具有一定的指導意義。且當地的馬尾松林存在植株老化死亡、自然更新困難等問題,其未來的持續經營和管理也亟需相關研究提供基礎資料參考。因此本研究通過野外調查三都澳6個海島的馬尾松林,分析其物種組成和植物功能性狀,同時劃分其植被類型,探究其演替規律,并分析物種多樣性及其與環境因子的相關性,旨在摸清三都澳海島馬尾松林群落特征、演替狀況,及其物種多樣性與環境因子的相關關系,豐富當地植被基礎資料,彌補內海島嶼相關研究領域的空缺,為海島植被修復、海島人工林建設及當地馬尾松林的合理經營和管理提供理論基礎。
1 材料和方法
1.1 研究區概況
三都澳位于福建省寧德市東南部,為中國1.84萬km黃金海岸線的中點,距寧德市區30 km,是世界級天然深水良港。共有島嶼126個,有居民島嶼17個,其中最大的島嶼是三都島,總面積29.3 km2,最高海拔461 m,青山島、長腰島、白匏島、雞公山島和斗帽島面積分別為9.48 km2、2.22 km2、1.12 km2、0.89 km2和0.6 km2[11]。研究區屬典型的亞熱帶海洋季風氣候,地形以丘陵為主,土壤類型以紅壤和黃壤為主,島上以次生馬尾松(Pinusmassoniana)針葉林和馬尾松針闊混交林分布最廣。
1.2 樣地設置與調查
基于實地踏查,于2022年6—8月采用典型選樣的方法對三都島、青山島、長腰島、雞公山島、白匏島、斗帽島6個面積較大和森林覆蓋率較高的有居民海島進行樣地設置,6個海島分別設置有20、6、5、4、3和2個樣地(圖1),共設置40個20 m×20 m的森林樣方。以20 m×20 m作為喬木層樣方面積,森林樣方4個角設置4個面積為5 m×5 m的灌木樣方,4個角及中心位置設置5個面積為1 m×1 m的草本樣方。記錄喬木層(株高≥ 5 m或胸徑≥ 3 cm的喬木、竹類、大型灌木、附生植物和藤本植物)、灌木層(株高<5 m且胸徑<3 cm的灌木、攀援灌木、喬木幼樹和竹類,統計時剔除以攀援狀依附植物生長的攀援灌木植株)和草本層(含攀援草本和蕨類,統計時剔除以攀援狀依附植物生長的攀援草本植株)每一棵植株的種名、胸徑、株高、冠幅(或蓋度)、數量等信息[13]。采用全球定位系統(GPS)和森林羅盤儀記錄樣方的經緯度、海拔、坡向、坡度和坡位。將坡位劃分為上、中、下坡;坡向劃分為陽坡(157.5°—247.5°)、半陽坡(112.5°—157.5°、247.5°—292.5°)、陰坡(0°—67.5°、337.5°—360°)、半陰坡(67.5°—112.5°、292.5°—337.5°)[14]。采用林冠數字圖像Photoshop分析法[15]測定群落郁閉度,樣方基礎信息見表1。
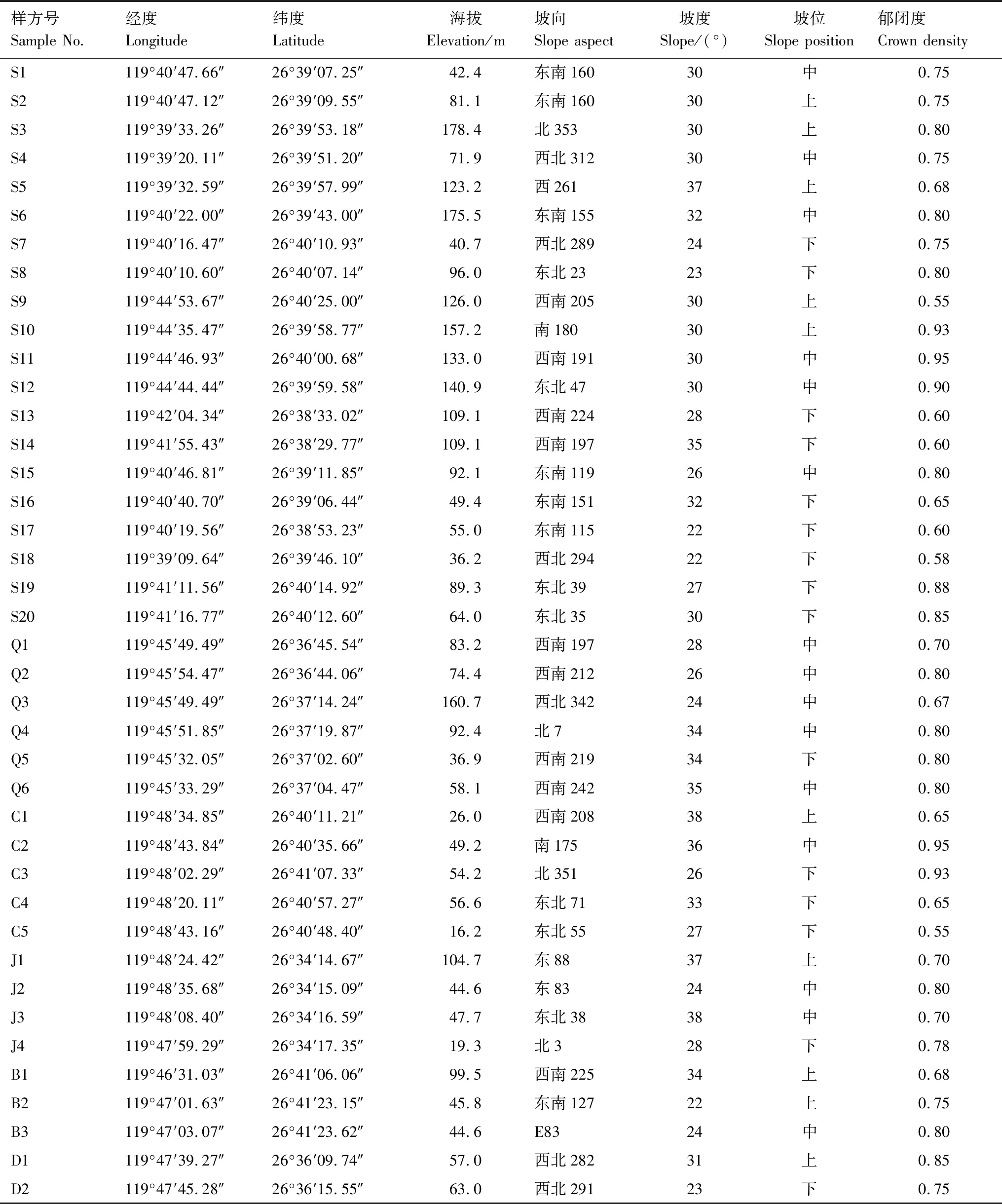
表1 樣方概況
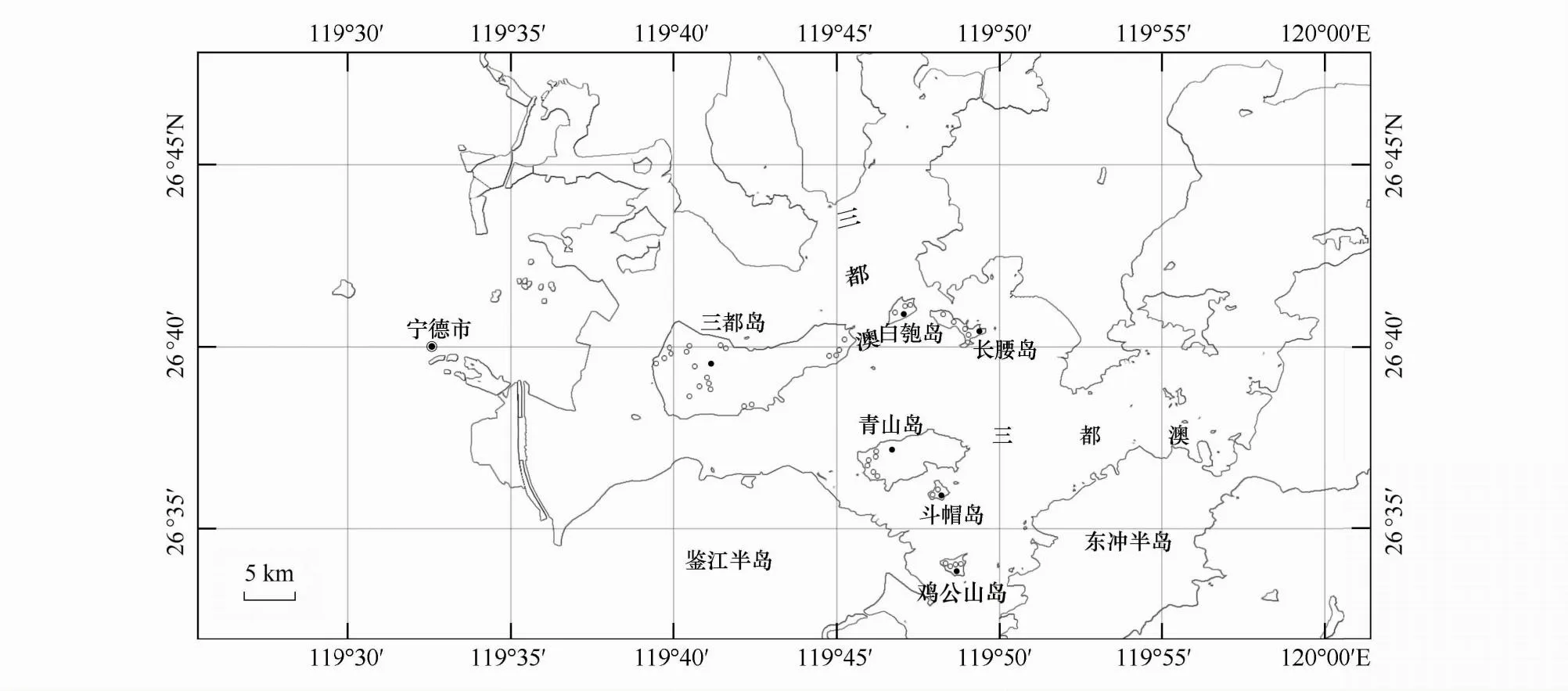
