干酪乳桿菌刺激小鼠腸道的轉錄組學分析
2023-12-09 03:29:58程榮葉李樹東楊啟堯余麗蕓侯喜林
微生物學雜志 2023年4期
程榮葉, 李樹東, 秦 達, 楊啟堯, 李 穎, 余麗蕓*, 侯喜林*
(1.黑龍江八一農墾大學 生命科學技術學院,黑龍江 大慶 163319;2.黑龍江八一農墾大學 動物科學技術學院,黑龍江 大慶 163319)
干酪乳桿菌(Lactobacilluscasei)是一種哺乳動物胃腸道宿主菌,在過去幾十年中,因其益生特性在臨床和動物模型中被廣泛研究[1]。但是其干預腸道菌群,加強免疫反應的詳細機制還不清楚,為了探究這個問題,擬通過干酪乳桿菌口服實驗動物鼠,進行腸道的轉錄組學測序,分析發生顯著變化的表達基因,挖掘這些基因在刺激免疫細胞分化成熟等方面的可能通路,為揭示這個科學問題提供參考。近20年來,益生菌的研究取得了突破性進展,Toh等[2]研究報道了干酪乳桿菌作為對人類和家畜的一種益生菌,能夠穩定腸道微生物群落,減少病原微生物入侵,減輕或預防由細菌和病毒引起的腹瀉,從而使腸道運動障礙達到一種正常化的狀態。口服免疫后,干酪乳桿菌攜帶抗原物質附著在腸道和泌尿生殖系統中,刺激機體產生免疫反應,進一步阻礙病原微生物的入侵和黏附,減少了炎癥反應,并且干酪乳桿菌還可激活機體相關免疫細胞的活性,包括巨噬細胞、NK細胞和T淋巴細胞[3-4]。除此之外,干酪乳桿菌進入機體后還可以產生一些富含抑菌活性的多肽或前體多肽,抑制有害菌感染機體,促進腸道內益生菌的增殖,進而發揮益生作用[5]。重要的是重組乳酸菌增強了黏膜表面和血清中的lgA、IgM和IgG的水平,誘導產生TNF-α、IFN-γ等細胞因子,繼而加強體液免疫和細胞免疫[6-8]。特別是組學手段為深入研究提供了目標和方向,加快了益生菌機制的研究進程。Wang等[9]采用宏基因組和宏轉錄組測序揭示了干酪乳桿菌的體內表達模式,發現攝入的益生菌必須改變其轉錄模式才能在人類腸道中生存和適應,時間依賴性的激活模式研究表明益生菌和人類腸道微生物之間存在高度動態的串擾。轉錄組學確定了驅動嗜酸乳桿菌(Lactobacillusacidophilus)在腸道適應性和活性的可能基因,揭示了這種益生菌促進抗炎反應、維持腸上皮穩態和調節宿主晝夜代謝軸的作用[10]。同樣的高通量轉錄組分析揭示了芽胞桿菌(Bacillus)對雞的免疫、腸道屏障系統和代謝具有改善作用[11]。但是干酪乳桿菌在刺激腸道組織后產生的轉錄因子功能還沒有完全被揭示,有待進一步的驗證和研究。轉錄組學研究已經取得了一些成果,揭示了一些轉錄因子、調控蛋白的作用,例如脂氧合酶(Aloxe)3基因編碼的一種新型代謝調控蛋白-表皮型Aloxe3的發現[12];白細胞分化抗原6 (Cluster of differentiation 6,CD6)鑒定為富含半胱氨酸的清道夫受體糖蛋白超家族中的一員[13],在胸腺發育、T細胞活化和機體的免疫應答中均起著至關重要的作用[14-16]。