柑橘酸腐病菌PacC的生物信息學分析及其響應pH值的表達模式
2023-12-14 12:45:18譚小麗胡近近陶能國歐陽秋麗李茂慧
食品科學 2023年22期
鄭 杰,譚小麗,胡近近,陶能國,歐陽秋麗,李茂慧,李 路
(湘潭大學化工學院,湖南 湘潭 411105)
柑橘作為我國重要的經濟作物,種植面積和產量均居世界第一,其風味獨特,營養豐富,深受廣大消費者的喜愛[1]。但柑橘在采后極易受到指狀青霉(Penicillium digitatum)、意大利青霉(P.italicum)、酸腐病菌(Geotrichum citri-aurantii)等采后致病真菌的侵染[2-3]。近年,柑橘酸腐病的發生呈上升趨勢,在潮濕多雨季節尤為嚴重[4-5]。化學防治是防控柑橘酸腐病的主要方法,但常用于柑橘病害防控的殺菌劑如抑霉唑、咪鮮胺等對酸腐病的防控均沒有顯著效果,目前只有雙胍鹽類藥物對酸腐病菌具有一定的抑制作用[6-7]。因此,深入研究酸腐病菌與柑橘果實在采后貯藏期的互作關系對于酸腐病的防控具有重要意義[8]。
環境pH值對病原真菌的生長發育和代謝具有一定調控作用,而病原真菌對環境pH值也具有一定的適應和調節能力。其中,由轉錄因子PacC介導的pH值信號響應途徑在微生物中廣泛存在,調控多種植物病原真菌的環境適應性、次級代謝和致病性[9]。例如,PacC的缺失可以影響果生刺盤孢(Colletotrichum fructicola)的菌絲生長、孢子萌發、附著胞形成及致病力[7];擴展青霉(P.expansum)PacC缺失突變體的生長與分生速率及其對蘋果和梨果實的致病力均顯著下降,且在pH>6.0時不能產生棒曲霉毒素[10]。目前,酸腐病菌對環境pH值的適應性以及PacC的調控作用還未完全揭示。因此,本研究首先研究不同pH值對酸腐病菌生長的影響,在全基因組水平篩選酸腐病菌PacC序列的信息,對其親緣關系、基因及其編碼的蛋白質結構進行預測,隨后測定其在不同pH值以及果實侵染過程中的表達模式,以期為進一步研究柑橘酸腐病菌的致病機理奠定基礎。
1 材料與方法
1.1 材料與試劑
酸腐病菌菌株從湘潭大學附近果園中自然腐爛的柑橘果實表面分離純化得到[11],現保存于湘潭大學生物與食品工程系菌種保藏室。
總RNA提取試劑盒 福州飛凈生物科技有限公司;RNA反轉錄試劑盒、qPCR SYBR Green Master Mix、PhyZolTMTotal RNA Extraction Reagent、Hifair?III 1stStrand cDNA Synthesis SuperMix for qPCR逆轉錄試劑盒、Hieff UNICON?Universal Blue qPCR SYBR Green Master Mix試劑盒 翌圣生物科技(上海)股份有限公司。
1.2 儀器與設備
PHS-W型pH計 上海班特儀器有限公司;ABI 2720型PCR儀 美國ABI公司;OSE-260-03型超微量定量儀 天根生化科技(北京)有限公司;LightCycler96型熒光定量儀 瑞士Roche公司;FA1004N型分析天平 上海精密科學儀器廠;SW-CJ-1D型垂直凈化工作臺 濟南啟科儀器設備有限公司。
1.3 方法
1.3.1 柑橘酸腐病菌PacC候選基因的篩選
本課題組前期送樣至北京諾禾致源生物信息技術有限公司使用SMRT技術,結合Illumina Hiseq-PE150測序和Pacific Biosciences RSII測序對酸腐病菌基因組進行測序[12]。根據基因注釋信息篩選出PacC候選基因,并對其進行分析。
1.3.2 不同物種間PacC基因編碼氨基酸序列比對與保守基序分析
