抗寒鍛煉中紫花苜蓿非結構性碳水化合物轉化轉運與抗寒性關系
2023-12-08 15:30:42趙金梅殷國梅孫娟娟衛媛李薇郭茂偉劉思齊張佳琪
草業學報 2023年12期
趙金梅,殷國梅,孫娟娟,衛媛,李薇,郭茂偉*,劉思齊,張佳琪
(1. 中國農業科學院草原研究所,內蒙古 呼和浩特 010010;2. 農業農村部牧草資源與利用重點實驗室,內蒙古 呼和浩特 010010;3. 內蒙古自治區農牧業科學院,內蒙古 呼和浩特 010031)
紫花苜蓿(Medicago sativa)為優良多年生豆科牧草,在全球廣泛種植[1]。我國北方氣候干燥寒冷,降水量少,適合苜蓿生長,是我國苜蓿主產區[2],但同時也是苜蓿凍害發生的主要地區,抗寒性成了該地區苜蓿品種選擇時考慮的關鍵因素。苜蓿在北方秋季日照時間縮短、溫度降低的氣候條件下,抗寒性會大幅提高。植物暴露在結冰以上低溫(一般為0~15 ℃)條件下一定時間后,抗寒性大幅度提高的過程稱為抗寒鍛煉(cold hardening, CH)[3]。在抗寒鍛煉過程中,植物體內會積累大量的滲透調節物質和抗氧化物質,如可溶性糖、淀粉等非結構性碳水化合物[4],使植物器官免受由低溫所產生的傷害,從而提高植株的抗寒性[5]。苜蓿根冠經過抗寒鍛煉后半致死低溫(semi-lethal low temperature, LT50)可以達到-20 ℃以下[6-7],在此過程中體內可溶性糖累積量明顯提高[7]。研究發現,抗寒性強的苜蓿品種秋季主根中可溶性糖含量較高,其含量與苜蓿再生率呈正相關[8]。Castonguay 等[9]指出抗寒的苜蓿品種在夏末到秋季末,根冠可溶性糖幾乎呈線性增加,其含量在100 mg·g-1以上[10],是冬季受凍害嚴重苜蓿品種可溶性糖含量的2 倍(50 mg·g-1)。淀粉為非結構性碳水化合物的重要成分之一,其在根冠中含量隨苜蓿抗寒鍛煉表現為先快速降低,然后緩慢降低,與可溶性糖含量變化有一定的互補性[7]。以上研究均表明,可溶性糖和淀粉在苜蓿抗寒性形成中起重要作用,同時在苜蓿抗寒鍛煉中存在可溶性糖與淀粉間的轉化。
植物在刈割或利用后再生初期,會將殘留器官中貯藏的非結構性碳水化合物轉化成可利用形態,用于地上部分生長,隨著地上部器官的再生和光合功能的恢復,在供應地上部繼續生長的同時將部分有機同化物運輸到根、根頸等器官并轉化為貯藏物質[11]。苜蓿1 年可以多次刈割利用,在無低溫脅迫的條件下,苜蓿根與根頸中碳水化合物總量隨秋眠級增加而增加,刈割后再生量也隨之增加[12],秋季刈割后,在溫度降低、日照時間變短的條件下,不同苜蓿品種再生高度不同,即苜蓿品種的秋眠級不同[13],同時苜蓿品種抗寒性和越冬性能也不同,因此推測刈割后苜蓿地上、地下器官中非結構性碳水化合物存在用于再生與提高越冬器官抗寒性之間的分配問題及品種差異。故需要對苜蓿秋季刈割后再生期間越冬器官-根與葉中非結構性碳水化合物的轉化、不同器官間的轉運及與苜蓿抗寒性的關系進一步研究。據此本研究模擬秋季抗寒鍛煉過程中溫度和光照變化,以2 個不同抗寒性苜蓿品種為對象,研究抗寒鍛煉過程中苜蓿根、葉中不同非結構性碳水化合物的動態變化、轉化、不同器官間的轉運和品種間的差異,解析非結構性碳水化合物利用模式與苜蓿抗寒性的關系,為苜蓿種質資源鑒定和抗寒性評價提供科學參考。
1 材料與方法
1.1 試驗材料