圖1 三都澳6個海島區位
1.3 數據處理與分析
根據《中國植物志》[16](http://www.iplant.cn/foc)和《福建植物志》[17]整理研究區植物名錄,記錄分布區域,并區分鄉土植物和外來植物,采用最新修訂的拉丁名和中文名。由于亞熱帶地區群落結構和物種組成復雜,應通過數量分類方法(聚類分析)進行植物群落分類[13],因此,根據群落的物種重要值(剔除重要值<5的偶見種),采用目前植物群落分類中較為常用的組平均聚類法[9—10, 18]劃分植物群落類型。利用冗余分析(RDA)解釋環境因子對物種多樣性的影響。利用二次函數擬合分析灌木層鵝掌柴(Heptapleurumheptaphyllum)與群落郁閉度的關系。重要值和物種多樣性計算公式[19—20]如下:
重要值=(相對多度+相對頻度+相對顯著度)/3
(1)
Shannon-Wiener指數(H):
(2)
Simpson指數(D):
(3)
Pielou指數(E):
(4)
Patrick指數(R):
R=S
(5)
式中,各層片相對顯著度分別采用喬木胸高(1.3 m處)斷面積和灌草層植物蓋度計算。S為各樣方中出現的物種數,N為所有物種個數之和,Pi=Ni/N,Ni為第i個物種的個體數(i=1,2,3...)。
群落多樣性指數=0.5D1+0.3D2+0.2D3
(6)
式中,D1、D2和D3分別表示喬灌草3個層片的多樣性指數[9, 21]。利用SPSS軟件的單因素方差分析計算各群系、各植被型的多樣性指數,同時選擇最小顯著性差異法(LSD)進行事后方差比較不同群系、及各層片之間的多樣性指數差異水平,由于植被型只有兩種,因此選擇t檢驗進行多樣性指數差異性水平分析。通過方差分析比較本研究地與平潭海島[9]森林植被物種多樣性的差異。
采用五點取樣法采集各樣方25 cm深的土壤樣品,研磨過篩后重復3次測定其9項指標:pH值、全鉀、速效鉀、全磷、有效磷、全氮、堿解氮、有機質和水溶性鹽總量[22],土壤信息見表2。
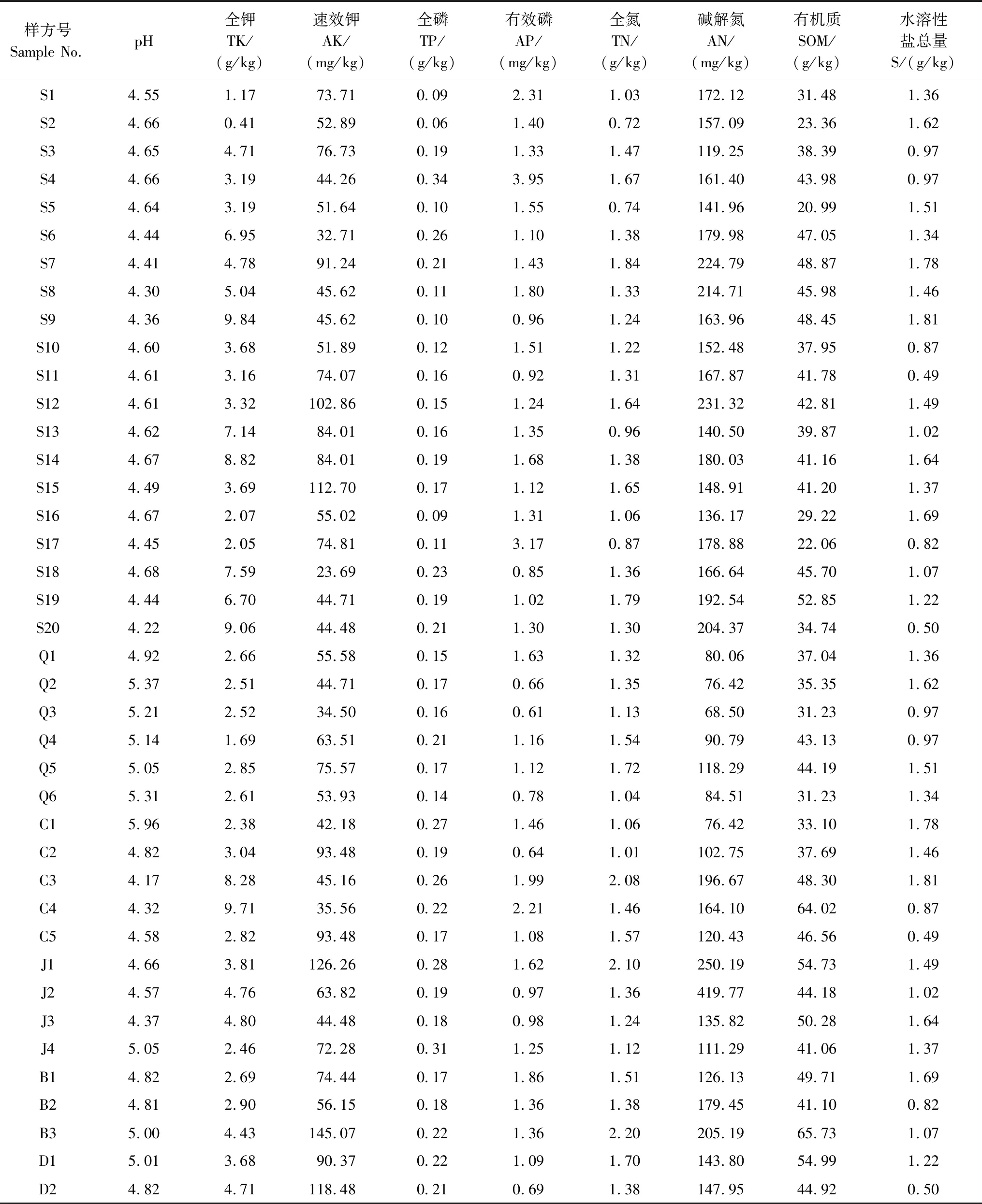
表2 土壤化學指標
分析群落中種子植物的生長型、生活型、對光的耐受性3項定性功能性質,具體劃分依據為:生長型分為喬木、灌木、草本、藤本;生活型劃分為9類,即高位芽植物(含大型>30 m、中型8—30 m、小型2—8 m和矮小型0.25—2 m共4類)、地上芽、地面芽、藤本附生和一年生植物;植物對光的耐受性分為陽性、中性和陰性[9, 23]。
2 結果與分析
2.1 物種組成和植物功能性狀
統計40個樣方調查數據,發現三都澳海島馬尾松群落內維管束植物共計255種(含5變種),隸屬于86科168屬,屬種系數為66.80%。其中以被子植物為主,共計72科150屬231種(含5變種),蕨類植物10科16屬22種,裸子植物僅2科2屬2種,即馬尾松和杉木(Cunninghamialanceolata)。研究區植物來源統計,鄉土植物78科159屬244種,外來植物7科12屬12種,其中含1級惡性入侵植物2種,即鉆葉紫菀(Symphyotrichumsubulatum)和小蓬草(Erigeroncanadensis),但在林中重要值占比很低。