在NCBI(https://www.ncbi.nlm.nih.gov)中查找并下載其他采后致病菌PacC編碼的氨基酸序列,利用軟件ClustalX和GeneDoc將不同物種間同源基因的氨基酸序列比對,利用在線網站MEME Suite 5.4.1(https://memesuite.org/meme/)并結合MEGA7.0軟件與TBtools軟件預測該基因的保守基序[13-14]。
1.3.3 不同物種間的PacC系統進化分析
利用MEGA 7.0軟件中的鄰接法構建系統發育樹,并進行1 000 次重新采樣評估,其他參數均為標準值[15]。
1.3.4 柑橘酸腐病菌PacC結構特征及染色體分布
利用在線網站GSDS 2.0(http://gsds.gao-lab.org/)對PacC轉錄因子家族成員的基因結構進行分析,利用在線網站MG2Cv2.1(http://mg2c.iask.in/mg2c_v2.1/)對其家族成員所在染色體位置進行分析[16]。
1.3.5 酸腐病菌PacC編碼蛋白結構與理化性質分析
利用NCBI ORF Finder(https://www.ncbi.nlm.nih.gov/orffinder/)對PacC開放閱讀框進行預測,利用在線分析網站ExPASy中的在線工具ProtParam(https://web.expasy.org/protparam/)對PacC編碼蛋白產物的相關理化性質預測[17],利用在線分析網站ExPASy ProtScal(https://web.expasy.org/cgi-bin/protscale/protscale.pl)程序,對PacC編碼產物的疏水性和親水性進行分析;利用在線網站TMHMM(https://services.healthtech.dtu.dk/service.php?TMHMM-2.0)對PacC相關跨膜結構域進行生物學特性分析;利用SignalP 5.0 Server(https://services.healthtech.dtu.dk/service.php?SignalP-5.0)工具對PacC信號肽進行預測[18];利用NetPhos3.1(https://services.healthtech.dtu.dk/service.php?NetPhos-3.1)在線分析工具對PacC磷酸化位點進行分析;利用在線網站WoLF PSORT(https://wolfpsort.hgc.jp)對PacC蛋白的功能進行亞細胞定位[19];通過在線網站Swiss-Modeling(https://swissmodel.expasy.org)對PacC的氨基酸序列進行蛋白質三維結構的同源建模,獲得其氨基酸序列,預測三級結構[20]。
1.3.6 不同pH值下酸腐病菌菌絲生長量的測定
參考Tan Xiaoli等[21]的方法,略有修改,于pH 1.0、2.0、2.2、2.5、3.0、4.0、5.0、6.0、7.0、8.0、9.0、10.0、11.0的PDB培養基中加入1 mL終濃度為106spores/mL酸腐病菌孢子懸浮液,每個處理3 瓶,置于28 ℃搖床(160 r/min)振蕩培養2 d。培養結束,離心收集沉淀,沉淀用無菌水重懸兩次,最后將沉淀置于-80 ℃超低溫冰箱冷凍,第2天進行真空冷凍干燥,分別稱量菌絲干質量。
1.3.7 果實接種