試驗材料為抗寒性強弱不同的2 個紫花苜蓿品種,其中:肇東(Zhaodong)苜蓿的抗寒性強,秋眠級1~2[8],由國家種質牧草中期庫提供;賽迪10(Sardi 10)的抗寒性較弱,秋眠級10 級,由百綠(天津)國際草業有限公司提供。
1.2 試驗地點與培養方法
試驗在中國農業科學院草原研究所實驗室內進行。于2019 年6 月25 日將肇東和賽迪10 苜蓿種子播種于塑料花盆中(口徑6 cm×6 cm,高11 cm),每個品種種植45 盆。盆中填裝營養土與蛭石的混合土,比例為3∶1。播種后花盆放置在培養箱(Percival LT-36VL,美國)中,在光照強度 300 μmol·m-2·s-1、光照時間13 h 和晝夜溫度25 ℃/18 ℃的條件下培養。待幼苗長出一片真葉時定苗,每盆保留3 株。生長至初花期將其刈割,再生7 d 后開始采集處理樣品。
1.3 試驗處理
2 個苜蓿品種植株均分為2 組,分別為抗寒鍛煉組(cold hardening group, CH)和正常條件生長組(normal growing group, NG),CH 組設3 個處理,分別為對照(CK)、抗寒鍛煉初期-降溫處理(CH1)和抗寒鍛煉后期-低溫處理(CH2)。再生7 d 后(處理前)為CK,接受抗寒鍛煉處理的CH1處理,初始晝夜溫度為25 ℃/18 ℃,光照時間13 h,晝夜溫度按照1 ℃·d-1,日照時間按照15 min·d-1遞減,處理7 d,處理結束時晝夜溫度為18 ℃/11 ℃,日照時間為11.25 h;CH2處理植株在接受CH1處理后繼續進行抗寒鍛煉處理,處理的初始晝夜溫度從18 ℃/11 ℃直接降到10 ℃/6 ℃,然后按照1 ℃·d-1的速率降溫,日照時間從11.25 h 開始,按照15 min·d-1遞減,處理7 d,夜間溫度降到1 ℃時不再降低,處理結束時晝夜溫度為3 ℃/1 ℃,日照時間為9.50 h。NG 組植株再生7 d 后仍在前述培養條件下生長。
1.4 取樣方法
CH 組植株分別在處理前(CK)、CH1處理后和CH2處理后采集葉片和根系樣品,NG 組分別在再生7、14、21 d取樣,2 組取樣時間序列相對應。每組每個處理3 個重復,各重復樣品取自同一處理的不同盆中植株。采集葉片樣品時,取各處理組植株頂端第3~6 個完整展開的葉片,一部分立即進行LT50測定,另一部分采樣現場用液氮快速冷凍后保存于-80 ℃超低溫冰柜中,用于可溶性糖和淀粉的測定。采集根樣品時,將根與營養土從培養缽中倒出,取出完整根系,流水沖去根表面土壤和其他物質,然后用去離子水沖洗3~5 次,用濾紙吸干根表面水分,一部分立即進行LT50測定,另一部分置于烘箱中105 ℃殺青30 min,65 ℃烘48 h,并粉碎備用。
1.5 測定指標與方法
1.5.1 根、葉半致死低溫的測定 參照梁莉等[14]的方法測定2 個品種苜蓿根、葉LT50,但略有改動,具體為先將梯度降溫盒按一定比例加入預冷的異丙醇和常溫異丙醇,達到不同設定的溫度。然后將所取完整葉片和切成段的主根置于凍存管中,凍存管置于不同溫度的梯度降溫盒中,每個梯度降溫盒中加入一枚紐扣溫度計,準確記錄盒中溫度。將梯度降溫盒置于冷藏柜中,放置2 h 后取出樣品,置于4 ℃冷藏過夜。過夜后的樣品置于加有25 mL 去離子水的離心管中,振蕩4 h,測定電導率C1,然后沸水中煮30 min,冷卻到室溫,振蕩4 h,測定總電導率C2。空白對照試管中加25 mL 去離子水,不加樣品,與其他樣品一起振蕩、煮沸并分別測定電導率,為Cck1、Cck2。離子滲透率計算公式如下:離子滲漏率(%)=(C1-Cck1)×100/(C2-Cck2)。不同處理溫度下離子滲漏率數據通過Logistic 方程進行擬合,在相關性顯著情況下,計算拐點溫度,即LT50[15]。