在藥物代謝中,肝臟含有發揮重要作用的多種藥物代謝酶,例如羧酸酯酶(Carboxylesterase,CES )組成一個多基因家族,其基因產物多數分布在機體的各個組織和細胞的內質網中,這些酶以十分高效的催化去水解內源性和外源性物質[17]。ZBTB16基因屬于抑癌基因家族成員之一,最早發現于早幼粒細胞白血病的研究中,故稱之為早幼粒細胞白血病鋅指蛋白(ZBTB16或PLZF或ZNF145)[18],該基因可在人的肺組織中表達[19-21]。糖蛋白非轉移性黑色素瘤蛋白B(GPNMB)是一種跨膜糖蛋白,因其在黑色素瘤細胞系中高表達且轉移性低而得名,由于GPNMB在成骨細胞分化和增加骨礦物質沉積中的作用,也被稱為骨激活素[22-24]。OX40(CD134)及其配體OX40L(CD252)是TNF家族的成員,在CD4+、CD8+T細胞及其他幾種淋巴和非淋巴細胞上均有表達[25]。UBASH3A是蛋白絡氨酸磷酸激酶家族中的一員,包含三個結構域:N-末端UBA(泛素相關)、SH3(Src同源性3)和C末端組氨酸磷酸酶PGM結構域[26-27]。LZTS1亮氨酸拉鏈腫瘤抑癌基因1位于染色體8p22上[28-29]。研究發現Nanos1基因作為轉錄后阻遏子參與調節翻譯過程[30]。孕激素和脂肪Q受體7(PAQR7)是膜孕激素受體中不可或缺的一員[31]。血清和糖皮質激素誘導的蛋白激酶1(SGK1)被鑒定為一種促腫瘤基因[34-35],是蛋白激酶“AGC”亞家族的成員,與AKT激酶家族具有結構和功能的相似性,也稱為Akt[32-33]。TRIM46基因屬于三重基序(Tripartite Motif,TRIM)蛋白家族成員,也稱為GENEY,位于人染色體1q21上[36-37]。 這些基礎研究提示益生菌調節腸道微生物群落的同時,有可能引起腸道組織的某些轉錄因子發生變化,進而調節免疫細胞的活性。因此,本研究以干酪乳桿菌為實驗材料灌胃小鼠,取不同時間節點的腸道樣本進行高通量轉錄組學測序,利用NGS分析干酪乳桿菌刺激小鼠腸道的轉錄組數據,對不同時間節點的轉錄組數據通過GO功能富集和KEGG通路富集,對富集的相關功能的差異基因進行生物信息學分析,進一步利用實時熒光定量技術(qRT-PCR)驗證相關差異基因的表達。
1 材料與方法
1.1 材料
1.1.1 供試菌株 試驗用菌株干酪乳桿菌CICC6105(LactobacilluscaseiCICC6105)購自中國工業微生物菌種保藏中心,-80 ℃冰箱保存。
1.1.2 實驗動物 20只雌性 Balblc小鼠(SPF級)購自北京維通利華動物技術有限公司,為5~6周齡,實驗開始前在無菌環境中適應性飼養一周,自由飲食。