選用成熟度、大小一致的宮川蜜橘(Citrus unshiuMarc.,pH>3.0)和尤力克檸檬(Citrus limon(L.) Burm.F.,pH<3.0)作為材料,參考Li Lu等[22]的方法進行接種。蒸餾水清洗后在2% NaClO溶液中浸泡2 min,蒸餾水清洗3 次并自然晾干。用無菌手術刀在果實赤道部位劃2 個3 mm×3 mm×2 mm的傷口,放置1 h后,在傷口處接種10 μL酸腐病菌孢子懸浮液(1×107spores/mL),貯藏于28 ℃、相對濕度85%~90%的密閉培養箱中,每2 d測量果實發病部位pH值,每組樣品3 個平行,并將樣品用液氮速凍,保存于-80 ℃超低溫冰箱[23],用于后續RNA的提取。
1.3.8 實時聚合酶鏈式反應(real-time polymerase chain reaction,real-time PCR)分析
離體條件下菌絲的收集參考Yuan Xingxing等[24]的方法,略有修改,過濾收集在pH 3.0 PDB培養基中搖瓶培養24 h的柑橘酸腐病菌菌絲,取等量分別轉移至pH 1.0、3.0、5.0、7.0、9.0、11.0的PDB培養基中,搖瓶培養1 h后,收集菌絲,于-80 ℃保存,用于后續RNA的提取。
使用PhyZolTMTotal RNA Extraction Reagent提取離體條件培養以及腐爛果實的酸腐病菌菌絲總RNA,利用Hifair?III 1stStrand cDNA Synthesis SuperMix for qPCR逆轉錄試劑盒合成cDNA第1鏈。用cDNA模板檢測各基因表達水平,內參基因為β-actin,利用軟件Primer 5.0設計熒光定量引物,引物序列見表1,送至擎科(長沙)生物科技有限公司合成。使用Hieff UNICON?Universal Blue qPCR SYBR Green Master Mix試劑盒進行real-time PCR。反應體系:10.0 μL Blue qPCR SYBR Green Master Mix,上、下游引物各0.4 μL(10 μmol/L),1.0 μL模板,8.2 μL ddH2O。每個樣品進行3 次技術重復。反應條件:95 ℃預變性120 s;95 ℃ 10 s,60 ℃ 30 s,45 個循環,離體培養的樣品以pH 3.0處理為對照,果實樣品以發病2 d組織中病原菌基因轉錄水平作為對照,采用2-ΔΔCt法計算PacC在不同處理中的相對表達量[25]。
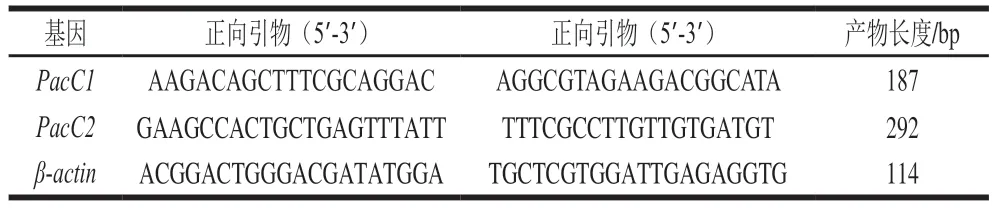
表1 real-time PCR引物序列Table 1 Primer sequences used for real-time PCR
1.4 數據處理與分析
實驗所得3 個平行數據用Excel軟件進行處理,采用Origin 2021軟件進行繪圖,利用SPSS 25.0采用單因素方差分析比較各組間的統計學差異,P<0.05認為存在顯著差異。
2 結果與分析
2.1 柑橘酸腐病菌PacC生物信息學分析
2.1.1 柑橘酸腐病菌PacC候選基因的篩選結果
根據柑橘酸腐病菌全基因組基因注釋信息篩選到2 個PacC候選基因PacC1和PacC2。它們與白地霉PacC基因同源性分別高達76.7%和74.7%,可能具備PacC相似功能,因此,后續對這兩個基因進行生物信息學和表達模式分析。
2.1.2 不同物種間PacC基因編碼氨基酸序列比對與保守基序分析