對255種維管束植物的3個定性功能性狀進行統計分析(見表3)。由表3可知,在生長型上,以喬木和灌木植物種類最多,分別占總物種數的37.25%和30.20%;在對光的耐受性上,以中性植物數量最多,占總物種數的69.02%,其中陽性樹種都為木本植物;在生活型上,以小型、矮小型和中型高位芽植物為主,分別占總物種數的29.80%、27.84%和24.71%,大型高位芽植物和一年生植物占比極小,分別占比1.57%和2.35%。

表3 三都澳6個海島馬尾松群落維管束植物功能性狀統計
2.2 植物群落類型劃分
參照《<中國植被志>研編內容與規范》有關植被分類原則和群落命名原則[13],選取40個樣方中重要值>5%的物種,利用組平均聚類法對各個樣方的物種重要值進行聚類分析(圖2)。由圖2可知,當歐氏距離為20時,由上至下將三都澳6個海島的馬尾松群落劃分為5類群系:馬尾松林、馬尾松+檵木(Loropetalumchinense)林、馬尾松+鵝掌柴林、馬尾松+楊桐(Adinandramillettii)林、馬尾松+茶竿竹(Pseudosasaamabilis)林,最終劃分為33類群叢(見表4)。統計40個樣方各層片重要值占比最大的優勢種重要值和數量(見表5),由表5可知,40個樣方的喬木層優勢種均為馬尾松,灌木層優勢種主要有:鵝掌柴、楊桐、九節(Psychotriaasiatica)、桃金娘(Rhodomyrtustomentosa)、玉葉金花(Mussaendapubescens)、豺皮樟(Litsearotundifoliavar.oblongifolia)等,草本層最主要優勢種為芒萁(Dicranopterispedata),此外還有團葉鱗始蕨(Lindsaeaorbiculata)、扇葉鐵線蕨(Adiantumflabellulatum)、狗脊(Woodwardiajaponica)等優勢種。

表4 三都澳6個海島馬尾松群落植被類型
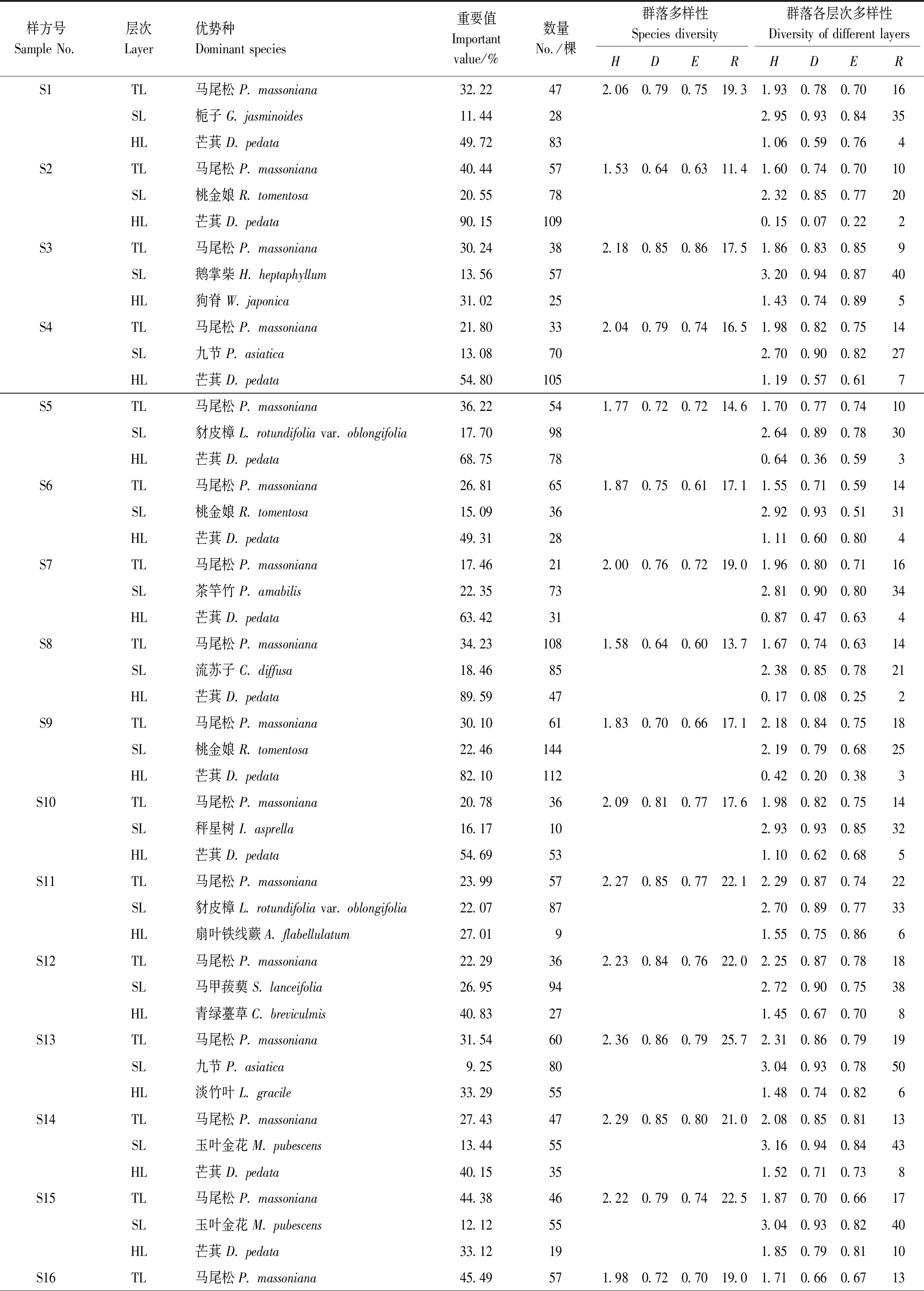
表5 三都澳6個海島馬尾松林植被特征及物種多樣性指數
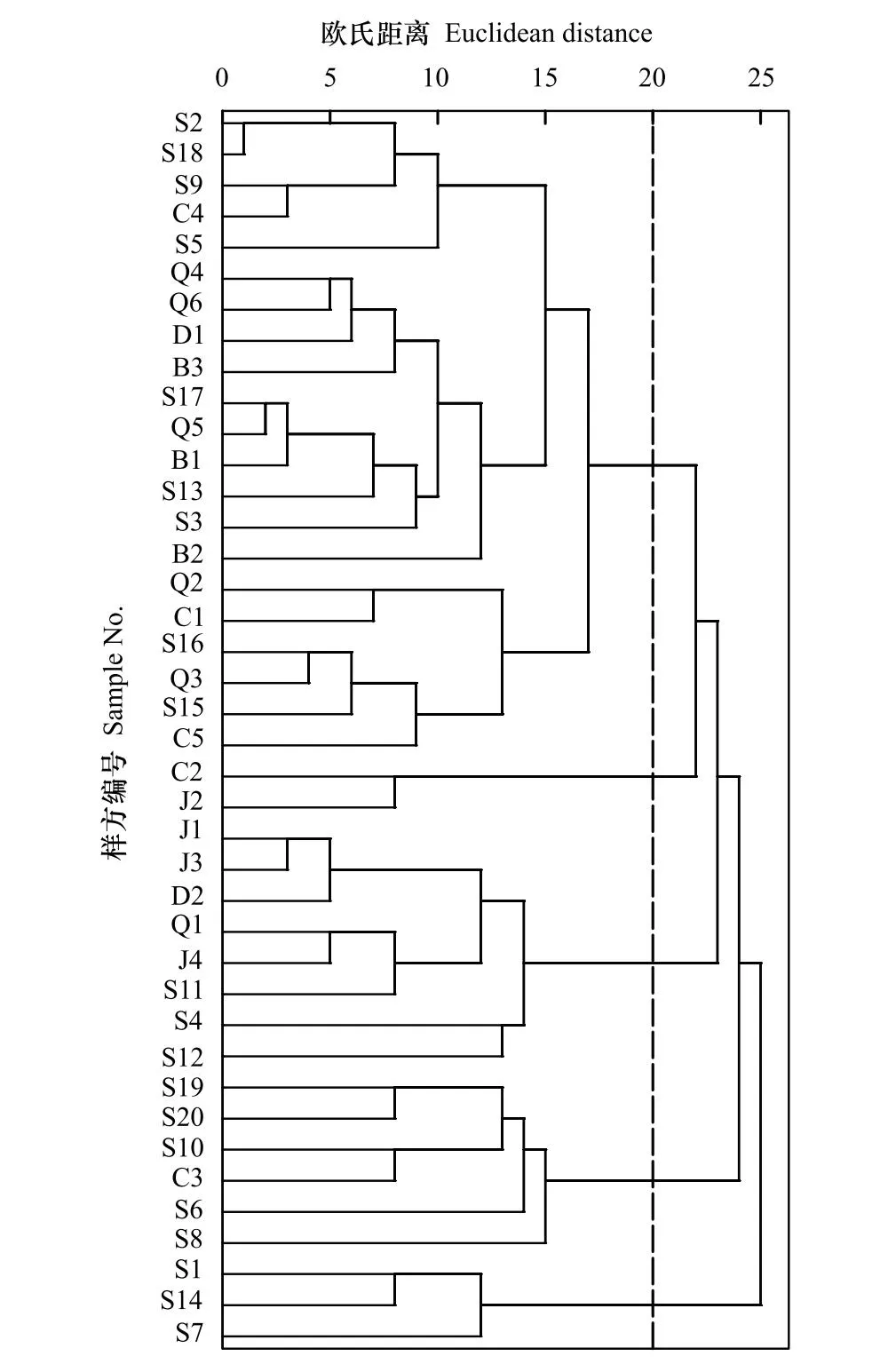
圖2 植物群落組平均聚類
2.3 植物群落物種多樣性與環境因子的相關性