1.5.2 可溶性糖和淀粉含量測定 采用蒽酮-硫酸法測定可溶性糖和淀粉含量[16],采用酸水解的方法降解提取可溶性糖后的殘渣,然后通過蒽酮-硫酸法以葡萄糖為標準物質進行測定[16],得到的葡萄糖含量乘以換算系數0.9 即為淀粉含量。
1.6 數據計算與統計分析
使用SAS 軟件ANOVA 程序進行單因素方差分析和多重比較,采用Sigmaplot 軟件作圖。結果以平均值±標準誤表示。
2 結果與分析
2.1 抗寒鍛煉過程中不同紫花苜蓿品種根、葉抗寒性變化
CH1的2 個苜蓿品種葉和肇東根的LT50顯著低于CK(P<0.05),賽迪10 根的LT50高于CK(表1)。CH2處理2 個苜蓿品種根、葉的LT50均顯著低于CH1(P<0.05),在抗寒鍛煉過程中肇東根、葉的LT50降低幅度明顯大于賽迪10,二者差異顯著(P<0.05)。肇東葉的LT50在處理前低于賽迪10,根高于賽迪10,在CH1和CH2處理后均低于賽迪10。

表1 不同苜蓿品種半致死低溫比較Table 1 Comparison of semi-lethal low temperature between two alfalfa varieties (℃)
2.2 抗寒鍛煉過程中不同紫花苜蓿品種根、葉中可溶性糖含量變化
NG 組肇東苜蓿從再生第7 天到第21 天根中可溶性糖含量逐步降低,賽迪10 在第14 天之前降低而后升高(圖1)。經CH1處理后,2 個苜蓿品種根中可溶性糖含量均升高,賽迪10 增幅較大(P<0.05)。經CH2處理后,肇東根中可溶性糖含量繼續增加,而賽迪10 降低,最終肇東高于賽迪10。NG 組苜蓿葉中可溶性糖含量隨再生變化較小,CH 組2 個苜蓿品種葉中可溶性糖含量均高于NG 組。CH1處理后肇東和賽迪10 葉中可溶性糖含量明顯高于CK,賽迪10 葉中可溶性糖含量顯著高于肇東(P<0.05)。CH2處理后肇東與賽迪10 葉中可溶性糖含量降低,賽迪10 的含量低于肇東。
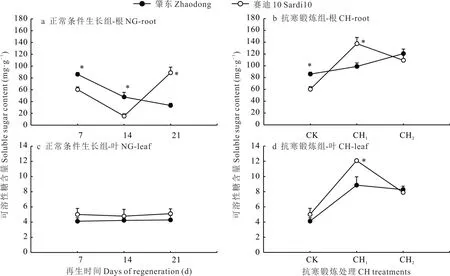
圖1 抗寒鍛煉和正常生長條件下苜蓿根和葉中可溶性糖含量變化Fig.1 Changes of soluble sugar content of alfalfa roots and leaves under cold acclimation and normal growth conditions
2.3 抗寒鍛煉過程中不同紫花苜蓿品種根、葉中淀粉含量變化
NG 組2 個苜蓿品種從再生第7 天到第21 天,根中淀粉含量均大幅度降低(圖2)。CH1處理后2 個品種根中淀粉含量基本保持不變,但經CH2處理后肇東根中淀粉含量降低,賽迪10 增加且顯著高于肇東(P<0.05)。NG組肇東葉中淀粉含量隨再生表現為先降低后增加的趨勢,賽迪10 則逐步降低。抗寒鍛煉過程中,2 個苜蓿品種葉中淀粉含量均先降低后增加,肇東的變化幅度較小,且在CK、CH1和CH2處理下均顯著高于賽迪10(P<0.05)。
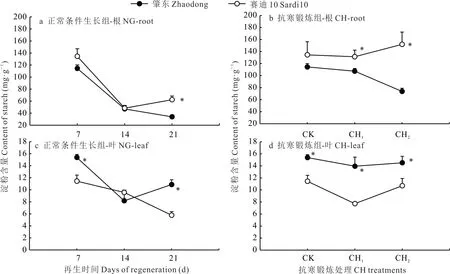
圖2 抗寒鍛煉和正常生長條件下苜蓿根和葉中淀粉含量變化Fig.2 Changes of starch content of alfalfa roots and leaves under cold acclimation and normal growth conditions