通過與釀酒酵母(Saccharomyces cerevisiae)、白色念珠菌(Candida albicans)、黑曲霉(Aspergillus niger)、指狀青霉(P.digitatum)、尖孢鐮刀菌(Fusarium oxysporum)等其他真菌的PacC基因編碼氨基酸序列進行對比,發現酸腐病菌PacC與上述比對真菌的蛋白結構相似,均含有3 個保守的Cys2His2鋅指蛋白結構域,屬于C2H2蛋白家族(圖1)。利用在線網站MEME并結合MEGA7.0和TBtools軟件預測上述真菌PacC的保守基序,共鑒定出10 個保守基序,其中Motif1、Motif2、Motif3、Motif7、Motif9高度保守。酸腐病菌PacC1與PacC2結構相似,具有相同的保守基序(Motif1、Motif2、Motif3、Motif4、Motif5、Motif6、Motif7、Motif9)(圖2)。

圖1 酸腐病菌PacC與其他真菌PacC的蛋白氨基酸比對Fig.1 Alignment of predicted amino acid sequence of PacC between G.citri-aurantii and other fungi

圖2 酸腐病菌PacC與其他真菌PacC保守基序分析Fig.2 Alignment of predicted conserve motif of PacC between G.citri-aurantii and other fungi
2.1.3 不同物種間的PacC系統進化分析
由系統發育樹可以看出,酸腐病菌的PacC1和PacC2親緣關系最近。測試物種中,與指狀青霉(P.digitatum)、產黃青霉(P.chrysogenum)以及黑曲霉(A.niger)的親緣關系較近,與釀酒酵母(S.cerevisiae)、白色念珠菌(C.albicans)等物種的親緣關系較遠(圖3)。
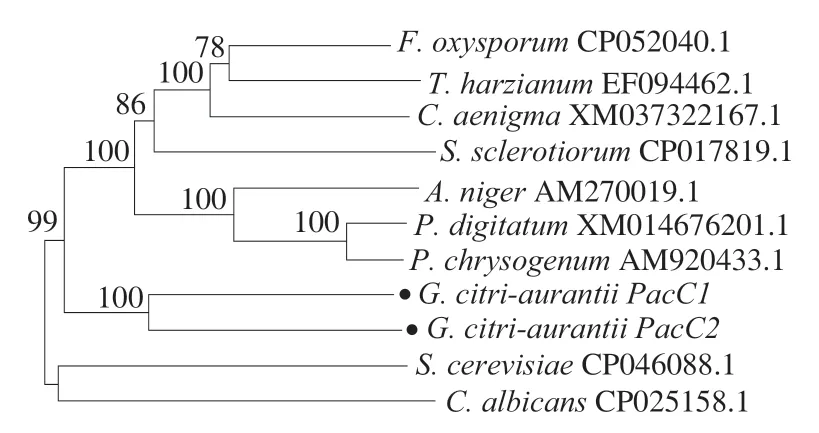
圖3 酸腐病菌PacC與其他真菌PacC的親緣關系Fig.3 Phylogenetic tree showing genetic relationship between G.citri-aurantii and fungi based on PacC
2.1.4 柑橘酸腐病菌PacC結構特征及染色體分布
利用在線網站GSDS 2.0與MG2C v2.1對PacC1與PacC2的結構與染色體分布進行分析。由圖4可知,PacC1與PacC2均只有外顯子結構,外顯子長度分別為1 872 bp和1 779 bp,均無內含子與UTR區域。根據圖5可知,PacC1與PacC2分別位于第3號染色體和第10號染色體上,第3號染色體長度為2 879 209 bp,PacC1位于1 832 639~1 834 510 bp之間。第10號染色體長度為1 531 806 bp,PacC2位于877 315~879 093 bp之間。

圖4 柑橘酸腐病菌PacC結構特征Fig.4 Structural characterization of PacC of G.citri-aurantii

圖5 柑橘酸腐病菌PacC染色體分布Fig.5 Chromosome distribution of PacC of G.citri-aurantii
2.1.5 酸腐病菌PacC蛋白結構與理化性質分析
由圖6可知,PacC1與PacC2的蛋白結構具有一定差異。對其理化性質進行分析,發現PacC1與PacC2均為親水性的不穩定蛋白[26],沒有跨膜位點與信號肽,WoLF PSORT亞細胞定位顯示二者均在細胞核內發揮功能調控作用。它們在分子質量、等電點、磷酸化位點等方面存在一定差異。其中,PacC1蛋白對應堿基數為1 872 bp,編碼623 個氨基酸,PacC1分子質量為67.80 kDa,等電點為6.71,可能發生磷酸化的位點有Ser56、Thr18、Tyr8;PacC2蛋白對應含1 779 個堿基,編碼592 個氨基酸,PacC2分子質量為64.32 kDa,等電點為8.50,Ser72、Thr18、Tyr13位點可能發生磷酸化(表2)。PacC1和PacC2蛋白高級結構預測見圖6。