根據調查數據計算40個樣方各層片及群落整體的物種多樣性指數(包括Patrick指數、Simpson指數、Shannon-Wiener指數、Pielou指數),見表5。由表5可知,在群落各層片上,灌木層的物種多樣性指數均遠高于同樣方的喬木層和草本層。對不同群系、植被型的樣方進行多樣性指數統計計算,并標記顯著性差異(見表6、表7)。由表6、表7和方差分析及t檢驗結果可知,5類群系和2種植被型的灌木層多樣性指數(除E外)均顯著(P<0.05)高于喬木層和草本層,表明研究地群落以灌木層物種最豐富、群落最穩定、結構最復雜。從植被型差異來看,馬尾松常綠針闊混交林僅喬木層D和群落整體E顯著(P<0.05)高于馬尾松常綠針葉林,其他指數差異不顯著(P>0.05)。從群系差異來看,5類群系喬木層多樣性指數差異均不顯著(P>0.05),II群系的灌木層R顯著(P<0.05)低于其他4類群系,I群系的草本層R顯著(P<0.05)高于其他群系,II群系的草本層H、D、R均顯著(P<0.05)低于其他4類群系,II群系的群落整體H、D、R顯著(P<0.05)低于其他3類混交林群系,V群系的群落整體H、D均顯著(P<0.05)低于除II外的其他混交林群系。

表6 不同群系的物種多樣性指數

表7 不同植被型的物種多樣性指數
對40個樣方的物種多樣性指數(含群落整體和各層片的Patrick指數、Simpson指數、Shannon-Wiener指數、Pielou指數,見表5)和14個環境因子(含表2的9個土壤因子、表1的4個地形因子(海拔、坡向、坡度、坡位)和群落郁閉度)進行冗余分析(RDA)分析,結果見圖3。分析結果篩選出2個顯著影響(P<0.05)物種多樣性的環境因子:土壤pH和速效鉀(AK)。土壤pH與群落Patrick指數、Shannon-Wiener指數及灌草層物種多樣性指數呈正相關,與群落Pielou指數、Simpson指數及喬木層物種多樣性指數呈負相關;速效鉀與群落整體、喬木層、灌木層和草本層的物種多樣性指數均呈正相關。而土壤pH和速效鉀這兩個環境因子共同對物種多樣性的總解釋量為27.6%,表明還有72.4%的未知因素影響了三都澳海島的馬尾松群落物種多樣性。
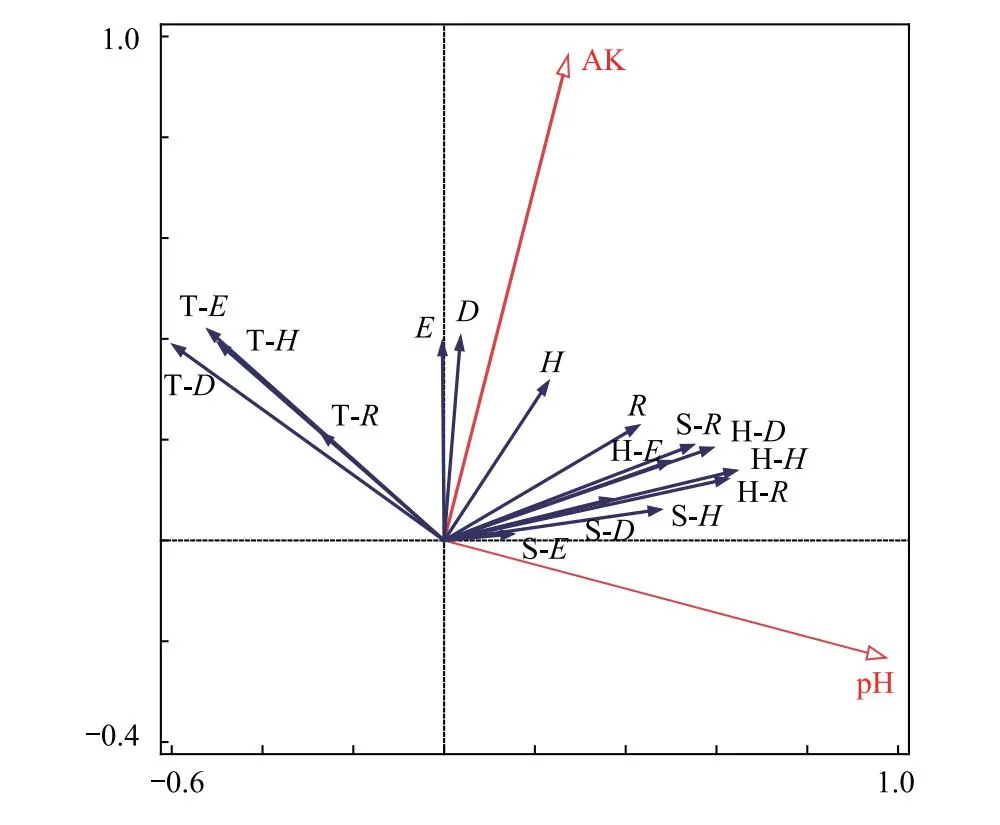
圖3 物種多樣性指數與環境因子RDA分析圖
鵝掌柴是研究地灌木層重要值最大的物種,屬半陽樹種,能于全日照、半日照、半蔭庇的環境中生存,以半陽或者半陰的環境生長最佳。由于RDA分析反映的是線性回歸分析結果,不能反映出非線性相關模型,因此對灌木層鵝掌柴的重要值與群落郁閉度進行二次函數擬合(剔除群落中未出現鵝掌柴的4個樣方S2、S6、S8、S18),結果見圖4。可知,灌木層鵝掌柴的重要值與群落郁閉度符合二次函數關系,二者呈顯著(P<0.05)相關關系。當郁閉度過高或者過低時,鵝掌柴的重要值均占比極小,符合鵝掌柴對光照的適應性結果。
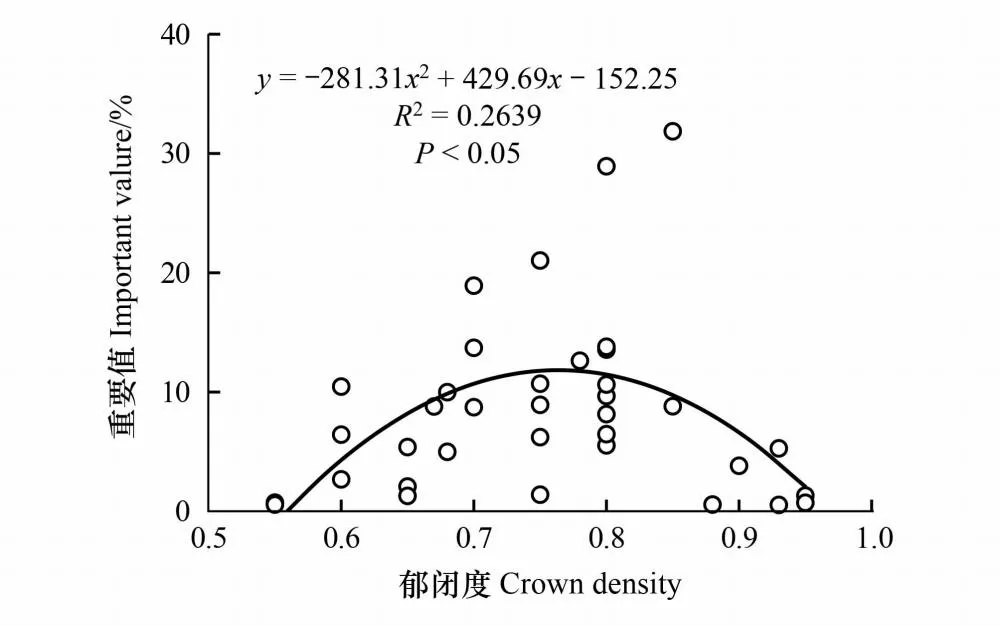
圖4 鵝掌柴重要值與群落郁閉度二次函數擬合
3 結論與討論
3.1 三都澳6個海島馬尾松群落分類和演替