1.1.3 主要試劑及儀器設備 蛋白胨、牛肉膏、酵母膏、吐溫-80、瓊脂購自Sigma公司;葡萄糖、無水乙酸鈉、檸檬酸三胺、磷酸氫二鉀、硫酸鎂、硫酸錳、碳酸鈣購自國藥集團; SYBR Green I核酸染料和HiScript III Reverse Transcriptase逆轉錄試劑盒購自諾唯贊公司; Recombinant RNasin Ribonuclease Inhibitor購自Promega公司。實時熒光定量PCR儀(LightCycler?96, 瑞士羅氏生物公司);微量紫外分光光度計(Evolution 220, 美國 ThermoFisher 公司);電熱恒溫培養箱(DRP-9162, 上海森信實驗儀器有限公司);空氣浴震蕩器(HZQ-C, 哈爾濱市東聯電子技術有限公司);旋渦震蕩器(QL-861, 海門市其林貝爾儀器制造有限公司);臺式低溫冷凍高速離心機(A-14C-1EU, 德國 Sigma 公司)。
1.2 方法
1.2.1 試驗動物模型分組 選取20只體重相近狀態良好的5~6 周齡SPF級Balb/c小鼠,在相同飼養環境條件下隨機將其分為兩組,分別為對照組(NTC)、干酪乳桿菌組(TOA)。NTC組:用生理鹽水飼喂小鼠30 d,分別在第10天、20天、30天取3只小鼠的小腸組織;TOA組:用濃度為5×1011cfu/mL的干酪乳桿菌生理鹽水懸液200 μL飼喂小鼠30 d,分別在第10天、20天、30天取3只小鼠的小腸組織。
1.2.2 干酪乳桿菌懸液的制備 將凍存的干酪乳桿菌菌株從-80 ℃冰箱取出,取出后將凍存菌平板劃線接種于MRS固體培養基中,37 ℃培養12 h后,挑取單菌落在5 mL MRS液體培養基中37 ℃靜置培養過夜,再以2%(體積分數)的比例接種于100 mL進行擴大培養,5 000 r/min,4 ℃離心10 min收集菌體,菌體用生理鹽水洗滌,重復3次。然后進行平板細菌計數,再用生理鹽水將菌體稀釋為5×1011cfu/mL,對小鼠進行每天一次的灌胃。
1.2.3 小鼠腸道的提取 對各組小鼠進行12 h禁食,禁食后,利用頸椎脫臼法處死小鼠,隨后將處死的小鼠放置濃度為75%的酒精中消毒,在無菌的超凈工作臺解剖小鼠,剪取胃幽門至回盲部的小腸,放入含有生理鹽水的平皿中,使用含有生理鹽水的注射器對小腸中的內容物進行洗滌,重復3次,洗滌后將小腸放入凍存管中。將凍存管放置液氮中速凍30 min,隨后將小腸保存至-80 ℃冰箱中,待整體試驗取樣完成后,將小腸用干冰運輸送至上海派森諾生物技術有限公司。
1.2.4 差異基因分析 小腸樣本經測序公司測序后返回,測序 Raw counts 數據采用 DESeq2 分析,得到顯著差異的基因,將篩選條件設為Pvalue≤0.05,且差異倍數|Fold change|>1 。隨后將表達差異分析結果進行可視化,通過熱圖可視化體現樣本間差異表達情況。