圖6 PacC1(A)與PacC2蛋白(B)高級結構預測Fig.6 Prediction of protein structure of PacC1 (A) and PacC2 (B) of G.citri-aurantii

表2 酸腐病菌PacC1與PacC2理化性質比較Table 2 Comparison of physicochemical properties between G.citri-aurantii PacC1 and PacC2
2.2 不同pH值對酸腐病菌菌絲生長量和PacC表達量的影響
2.2.1 不同pH值對酸腐病菌菌絲生長量的影響
如圖7所示,酸腐病菌在pH 2.0~11.0范圍內均能生長,其中pH 3.0菌絲生長量顯著高于其他pH值,其次是pH 4.0。在pH 2.5和pH 5.0~10.0范圍內菌絲生長量沒有顯著差異。pH 2.2和11.0菌絲生長受到顯著抑制,在pH<2.2時幾乎不生長。由此可知,pH值對酸腐病菌的生長具有調控作用,酸腐病菌具有廣泛的pH值適應性,但其最適生長pH值約為3.0,這與許多柑橘品種果實pH值接近[27]。
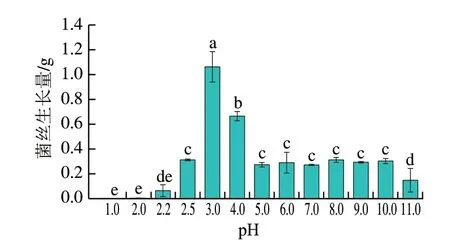
圖7 不同pH值對培養2 d酸腐病菌菌絲生長量的影響Fig.7 Effect of different pH on the growth of G.citri-aurantii mycelium cultured for two days
2.2.2 離體條件下不同pH值對PacC表達量的影響
以酸腐病菌最適生長pH值(pH 3.0)為對照,比較PacC1和PacC2在不同pH值的表達情況。結果表明,PacC1和PacC2的表達均受pH值調控,且兩個基因的表達變化趨勢具有一定相似性。處理1 h時,PacC1和PacC2表達均在pH 11.0顯著上調(P<0.05),相對表達量分別約為對照的2.24 倍和1.62 倍;PacC1表達量在其他pH值條件下沒有顯著差異,但PacC2在pH 1.0時幾乎不表達(圖8)。處理24 h時,PacC1和PacC2表達量在pH 9.0和pH 11.0均顯著上調,其中pH 9.0處理中的兩個基因表達量均約為對照的4.8 倍,PacC1在pH 11.0的表達量約為對照的8.5 倍,而PacC2的上調倍數更高,約為對照的13 倍,兩個基因在其他pH值的表達水平相似,但pH 1.0時PacC2的表達量顯著上調。

圖8 不同pH值對酸腐病菌PacC1與PacC2表達的影響Fig.8 Effects of different pH on the expression levels of PacC1 and PacC2 in G.citri-aurantii
2.3 PacC在酸腐病菌侵染柑橘果實侵染過程中的表達情況
宮川蜜橘果實人工接種酸腐病菌2 d時,果實傷口處可見明顯軟腐癥狀,病斑直徑為(1.97±0.13)cm,4 d出現白色霉層,病斑直徑達到(3.88±0.06)cm,至發病6 d時,果實幾乎完全腐爛(圖9A、B)。果實發病過程中腐爛部位的pH值呈下降趨勢,在6 d時下降約0.2,顯著低于對照(圖9C)。酸腐病菌兩個PacC的表達總體呈上調趨勢,但PacC1對pH值變化的響應更快,侵染4 d時表達量即顯著高于2 d,侵染6 d時表達量上調至2 d的3.9 倍,而PacC2的表達量在6 d才有顯著上調(圖9C)。尤力克檸檬果實人工接種酸腐病菌2 d時,果實傷口處也可見明顯軟腐癥狀,病斑直徑為(1.92±0.14)cm,4 d時軟腐面積加大,病斑直徑為(3.80±0.05)cm,至發病6 d時果實幾乎被白色霉層覆蓋,完全腐爛(圖10A、B),發病過程中腐爛部位的pH值呈上升趨勢,至接種6 d時上升約0.4,顯著高于對照(圖10C)。酸腐病菌兩個PacC的表達量均在侵染6 d時發生顯著上調(圖10C)。