在前人的研究中發現,自然或近自然的亞熱帶森林植被以中型和小型高位芽植物占比為主,一年生植物相對較少,而次生群落則表現出灌木和(一年生)草本植物占比優勢明顯,高位芽植物生態位喪失[9,24—25]。本研究區以小型、矮小型和中型高位芽植物為主,一年生植物占比極小,喬木樹種占比最大,表現為近自然狀態的植被類型,而矮小型高位芽植物占比大可能是受海島環境影響,植物的適應性選擇[10]。但生境異質性會導致同一植物物種表現為不同的生活形態[10],受海島環境的限制與影響,研究區內的大型高位芽植物種類占比最小,研究地記錄的最高物種為馬尾松,屬于大型高位芽植物(30m以上),但群落中最高統計記錄不超過25 m,計算其平均高度僅10 m左右,且本研究地記錄的95種喬木樹種中,喬木層僅見81種,其他14種以灌木狀分布于灌木層,具體體現為灌木層多樣性指數(除E外)均明顯高于喬木層(表5、6和7),表現出了次生群落的特征。因此,可以將研究地的植被類型劃分為具次生性質的自然植被,或稱之為半自然植被[26]。
出現在喬木層的竹類、大型灌木、附生植物和藤本植物是喬木層中穩定的組成成分,因此要統計為該層的物種,而根據需要,喬木層可進一步劃分為大喬木層(高度≥25 m)、中喬木層(8 m≤高度<25 m)和小喬木層(5 m≤高度<8 m)3個亞層[13]。研究地記錄的最高物種為馬尾松,高度未超過25 m,且計算每個樣方的物種平均高度發現,馬尾松平均高度9.1—12 m不等,而喬木層中重要值較高的鵝掌柴、楊桐、茶竿竹、檵木平均高度為5.5—7.5 m不等,喬木層幾種優勢植物平均高度差異不明顯,從實際生長高度來看都屬于中小型喬木,因此喬木層不做進一步劃分。根據各群落中物種重要值的組平均聚類,將馬尾松群落劃分為5類群系,分屬于2種植被型(見圖2和表5)。由于灌木層結構復雜,且部分樣方的灌木層和草本層優勢種不明顯,表現出同一層片多優勢種共存的現象,符合亞熱帶群落結構和物種組成復雜多樣的特征[13],因此最終劃分為33類群叢(見表5)。5類馬尾松群落類型中,喬木層重要值最高的都是馬尾松,是絕對優勢樹種。大多數樣方草本層重要值較高的物種較為相似,以芒萁重要值占比較大,表明三都澳海島馬尾松群落林下草本層重要值較高的物種種類受林分類型影響不大,但不同林分類型中,林下灌木層各物種重要值存在較大差異,這與張維偉等[27]的研究結果類似。在群落中,物種之間的相互作用會影響群落的物種組成及其個體數量[28],同時外界的干擾也會導致物種組成差異[29]。由圖4可知,當群落郁閉度過高或者過低時,鵝掌柴重要值均較小。根據重要值數據,在S10—12、S19和C2—3這6個郁閉度>0.85的樣方中,灌木層中鵝掌柴的重要值僅有5.28、1.32、3.82、0.57、0.72和0.54,鵝掌柴屬半陽樹種,以半日照或者半陰環境生長最好,過高的林分密度則不利于鵝掌柴的生長。