1.2.5 差異基因的GO和KEGG富集分析 為了研究干酪乳桿菌刺激小鼠腸道后產生的差異基因功能,利用派森諾平臺(https://www.genescloud.cn/login)對篩選出的差異基因做基因本體分析(GO)和KEGG通路分析,差異基因顯著富集結果利用柱狀圖對其進行可視化。
1.2.6 基本特性分析 使用在線網站ExPASy(https://web.expasy.org/protparam/)對14個基因分子量、等電點、不穩定系數和脂溶系數進行分析;利用Cell-PLoc 2.0(http://www.csbio.sjtu.edu.cn/bioinf/Cell-PLoc-2/)在線網站對基因編碼蛋白進行亞細胞定位分析;使用NCBI的Conserved-Domains數據庫對14個差異表達基因進行蛋白的功能結構域預測分析。
1.2.7 染色體定位分析 從NCBI數據庫中獲得的小鼠基因組gff文件,利用Tbtools軟件對14個差異表達基因進行染色體定位分析。
1.2.8 基因結構與保守基序分析 使用在線網站MEME (http://meme-suite.org/tools/meme)對14個基因的保守基序進行預測;使用Gene Structure Dispaly Server(http://gsds.gao-lab.org/)在線分析基因結構,得出基因中CDS (Sequence coding for amino acids in protein)分布情況。
1.2.9 qRT-PCR表達分析 利用NCBI primer designer,以14個差異表達基因的核苷酸序列為模板設計熒光定量引物,擴增長度控制在80 ~ 150 bp以內,SDHA作為內參基因,引物合成由生工生物工程(上海)股份有限公司完成(表1)。qRT-PCR反應時以反轉錄的第一鏈cDNA稀釋10倍作為模板。反映體系參照諾唯贊公司的HiScript III Reverse Transcriptase說明書,利用 Lightcycler 96 Real-time PCR System進行 qPCR。所有實驗數據按照平均數±標準方差計算;數據分析均使用 Graphpad prism 9.0 軟件進行;組間顯著性差異使用 Student′st-test 法進行分析;P< 0.05 代表組間顯著(*P< 0.05, **P< 0.01 和 ***P< 0.001, ****P< 0.000 1)。
2 結果與分析
2.1 表達差異基因分析
為了分析處理組與對照組之間差異基因的表達情況,對D10NTC vs.D10TOA 組、D20NTC vs. D20TOA組、D30NTC vs. D30TOA 組的差異基因利用 Deseq2 包對原始 counts 數據進行分析,閾值設置為P≤ 0.05 和|log2(Fold change)|>1,結果發現D10NTC vs. D10TOA 組、D20NTC vs. D20TOA組、D30NTC vs.D30TOA 組中分別有176、32、261個基因發生變化。其中在 D10NTC vs.D10TOA 組有 59 個基因上調與 117 個基因下調;D20NTC vs. D20TOA 組有 23 個基因上調與 9 個基因下調;D30NTC vs. D30TOA 組有 181 個基因上調與 80 個基因下調;利用熱圖對上述差異基因進行可視化(圖1)。