2.4 非結構性碳水化合物含量與紫花苜蓿抗寒性的相關性分析
根可溶性糖與根LT50、葉LT50、根淀粉、葉可溶性糖的相關性達到極顯著水平(表2,P<0.01),葉可溶性糖與根LT50、葉LT50、根可溶性糖、根淀粉的相關性達到顯著水平(P<0.05)。根淀粉與根LT50、根可溶性糖、葉可溶性糖的相關性達到極顯著水平(P<0.01),葉淀粉含量與葉LT50極顯著相關(P<0.01),與其他指標相關性較低。苜蓿根、葉的LT50與各指標為負相關,根、葉可溶性糖和淀粉之間的相關性為正相關。

表2 不同指標間相關性分析Table 2 Correlation analysis among different indexes
3 討論
3.1 紫花苜蓿可溶性糖和淀粉含量及其與抗寒性的關系
有研究表明糖類的代謝變化與植物抗寒性直接相關,植物在低溫脅迫條件下積累可溶性糖可以增加細胞的滲透調節能力,預防凍害的發生[4,6,17];越冬期間的植物和抗寒性強的植物可溶性糖含量較高[7]。Castonguay 等[7]在3 個不同抗寒性苜蓿品種(Rambler,Apica,CUF-101)上的研究表明苜蓿根中可溶性糖含量較高時,LT50較低。本試驗結果亦是如此,接受抗寒鍛煉后肇東和賽迪10 根、葉的LT50隨著其中可溶性糖含量的增加而降低(相關系數r值分別為-0.728、-0.587 和-0.601,-0.387,P<0.05)。以上研究結果證實可溶性糖含量越高,苜蓿抗寒性越強。淀粉是植物抗寒鍛煉過程中非結構性碳水化合物的主要儲備形式[7],在植物抗寒性能的提高和安全越冬中起著重要作用。本研究2 個苜蓿品種根、葉淀粉含量及劉香萍等[8]苜蓿根冠淀粉含量,均與抗寒性間呈顯著負相關關系。Xu 等[6]研究表明,苜蓿抗寒鍛煉后根頸淀粉含量大幅度增加,在溫度繼續降低情況下,淀粉含量降低。本試驗中,經歷CH1降溫處理后苜蓿根淀粉含量變化較小,但仍明顯高于正常生長植株,保持在較高水平,但接受CH2低溫處理后抗寒性強的肇東根中淀粉含量下降31.39%,CH2處理后變化與Xu 等[6]用抗寒性較強的苜蓿品種得出的結果一致。在抗寒鍛煉前期二者淀粉含量變化產生差異可能主要是由于本試驗是在苜蓿刈割后再生期進行的抗寒鍛煉。從正常條件下生長植株根、葉中淀粉和可溶性糖的變化中可以看到,苜蓿再生初期需要大量消耗根、葉中可溶性糖和淀粉。本試驗中,與正常條件下生長植株根中可溶性糖、淀粉含量大幅度降低相比,抗寒鍛煉后植株可溶性糖和淀粉保持在較高水平,也是苜蓿再生過程對抗寒鍛煉的響應,使器官保持較高的抗寒性。
不同苜蓿品種抗寒性差異與其根中累積可溶性糖的能力有關。石立媛等[18]研究表明,抗寒鍛煉后(10 月15日)抗寒性強的苜蓿品種根中可溶性糖含量高于抗寒性弱的品種。同樣地,本研究CH1處理后抗寒性強的肇東根中可溶性糖含量仍低于抗寒性弱的賽迪10,但繼續進行CH2處理,使溫度降低到晝夜溫度為10 ℃/6 ℃后,肇東根、葉中可溶性糖含量高于賽迪10。Castonguay 等[19]研究指出苜蓿在溫度降低到10 ℃以下時,抗寒性極強的苜蓿品種根冠中淀粉含量低于不抗寒的品種,隨著溫度繼續降低,抗寒性極強的苜蓿根中的淀粉含量快速降低,而抗寒性弱的品種淀粉含量快速增加。抗寒鍛煉前后,肇東根中的淀粉含量低于賽迪10,在CH2處理后肇東的淀粉含量降低,賽迪10 含量增加,表現出的品種間差異非常明顯。CH1處理后,賽迪10 根、葉中可溶性糖含量大幅增加,且顯著高于肇東,葉中淀粉含量大幅降低,且顯著低于肇東。而抗寒性弱的苜蓿品種在降溫期間,地上生物量增加且顯著高于抗寒性強的品種[20],因此賽迪10 不斷消耗根中物質[20]與葉中的淀粉,使根、葉中可溶性糖含量保持在較高水平,并使器官保持較高再生能力,導致抗寒鍛煉后根的抗寒性與肇東差異較大。