在S11—12兩個馬尾松-鵝掌柴混交林中,鵝掌柴雖在喬木層占據極大的重要值,但在高郁閉度的林下灌木層仍然難以生存,長此以往,這些樣方的鵝掌柴將逐漸被其他物種淘汰,使得林分結構發生改變,群落穩定性受到影響。同理,馬尾松屬陽性植物,在灌木層僅在S9和C5兩個極低郁閉度的馬尾松林中出現,在林分結構復雜且郁閉度過高的群落中明顯存在自然更新問題。因此,本研究5種類型馬尾松群落灌木層之所以出現物種組成及其重要值差異,原因之一極可能是受郁閉度的影響。
從演替方向看,研究地以人工馬尾松常綠針葉林作為起點,起初受人為砍伐破壞的影響,群落演替速度較慢,后經當地政策的干預,人為干擾逐漸減弱,在群落恢復過程中表現出次生性質。隨著時間的推移,群落正逐漸恢復至近自然狀態,但是目前仍然存在次生群落的特征。群落由常綠針葉林逐步向常綠針闊葉混交林演替的過程中,強陽性針葉樹種馬尾松將逐步被其他陽性闊葉樹種取代,如鵝掌柴、楊桐等,闊葉樹種將逐漸占據群落的主要地位,群落郁閉度也將逐漸增加。當郁閉度高達一定的程度時,不耐蔭蔽的闊葉樹種如鵝掌柴等,在高郁閉度的林下將難以生存,自然更新能力下降,生態位逐漸降低,將被逐步淘汰,而強耐陰的陽性樹種如楊桐等,將逐步成為該區域穩定群落的優勢種或建群種。最終,研究地群落將自然演替為亞熱帶頂級群落-常綠闊葉林,同時針葉林林下草本層最主要優勢種芒萁也將隨著群落從針葉林向闊葉林的演替而逐漸喪失優勢地位。
3.2 三都澳6個海島馬尾松群落物種多樣性特征及環境因子解釋
Shannon-Wiener指數(H)越高,群落越復雜,生物種類越多;Simpson指數(D)越大,群落內物種數量分布越不均勻[7]。本研究中,不同林分類型物種多樣性有所不同,總體而言,馬尾松林的群落整體H、D均顯著(P<0.05)低于除II外的其他混交林(表6)。主要原因是,混交林中喬木層植物種類多,生態位差異大,對資源利用更充分[27]。而單一馬尾松林喬木層形成的單一脅迫,會降低林下植物的數量,進而影響其物種多樣性指數,且植物數量的減少難以提供足夠的凋落物供給微生物分解,使得馬尾松林內的生境條件比其他混交林差,進一步降低了物種多樣性指數。同時,根據表2土壤數據可知,速效鉀含量最高的10個樣方中,除C4屬于馬尾松林外,其他都是混交林,從RDA分析結果也發現,速效鉀與各層片物種多樣性指數均呈正相關性,隨著植物生長所必須的營養元素含量的增加,使得各層片植物能獲取的營養更多,多種植物生長狀況越好,群落就越復雜,從而逐漸導致優勢種的地位越下降,最終群落表現出各層片多個優勢種共存的現象。反之,各層片營養物質匱乏,能夠適應最低限度的營養物質的物種才能存活并繁衍,各層片的優勢種明顯,占據著群落分布中的優勢地位,因此,較之混交林,馬尾松針葉林各層片的結構表現就更為簡單,從而導致其群落整體H、D均顯著(P<0.05)低于除II外的其他混交林群系。何雅琴等[9]對大練島的森林群落多樣性研究也表明,臺灣相思+木麻黃林的群落整體H、D均顯著(P<0.05)高于臺灣相思林和木麻黃林。表明混交林較于純林,更能增加群落的生物種類,并提高Shannon-Wiener和Simpson指數的大小。