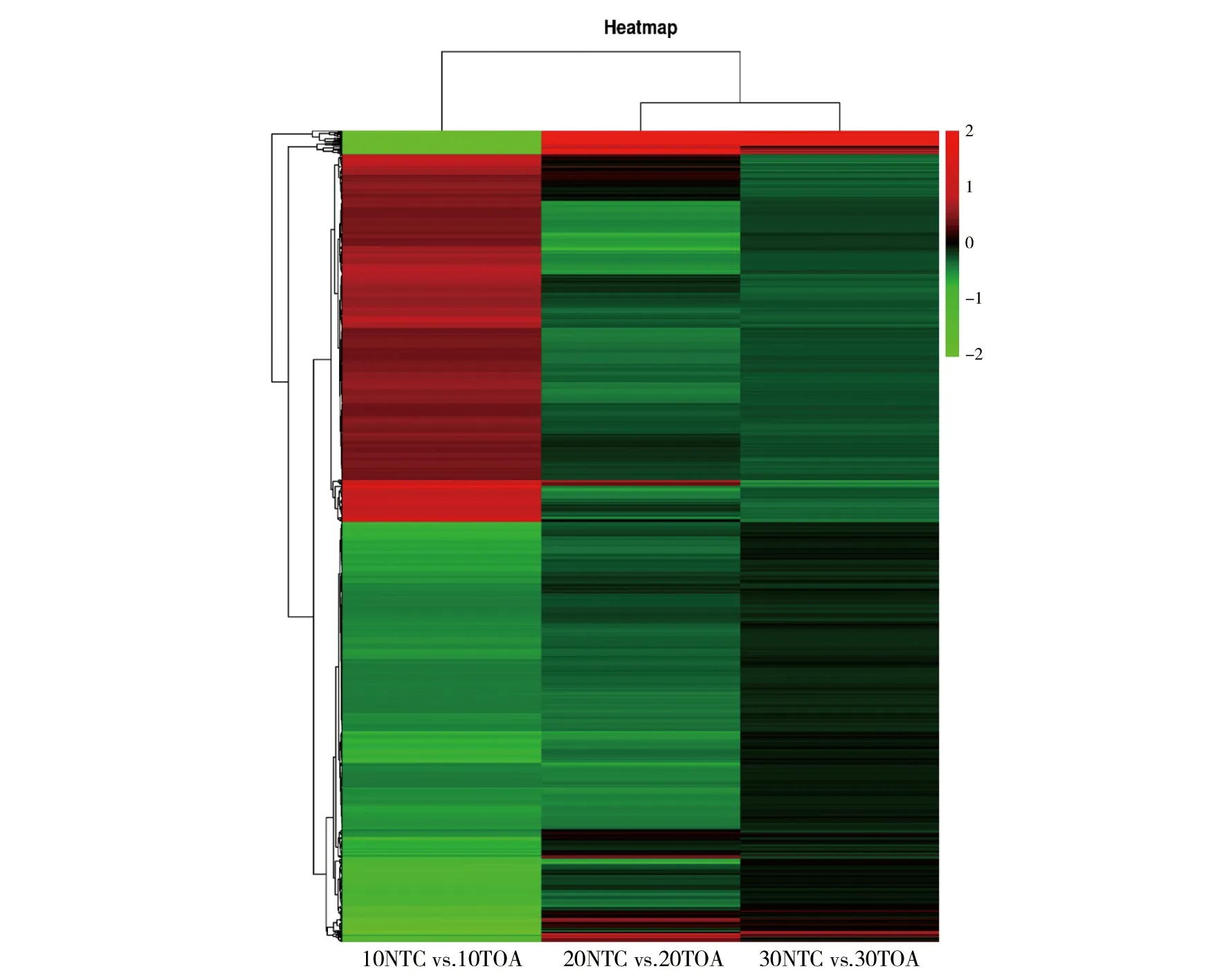
圖1 各組之間差異基因表達熱圖Fig.1 The heat map of differential gene expression among groups
2.2 GO功能富集分析結果
對差異基因的生物進程進行GO功能富集分析,干酪乳桿菌組(TOA)和生理鹽水組(NTC)的GO富集分析結果(圖2A、圖2B、圖2C):D10NTC vs. D10TOA 組表達的差異基因主要富集在免疫系統負調控、細胞分化、細胞黏附的調節、T細胞激活等進程;D20NTC vs. D20TOA 組表達的差異基因主要富集在淋巴細胞活化、調節細胞分化進程和T細胞激活等進程;D30NTC vs. D30TOA組表達的差異基因主要富集在免疫反應調節過程、免疫系統調節正調控、T細胞激活等進程。以上分析結果發現與細胞免疫相關的功能基因有Gpnmb、Hist1h4c、Zbtb16、Hist2h4、Sgk1、Aloxe3、Cspg5、Lzts1、Paqr7、Trim46、Ces1d、Nanos1、Cd6、OX40,故后續實驗將選取這14個基因進行驗證。

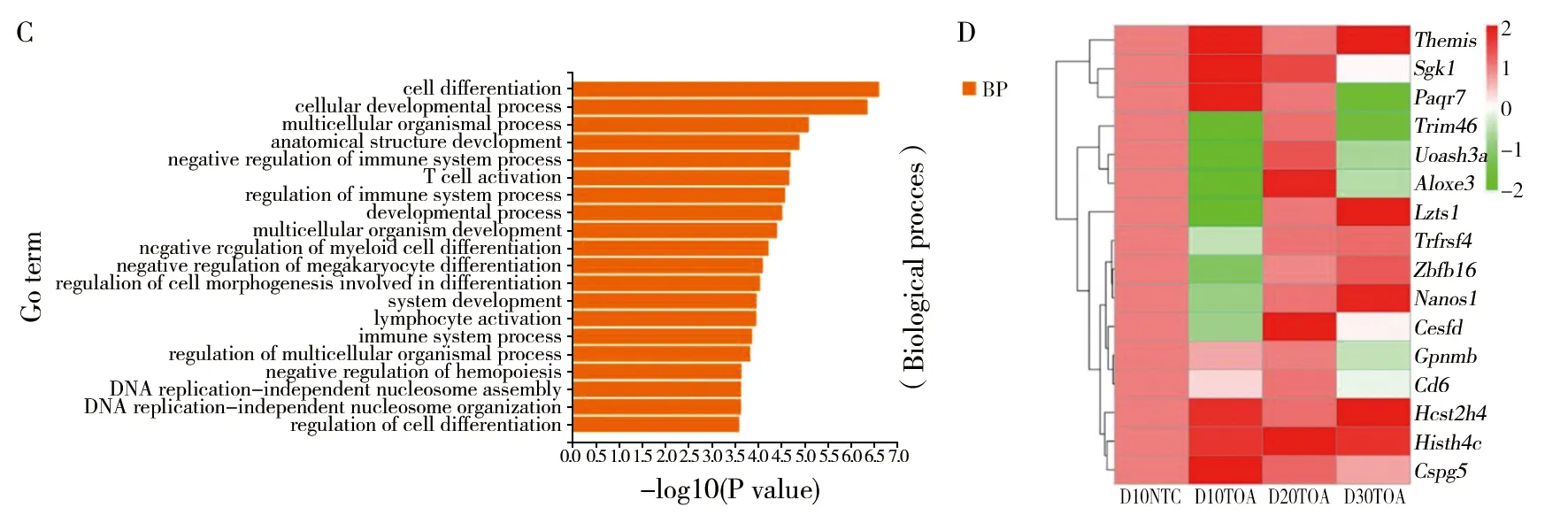
圖2 干酪乳桿菌組與空白組GO功能富集柱狀圖Fig.2 Histogram of GO function enrichment in Lactobacillus casei group and blank groupA: D10NTC vs.D10TOA組柱狀圖; B: D20NTC vs. D20TOA組柱狀圖; C: D30NTC vs. D30TOA組柱狀圖;D:14個差異表達基因熱圖 A: D10NTC vs. D10TOA; B: D20NTC vs. D20TOA; C: D30NTC vs. D30TOA;D: Heat map of 14 differentially expressed genes
2.3 KEGG富集分析結果
對差異基因進行KEGG通路富集分析,干酪乳桿菌組(TOA)和生理鹽水組(NTC)的KEGG分析結果(圖3):D10NTC vs. D10TOA 組的通路主要富集在PPAR信號通路、半乳糖代謝、脂肪的消化和吸收類固醇激素的生物合成等(圖3A);D20NTC vs. D20TOA 組主要富集在皮質醇的合成和分泌、脂肪細胞因子信號通路、膽固醇代謝、煙酸和煙酰胺代謝等(圖3B);D30NTC vs. D30TOA 組主要富集在細胞黏附分子(CAM)、B細胞受體信號通路、趨化因子信號通路、C型凝集素受體信號通路等(圖3C)。