3.2 紫花苜蓿根、葉中淀粉與可溶性糖之間的轉運與轉化
抗寒鍛煉改變了植物非結構性碳水化合物地上、地下的轉運,特別是淀粉,作為植物體內非結構性碳的重要貯存形式,在低溫脅迫下其可以水解成可溶性糖來提高器官的抗寒性[6-7,21-22]。正常條件生長植株在再生初期,根中淀粉、可溶性糖含量降低,淀粉轉化成可溶性糖,并供應給地上部生長,同時葉中淀粉含量也降低,可溶性糖含量保持在較低水平,消耗的這些非結構性碳水化合物主要用于地上部再生[11,20]。CH1處理后,苜蓿葉中淀粉含量降低,根、葉中可溶性糖含量增加,而根中淀粉含量變化較小,葉中淀粉向可溶性糖轉化,并向根中轉運;CH2處理后地上、地下的轉運較少。
本試驗中,抗寒鍛煉處理下苜蓿根、葉中淀粉含量與可溶性糖含量表現出互補的變化趨勢,發生淀粉與可溶性糖的相互轉化,且根、葉間及品種間表現出不同的轉化方向。除上述CH1處理后,葉中發生淀粉向可溶性糖轉化外,CH2處理后,2 個品種葉中可溶性糖含量降低,淀粉含量增加,葉中可溶性糖向淀粉轉化,而2 個品種根中非結構性碳水化合物的轉化方向不同,肇東根中淀粉含量降低,可溶性糖含量增加,淀粉向可溶性糖轉化,賽迪10根淀粉含量增加,可溶性糖含量降低,可溶性糖向淀粉轉化。抗寒性較強的苜蓿品種WL353LH(秋眠級4)的研究結果表明,苜蓿根莖中淀粉含量在抗寒鍛煉后期被利用[6],在苜蓿其他品種[7,21,23]的研究表明苜蓿在自然降溫條件下根中淀粉含量降低,使可溶性總糖含量增加,并認為低水平的淀粉含量有利于提高苜蓿抗寒力。姚勝蕊等[22]和Palonena 等[24]也在桃(Prunus persica)和紅樹莓(Rubus idaeus)的研究中指出越冬器官中淀粉利用率越高,抗寒性越強。本試驗中,2 個不同抗寒性苜蓿品種根中淀粉利用率差異較大,抗寒性強的肇東苜蓿在CH2處理后淀粉向可溶性糖轉化,利用率提高,抗寒性弱的賽迪10 根中可溶性糖向淀粉轉化,不僅不利用淀粉反而積累淀粉。本研究2 個苜蓿品種在低溫處理后根中非結構性碳水化合物轉化方向不同可能是因為二者抗寒性差異大所致,這也說明抗寒鍛煉后期苜蓿根中淀粉向可溶性糖的轉化是保障地下器官抗凍性的重要途徑,而轉化方向的不同也可能是品種間抗寒性差異大的重要原因。
4 結論
紫花苜蓿在抗寒鍛煉過程中根淀粉含量及根、葉中可溶性糖含量與其抗寒性高度相關,在抗寒鍛煉初期葉中淀粉轉化成可溶性糖并向根部轉運,提高根、葉的抗寒性,在抗寒鍛煉持續推進后,根中發生淀粉向可溶性糖轉化,從而使根的抗寒性得到不斷提高。抗寒性強弱不同的苜蓿品種在抗寒鍛煉中對于非結構性碳水化合物的利用與轉化模式也不同。抗寒鍛煉初期,與抗寒性強的肇東相比,抗寒性弱的賽迪10 根、葉中消耗更多的非結構性碳水化合物保障和用于再生。抗寒鍛煉后期肇東根中淀粉向可溶性糖轉化,賽迪10 根中可溶性糖向淀粉轉化。因此,苜蓿在抗寒鍛煉初期消耗較少的非結構性碳水化合物,后期根中淀粉向可溶性糖轉化,有利于品種抗寒性增加。