Pielou指數(E)越高,植物群落結構越穩定[30]。本研究地馬尾松常綠針闊混交林群落整體E值顯著(P<0.05)高于馬尾松常綠針葉林(表7),與大練島[9]的方差分析比較結果也表明,大練島的臺灣相思林和臺灣相思+木麻黃林的群落整體E值顯著(P<0.05)高于本研究地的馬尾松常綠針葉林,這與群落演替規律相符,群落演替越后期,群落越穩定[31]。Patrick指數(R)直接反映群落物種豐富度[7]。II群系(馬尾松+楊桐林)的灌木層豐富度指數R顯著(P<0.05)低于其它4類群系,主要原因可能是,II群系的6個樣地郁閉度均較大(0.8,0.8,0.85,0.88,0.93和0.93),研究地有41種陽性樹種(都為木本植物)和鵝掌柴等不耐蔭庇的半陽性樹種,鵝掌柴在陽光不足的高郁閉度林下灌木層占比較低,甚至難以存活(圖4),而馬尾松林樹冠較之更窄,更有利于光照進入林下,使得林下植物,特別是陽性木本植物的豐富度更高。與大練島[9]的方差分析比較結果發現,本研究地馬尾松林豐富度指數R顯著(P<0.05)高于大練島的所有森林群落類型(臺灣相思林、木麻黃林及2種物種的混交林)。引起這一結果的主要原因在于,馬尾松林喬木層和灌木層豐富度指數R均顯著(P<0.05)高于大練島的所有森林群落類型的喬木層和灌木層R值。由于馬尾松較于木麻黃和臺灣相思,生長高度更高,與其他喬木物種存在較大的高度差,在資源競爭時能形成垂直方向的互補[32],同時其林冠密度更低,更有利于陽性樹種的生長,因此其喬木層物種更豐富;而灌木層計算喬木幼樹,因此豐富的喬木層物種也一定程度上促進了灌木層物種的豐富度,因此馬尾松林喬灌層指數R更高。但馬尾松林草本層指數R顯著(P<0.05)低于大練島的所有森林群落類型的草本層R值。其主要原因在于,馬尾松林草本層多以芒萁為優勢物種,芒萁對雜草密度、種子萌發和幼苗生長的具有化感效應[33—34],嚴重減少了草本層物種,同時,竹類在馬尾松林喬木層、灌木層均分布較廣,導致草本層光照嚴重不足,且竹類的凋落物也降低了草本植物的生存空間[35],因此導致馬尾松林草本層R指數最低。I群系(馬尾松+鵝掌柴林)的草本層R值均顯著(P<0.05)高于本研究的其他群系,II群系的草本層H、D、R均顯著(P<0.05)低于其他4類群系,主要原因是II群系所有樣方草本層均以芒萁為單一優勢種,III—V群系的草本層也以芒萁作為優勢種居多,但I群系有一半的樣方草本層優勢種不是芒萁,以芒萁作為優勢種的Q1、J1樣方芒萁的重要值不高,沒有形成致密層。芒萁在林下形成致密層時會大量截留林冠喬木的凋落物,進而影響其凋落物分解和養分循環過程[36],加之芒萁具有化感效應[33—34],因此抑制了草本層其他物種的生長,物種豐富度也大大降低。