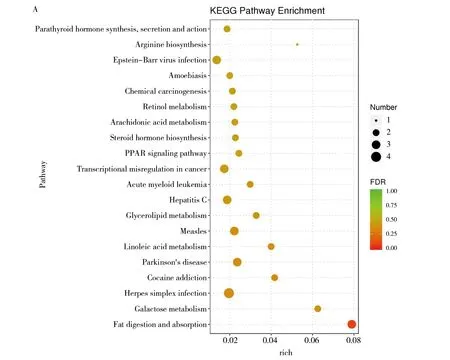
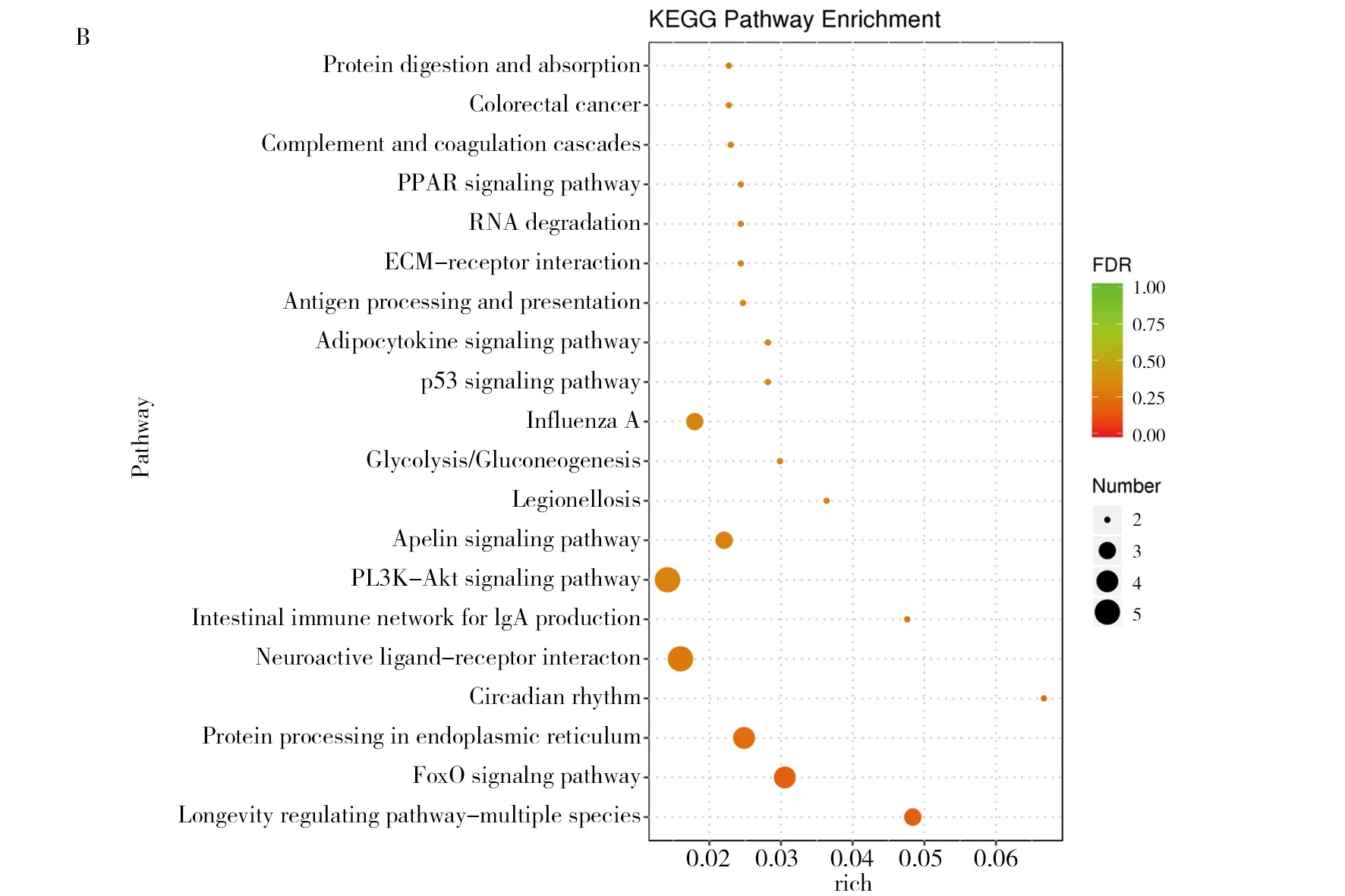
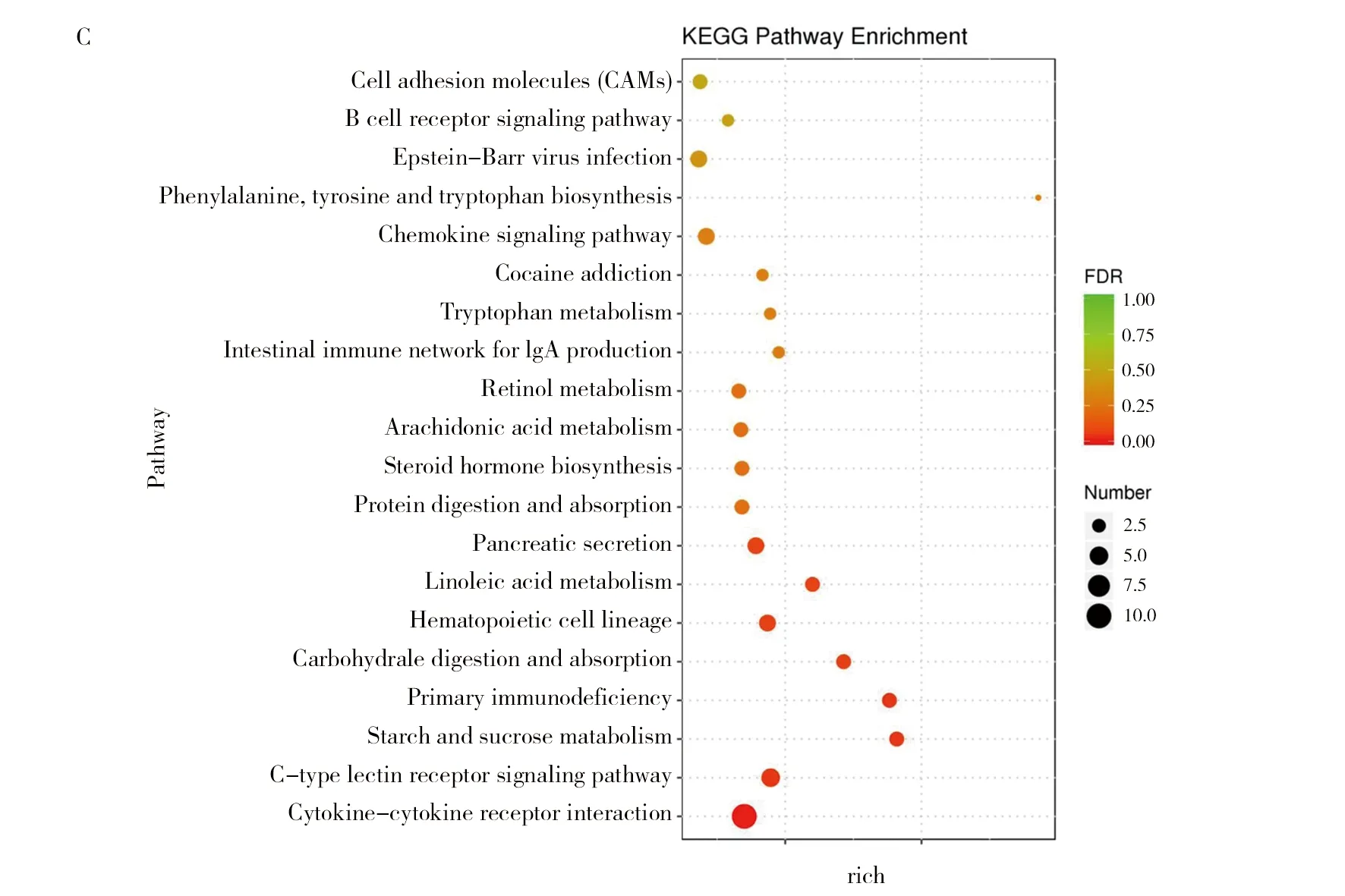
圖3 干酪乳桿菌組與空白組KEGG通路富集氣泡圖Fig.3 The enrichment Bubble Diagram of KEGG pathway in L. casei group and blank groupsA:D10NTC vs.D10TOA;B:D20NTC vs.D20TOA;C:D30NTC vs.D30TOA
2.4 基本特性分析結果
供試的14個差異表達基因的蛋白質長度為103~759 aa,分子量在11.37~83.45 kDa之間,其中ENSMUSG00000042766(Trim46)編碼的蛋白長度最大為759 aa,ENSMUSG00000060678(Hist1h4c)和ENSMUSG00000091405(Hist2h4)蛋白最小為103 aa;等電點pI在4.42~11.36之間,其中ENSMUSG00000060678(Hist1h4c)和ENSMUSG00000091405(Hist2h4)的pI最大,ENSMUSG00000032482(Cspg5)的pI最小;亞細胞定位預測表明,它們分別定位在細胞質、溶酶體、高爾基體、內質網、細胞外和細胞核內,均為不穩定的脂溶性蛋白,且均含有功能結構域(表2)。
2.5 染色體分析結果
14個差異表達的基因分布于小鼠染色體的3、4、6、8、9、10、11、13、和19號染色體上,其中Trim46和Hist2h4基因位于3號染色體,Paqr7和OX40基因位于4號染色體,Gpnmb基因位于6號染色體上,Lzts1和Ces1d基因位于8號染色體上,Zbtb16和Cspg5基因位于9號染色體上,Sgk1基因位于10號染色體上,Aloxe3基因位于11號染色體上,Hist1h4c基因位于13號染色體上,Cd6和Nanos1基因位于19號染色體上,從分布結果可以看出基因在染色體分布較為分散,沒有出現聚集并形成基因簇的現象(圖4)。
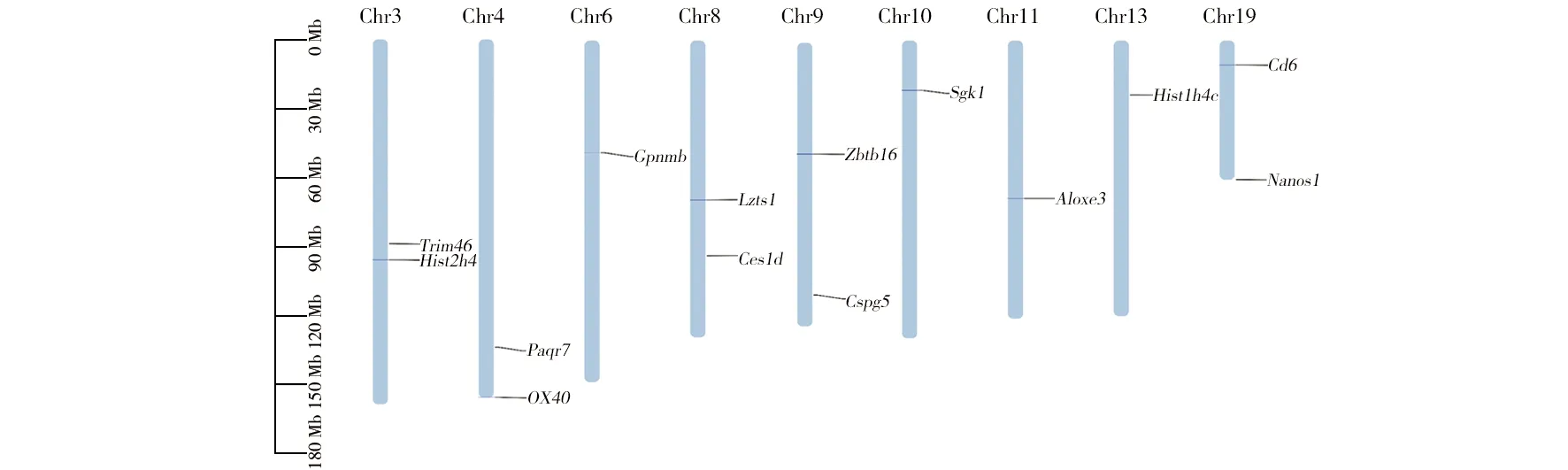
圖4 染色體定位Fig.4 Chromosomal localization
2.6 基因結構與保守基序分析結果
基因上的保守基序Motif分布稀疏,其中Hist2h4和Hist1h4c的保守基序分布及種類相似度高,保守基序模型較為保守。其余基因保守基序相似性不高,表明它們之間不具有類似的功能(圖5 A)。Hist1h4c、Hist2h4不含CDS序列,Gpnmb、Zbtb16、Aloxe3和Ces1d含有一個CDS序列,Sgk1、Cspg5、Paqr7、Trim46、Nanos1、Cd6和OX40含有2個CDS序列,Lzts1基因均含有3個CDS序列 (圖5 B)。