圖9 酸腐病菌侵染過程中PacC的表達量與蜜橘果實發病部位pH值的關系Fig.9 Relationship between PacC expression and pH change of mandarin wounds infected with G.citri-aurantii

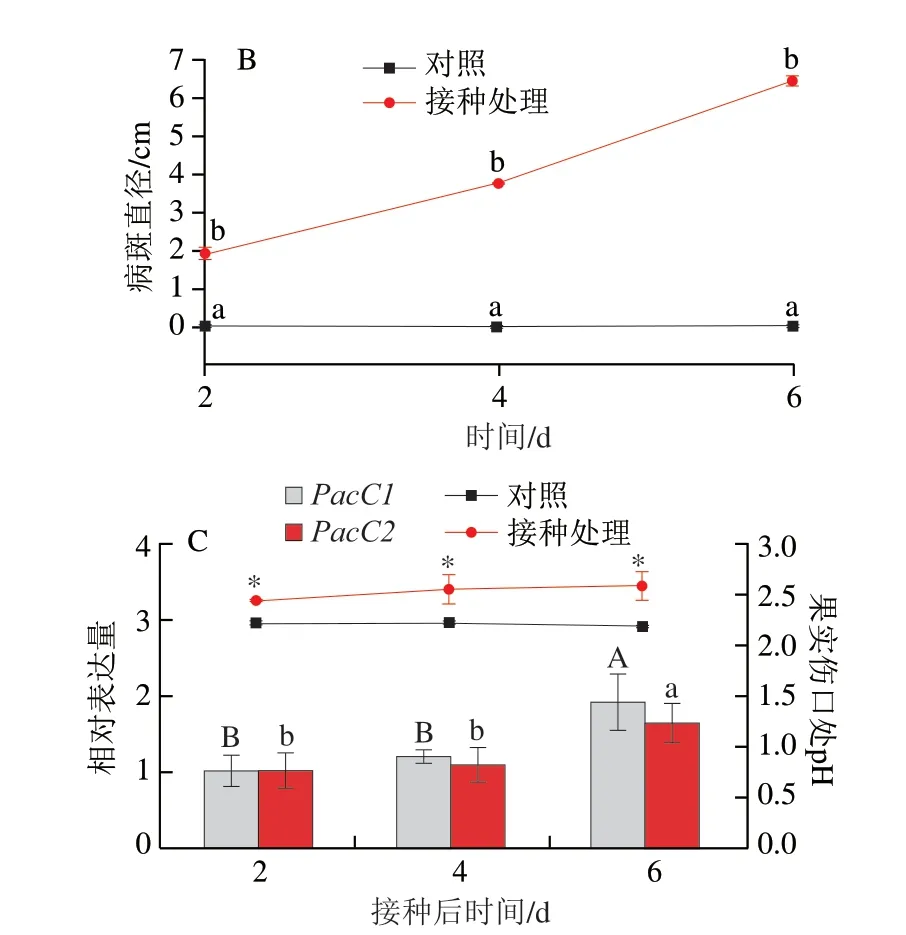
圖10 酸腐病菌侵染過程中PacC的表達量與檸檬果實發病部位pH值的關系Fig.10 Relationship between PacC expression and pH change of lemon fruit wounds infectedwith G.citri-aurantii
3 討論
采后病原真菌的侵染能力受寄主傷口、溫度、濕度和環境pH值等因素的影響,其中環境pH值對病原真菌的生長和致病力有重要調控作用[28]。許多病原真菌通過調整對微量元素的利用和體內一些蛋白質的功能應對環境pH值的變化,因此具有廣泛pH值適應性。目前,絲狀真菌中報道較多的pH值響應機制為pal途徑,該途徑包含7 個基因:pacC/pacl、palA、palB、palC、palF、palH和palI,其中,PacC作為重要的環境pH值調節因子,在微生物調節、適應環境的過程中發揮重要作用[9]。PacC的結構高度保守,PacC蛋白屬于C2H2蛋白家族,含有3 個保守的Cys2His2鋅指蛋白結構域[29-31]。采后致病真菌中,意大利青霉和指狀青霉PacC均具有此結構[9]。本實驗對酸腐病菌候選PacC的氨基酸序列和保守基序進行分析,發現2 個候選基因均具有PacC的保守結構,且系統發育樹也表明其親緣關系與柑橘采后致病菌指狀青霉較近,說明其可能與指狀青霉PacC具有類似功能。指狀青霉同樣是嗜酸真菌,在侵染柑橘果實過程中PacC表達水平呈顯著上調,且可能參與酸分泌過程改變環境pH值,PacC功能的缺失可導致病菌致病性顯著下降[9,32]。此外,指狀青霉PacC在離體堿性條件下有高水平表達,對指狀青霉的環境適應性有重要意義[32]。因此,推測酸腐病菌PacC在柑橘果實致病性和病原菌環境適應性中可能具有重要調控作用。進一步對酸腐病菌兩個PacC的結構和性質進行分析,發現PacC1與PacC2位于不同染色體(圖5),在等電點和磷酸化位點等方面有較大差異(表2),推測這兩個基因在功能上也存在一定差異。
不同的寄主組織存在顯著性的pH值差異,不同病原菌生長的適宜pH值及耐受范圍也存在較大差異[28]。陳彥等[33]研究發現葡萄白腐病菌(Coniella diplodiella)菌絲在pH 3.0~5.0條件下生長較為迅速,且最適pH值約為4.0;而王宏等[34]研究發現梨鏈格孢菌(Alternaria alternata)可在pH 4.0~12.0范圍內生長,最適pH值約為7.0~8.0。本實驗中,離體條件下,柑橘酸腐病菌具有較廣的pH值適應能力,但pH 3.0~4.0范圍的生長量顯著高于其他pH值(圖7),而其寄主pH值范圍大多約為2.0~6.0,其中檸檬類果實pH值約為2.2,寬皮柑橘類果實pH值約為3.0,甜橙類果實pH值約為5.0[35]。因此,推測酸腐病菌在寬皮柑橘類果實中具有更強的生長能力,而實際上,酸腐病菌對pH值極低的檸檬果實也有較強侵染力,這可能與其pH值調節能力有關。