同時,這些樣方的土壤pH和速效鉀較于其他群系樣方的更低,土壤pH和速效鉀與灌草層的物種多樣性指數均呈正相關性,pH過低,植物對大量元素的吸收減弱[37],而鉀含量過低,會降低植物酶的激活、蛋白質的合成和養分的運輸等[38],從而制約了植物的生長發育。而土壤pH與喬木層的物種多樣性呈負相關性,主要原因可能是喬木層的主要物種如馬尾松、鵝掌柴、茶竿竹等都喜歡生活在偏酸的土壤中,因此pH越高,越不利于喬木層植物的生長。
4 建議
馬尾松混交林相較于馬尾松林物種豐富度更高,群落穩定性更強,但相對的,混交林過高的林分郁閉度嚴重制約了林下灌木層植物的生存環境,物種豐富度明顯降低,馬尾松、鵝掌柴等建群種的自然更新受到嚴重制約,不利于群落的長期穩定發展。因此未來馬尾松森林經營中要結合考慮這兩個問題,通過人為撫育間伐,促進馬尾松林向混交林轉換,同時控制林窗面積,增大林內的輻射強度,也通過補植、控制土壤pH和速效鉀的含量等,促進群落長期穩定發展。
研究地中已見1級惡性入侵植物鉆葉紫菀和小蓬草2種,且在樣線踏查時,發現島上也分布有其他1級惡性入侵植物,如假臭草(Praxelisclematidea)、鬼針草(Bidenspilosa)、馬纓丹(Lantanacamara)等。研究地草本層以芒萁為最高優勢種,芒萁的化感效應不僅抑制了惡性入侵植物的入侵,也抑制了草本層一些植物的生長,導致大部分樣方草本層多樣性指數最低。且芒萁單優草本層片雖然廣泛分布于亞熱帶低山丘陵區的多種退化群落,但消退于常綠闊葉森林群落。若芒萁消退,對入侵植物的化感作用也將減少,而1級惡性入侵植物的入侵性極強,會嚴重威脅本土植物的生態位,因此需要通過人為干預手段控制林下芒萁密度,既能增加草本層物種豐富度,也能有效抑制入侵植物的擴散。
研究地分布有國家一級保護植物野生建蘭(Cymbidiumensifolium)、國家二級保護植物野生茶(Camelliasinensis)和花櫚木(Ormosiahenryi),在對研究地進行人為撫育時,應注意加強對這些植物的保護力度,防止人為采摘、毀壞,必要時應采取圈地、引種等保護手段。
猜你喜歡
課堂內外·初中版(科學少年)(2025年1期)2025-02-28 00:00:00
課堂內外·初中版(科學少年)(2025年2期)2025-02-28 00:00:00
英語世界(2023年10期)2023-11-17 09:18:18
科學大眾(中學)(2019年3期)2019-05-17 10:04:30
汽車觀察(2018年10期)2018-11-06 07:05:26
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
少兒科學周刊·兒童版(2015年10期)2015-11-07 03:42:03