圖5 基因結構、保守基序可視化分析Fig.5 Visual analysis of gene structure and conserved motifsA:基因結構; B:保守基序A:Gene structure; B:Conserved motif
2.7 qRT-PCR分析結果
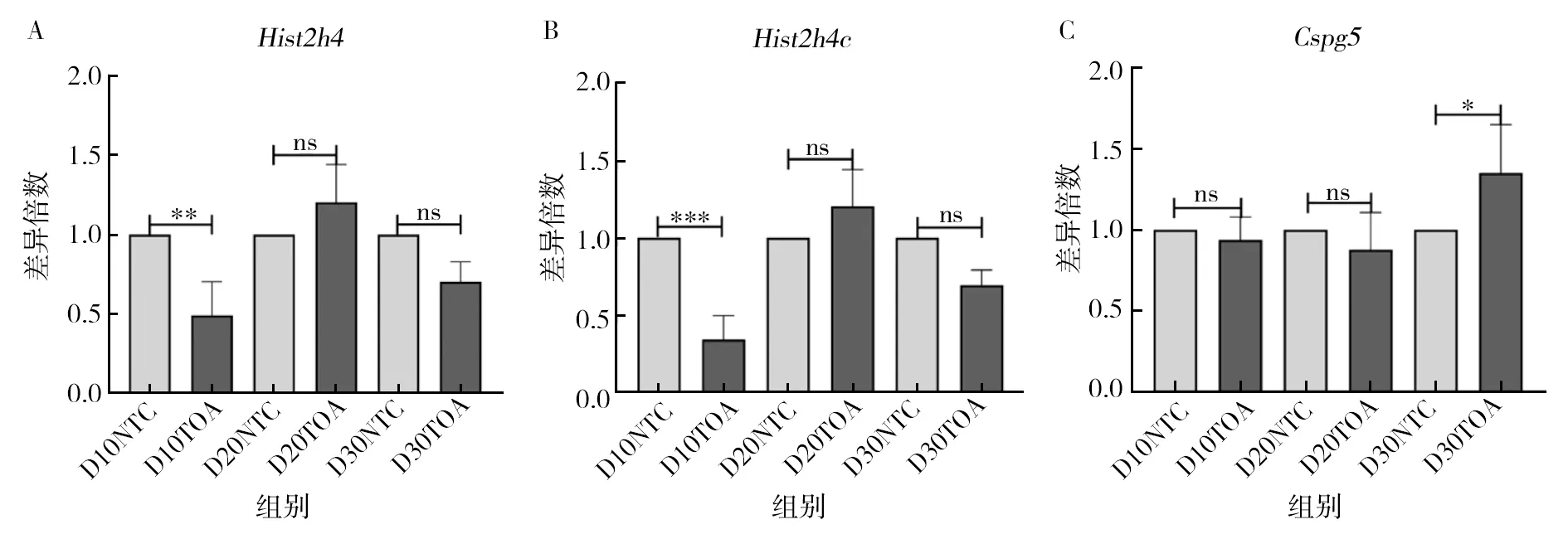
為驗證 RNA-seq 分析結果,利用實時熒光定量技術(qRT-PCR)對篩選出來的14個基因進行驗證,分析在不同時間差異基因的表達水平,結果發現,其中Hist1h4c、Hist2h4、Cspg5、Cd6、Gpnmb、Trim46和Nanos1基因的表達模式一致,在不同時間處理下的整體表達水平都比較低,組間表達不顯著。Sgk1、Ces1d、Lzts1、Paqr7、Aloxe3、Zbtb16和OX40整體表達水平較高,表達峰值明顯與轉錄組數據相符。其中Sgk1基因在第20天和第30天有顯著差異,Ces1d、Lzts1、Paqr7、Aloxe3、Zbtb16基因在第10天和第20天有顯著差異,OX40基因在第10天、第20天和第30天均有顯著差異(圖6)。
3 討 論
Jung等[38]發現干酪乳桿菌能預防病原體感染、對機體有保護和預防癌癥等作用。Devi等[6]發現干酪乳桿菌進入機體后,刺激黏膜激活機體免疫系統,影響特異性免疫應答能力,促進T、B細胞的增殖和成熟。腸道屬于最早接觸環境因素(食物、毒素和病原體等)的主要器官之一,具備近端到遠端的黏膜結構梯度和營養處理能力[39]。干酪乳桿菌對腸道的研究具有重要的意義,但是目前干酪乳桿菌刺激小鼠腸道,其對免疫細胞的影響還不清楚,因此,我們的研究旨在揭示干酪乳桿菌干預腸道后什么基因變化,能夠激活T細胞,致使T細胞增殖;T細胞激活對黏膜免疫有什么樣的作用效果?本研究通過RNA-seq測序探究了干酪乳桿菌刺激小鼠腸道后基因表達譜的變化,發現7個基因功能主要與免疫系統過程、固有免疫反應、T細胞激活等相關,為進一步深入研究奠定了基礎。
在建立干酪乳桿菌刺激小鼠腸道模型中,發現干酪乳桿菌飼喂10 d時,飼喂組對比對照組的上調基因數量比下調基因少,飼喂到20 d、30 d時,上調基因數量相比下調基因的數量增多。這種現象的原因可能是飼喂干酪乳桿菌10 d后,小鼠腸道內菌群的變化還沒有趨于穩定,有些作用還沒有顯現。但是繼續飼喂干酪乳酸菌之后,隨著飼喂時間的增加,小鼠可能產生耐受反應,小鼠腸道內益生菌群趨于穩定,有些基因開始上調。