大量研究表明,中性到堿性條件下,PacC在細胞核中調節基因的表達,這些基因參與病原真菌的細胞壁合成、跨膜運輸、細胞穩態、真菌毒力、菌絲生長、離子脅迫等方面[36-38]。如指狀青霉PacC在堿性條件下處理48 h時的表達量即上升為對照的8.4 倍;指狀青霉PacC缺失突變體在堿性環境中無法生長,對果膠的利用能力和柑橘的致病性也大幅下降[9]。但是PacC在酸性條件下的表達模式不盡相同。馮越等[39]研究發現,意大利青霉PacC在酸性環境中的表達量顯著下調。而指狀青霉PacC的表達不受果皮酸性環境的影響[9]。同樣,灰霉病菌PacC缺失突變體的致病性在組織pH值為酸性的寄主中也沒有變化[40]。本實驗中,離體條件下培養的酸腐病菌從最適生長pH 3.0培養基轉移到不同pH值培養基中繼續培養1 h時,PacC1和PacC2均能對環境pH值進行迅速響應,在顯著抑制菌絲生長的pH 11.0處理中表達迅速上調(圖8),培養24 h時,兩個PacC的表達量在pH 9.0和pH 11.0的處理中均顯著上調,且pH 11.0處理中表達量更高(圖8),說明堿性環境能誘導酸腐病菌PacC的高水平表達,這與上述采后致病菌PacC的表達情況一致。值得注意的是,PacC1和PacC2在某些酸性條件下的表達趨勢有顯著差異,如pH 1.0處理1 h,PacC2的表達被顯著抑制,同樣,PacC2的表達水平在pH 3.0處理24 h也顯著下降,而PacC1的表達未發生顯著變化(圖8),說明酸腐病菌PacC的表達也受酸性環境調控,可能在侵染柑橘果實過程中也有類似情況,而這兩個基因呈現出不同的響應模式可能與其結構和性質的差異相關。
在侵染初期,采后病原真菌通常分泌酸或堿以調節侵染位點pH值以加速侵染進程,這一過程可能與PacC的調控有關[41-43]。如炭黑曲霉(A.carbonarius)PacC缺失株侵染葡萄時不再分泌葡萄糖酸和檸檬酸[42];核盤病菌(Sclerotinia sclerotiorum)PacC直接調控草酸的分泌降低侵染位點pH 值[10]。膠胞炭疽菌(C.gloeosporioides)可在鱷梨組織中分泌氨,進而導致侵染位點pH值升高[44]。本實驗中,人工接種酸腐病菌的宮川蜜橘和尤力克檸檬果實侵染位點pH值的變化呈相反趨勢。宮川蜜橘果實初始pH值約為3.30,隨著病斑面積的擴大,發病部位的pH值逐漸下降至3.11,說明酸腐病菌與宮川蜜橘果實互作過程中的產物可導致果實腐爛部位pH值的下降,有利于病原菌的擴繁。接種后4~6 d果實病斑面積急劇增大,同時酸腐病菌PacC的表達量也上調,說明酸腐病菌PacC很可能參與對環境pH值的調控,從而促進侵染進程。PacC在酸腐病菌侵染尤力克檸檬果實過程中也發揮類似作用,但侵染位點pH值呈上升趨勢。雖然不同品種果實發病部位pH值的變化趨勢不同,但發病后期的組織pH值均更接近酸腐病菌生長的最適pH值。王哲[45]的研究發現,酸腐病菌侵染柑橘果實時分泌葡萄糖酸和另一種有機酸使侵染位點pH值下降,但酸腐病菌是否能通過分泌氨上調侵染位點pH值還有待證實。
4 結論
環境pH值對柑橘酸腐病菌的生長具有調控作用,同時,酸腐病菌也可以一定程度的調節環境pH值。酸腐病菌的2 個PacC均能在堿性條件下被誘導表達,但由于基因結構和理化性質的差異,它們對pH值的響應程度和方式也有一定差別。兩個PacC對于酸腐病菌廣泛的pH值適應性以及柑橘果實侵染進程具有重要作用。