干酪乳酸菌在小鼠腸道中為優勢菌屬,刺激小鼠腸道細胞上調相關基因,類似的研究報道表明,當飼喂益生菌之后,雞腸道上調基因數目相比下調基因數目增多[40],但具體機制需要進一步深入研究。對差異基因進行生物進程富集分析,顯示差異基因多數富集在T 細胞活化、免疫調節負調控、細胞分化和先天免疫應答的調節上。KEGG通路富集顯示和PPAR信號通路、半乳糖代謝、B細胞受體信號通路、趨化因子信號通路、C型凝集素受體信號通路相關。以上結果進一步說明了干酪乳桿菌刺激小鼠腸道可能通過促進T細胞激活、細胞分化等以增強免疫系統能力。Lian等[41]研究結果表明抑制CES1(Ces1d的人類同源基因)可能是預防和治療非酒精性脂肪肝(NAFLD)的一個新的藥理學靶點。Kawaue等[42]研究證明Lzts1可作為細胞動力學的主要調節器發揮作用,在進化過程中增加了大腦結構的復雜性。Tan等[43]證明PAQR7可能介導孕酮誘導BPBC EMT逆轉,由孕酮激活的PAQR7與小窩蛋白-1相互作用(Cav-1)或調節表皮生長因子的活性受體(EGFR),使PI3K/Akt信號通路失活,最終抑制BPBC中的EMT。Higgins等[44]研究發現Aloxe3作為一種潛在的新的肝細胞禁食反應效應器,利用PPARy介導和多效性效應來增強肝臟和整個宿主的代謝,作為改善代謝性疾病的一個有希望的靶點。對于顯著表達的PLZF基因(早幼粒細胞白血病鋅指蛋白),有研究發現其對Na?ve T細胞的正常發育起著至關重要的作用[45-47]。 Zhang等研究發現,OX40調節T細胞的增殖,主要在后期對維持細胞存活發揮著至關重要的作用[48-49]。以上基因在乳酸菌刺激腸道后表達也顯著上升,和腸道黏膜免疫有什么相關性還有待于進一步驗證。
本研究用干酪乳桿菌刺激小鼠腸道,轉錄組測序的生物信息學分析發現了與T細胞激活相關的功能基因,同時也發現細胞黏附分子(CAM)、B細胞受體信號通路發生了顯著變化,初步表明干酪乳桿菌對機體的免疫過程發揮著重要的作用,為后續深入詳細研究干酪乳桿菌對免疫系統的作用機制提供參考。
猜你喜歡
興趣閱讀·興趣作文與閱讀(低年級)(2025年8期)2025-08-18 00:00:00
民用飛機設計與研究(2020年4期)2021-01-21 09:15:02
學苑創造·A版(2020年9期)2020-10-13 09:41:02
電子制作(2018年18期)2018-11-14 01:48:24
小學生學習指導(低年級)(2017年10期)2017-10-10 01:00:05
山東工業技術(2016年15期)2016-12-01 05:31:22
中國中醫藥現代遠程教育(2014年11期)2014-08-08 13:23:44
云南中醫學院學報(2014年3期)2014-07-31 18:57:34
終身教育研究(2014年5期)2014-02-28 01:23:06
七彩語文·畫刊(2012年3期)2012-04-29 00:00:00
