鉛鹵鈣鈦礦泄漏對垂穗披堿草幼苗生長的影響
2023-12-08 15:30:40李庭倫李一亨余慧江再莉唐立濤王長庭胡雷
草業學報 2023年12期
李庭倫,李一亨,余慧,江再莉,唐立濤,王長庭,胡雷*
(1. 西南民族大學青藏高原研究院,四川 成都 610225;2. 小金縣林業和草原局,四川 阿壩藏族羌族自治州 624099)
鉛鹵鈣鈦礦作為鈣鈦礦電池中使用最多的材料,是由有機陽離子與鹵化鉛八面體形成的三維結構材料,其光電轉化效率已達25%,超過了目前商業主流的光伏器件(多晶硅)的光電效率,并有望完全替代多晶硅,在新能源開發中具有巨大的發展潛力和應用前景[1-2],但其本身具有離子性質,它的重金屬離子對環境變化十分敏感[3],長時間紫外線照射、氧氣和溫度的升高等都會顯著影響鈣鈦礦材料中重金屬的泄漏[4-5],而由此導致的生態效應知之甚少。一旦鈣鈦礦材料導致重金屬泄漏至土壤中,這些重金屬會在生物體內不斷累積,并轉變為毒性甲基化合物,這種化合物將會隨著食物鏈而最終進入人體,嚴重影響人體健康。目前有關鈣鈦礦材料導致重金屬泄漏的生態影響效應研究,其關注對象多為人體細胞[6]、土壤微生物或大型動物。Zhai 等[7]研究了鉛基鈣鈦礦納米粒子對土壤細菌群落的影響,發現Pb2+的存在通過為鈣鈦礦納米粒子與土壤細菌相互作用提供陽離子橋,從而增加了鈣鈦礦納米粒子對土壤細菌群落的毒性效應。Danae 等[8]評估了5 種鈣鈦礦型納米粒子[碘化鉛(PbI2)和溴化鉛(PbBr2)基材料]對斑馬魚幼蟲和成魚的生物毒性,鈣鈦礦材料中的可溶性Pb 對斑馬魚死亡率和基因表達方面產生顯著影響。然而,目前對食物鏈的第一營養級—植物影響的研究則十分缺乏,且主要集中在經濟作物上,如辣椒(Capsicum annuum)、卷心菜(Brassica campestris)和薄荷(Mentha spicata)等[9]。
Pb 作為最嚴重的重金屬污染元素之一,不易分解,對生物體能夠產生顯著毒性且持久的物質[10]。環境中的Pb 主要通過葉片的吸附和根系的吸收兩個途徑進入植物體內[11],一旦進入植物體內,輕度Pb 脅迫影響植物生長,重度則導致其死亡[12]。然而,不同植物Pb 耐受能力存在顯著差異。對于自然界中的超富集植物而言,如牧草剪股穎(Agrostis matsumurae)和羊茅草(Festuca ovina),其對Pb 的耐受能力分別達到13488 和11750 mg·kg-1[13-14],而鹽生草(Halogeton glomeratus)對Pb 的耐受范圍則為207~621 mg·kg-1[15]。不同的Pb 耐受能力決定了其對Pb 脅迫的響應程度。但也有研究表明,低濃度Pb 脅迫可能對植物生長具有積極的刺激作用。如,低濃度Pb 處理下印度芥菜(Brassica jouncea)根系生長更好[16],但高濃度Pb 則會導致根系體積和生物量的降低[17]。因此,明確植物的Pb 耐受范圍,是科學研究Pb 脅迫下植物生長響應的關鍵。
盡管不同植物Pb 耐受能力不同,但只要Pb2+被植物吸收,其在植物體內富集能力和遷移能力就會變強,將會對植物生長產生更為嚴重的危害。研究指出,向土壤中添加Pb 后,土壤中有效Pb 濃度僅提高了10%,而Pb 富集能力較強的植物,如薄荷組織內的鉛濃度則提高100%,高濃度的鉛污染(250 mg·kg-1)將導致大多數薄荷由于鉛中毒而變黑腐爛[9]。而對Pb 富集能力較弱的植物來說,輕度Pb 污染都將對其產生嚴重的影響。如辣椒和卷心菜,一旦受到Pb 污染,兩種植物富集在體內的Pb 的遷移性將增加10 倍以上[9]。因此,研究植物生長對Pb 脅迫的響應不僅需要關注其Pb 耐受能力,更需要關注其Pb 富集能力[18-20]。
鉛鹵鈣鈦礦材料在太陽能資源豐富的青藏高原具有廣闊的應用前景,披堿草屬植物是青藏高原高寒草甸植物群落優勢物種,植株生長茂盛,粗蛋白含量高、適口性好,是良好的牧草品種[21-22]。然而,目前鮮有研究關注鉛鹵鈣鈦礦材料泄漏對披堿草植物生長發育的影響,尤其是對其耐受能力和富集能力影響的研究極少。這極大限制了研究者對鉛鹵鈣鈦礦材料廣泛應用背景下披堿草屬植物富集能力和耐受范圍的認知。因此,本研究通過盆栽方法,模擬不同鉛鹵鈣鈦礦材料泄漏導致Pb 脅迫梯度下,垂穗披堿草(Elymus nutans)幼苗生長、Pb 富集能力和耐受范圍,充分了解鈣鈦礦泄漏引起的生態效應,為高寒草甸土壤鉛污染治理和草場健康管理提供數據和理論支撐。
1 材料與方法
1.1 試驗樣地
試驗樣地位于四川省西南民族大學青藏高原研究院溫室大棚內(102°57′53″ E,30°33′45″ N),該地區是亞熱帶濕潤季風氣候,年平均氣溫16 ℃,年降水量1000 mm 左右,海拔499 m。試驗于2022 年5-7 月進行。
1.2 試驗材料
植物材料是“康北”垂穗披堿草,由西南民族大學青藏高原研究院提供。供試土壤為無農藥殘留的田園土(保持土壤性質基本一致,該土壤不含種子、土壤動物、侵入體和新生體,可以保證種植一致性)。
盡管鉛鹵鈣鈦礦泄漏的影響主要是Pb 脅迫引起的生態效應,但鉛鹵鈣鈦礦材料中主要是碘化鉛(PbI2),為了更為真實模擬鉛鹵鈣鈦礦泄漏引起的生態效應,本研究選擇PbI2。該材料購買自麥克林(https://www.macklin.cn/)。
1.3 試驗方法
1.3.1 試驗設計 采用盆栽試驗,設置1 個不添加碘化鉛的處理為對照組,5 個不同鉛離子濃度處理,共計6個處理,每個處理3 個重復。
鉛鹵鈣鈦礦太陽能電池中的甲基胺碘化鉛(CH3NH3PbI3)層厚度約為600 nm,CH3NH3PbI3中的Pb 含量為33.4%(質量比)。由于中國土壤平均Pb 含量在36 mg·kg-1左右,所以第1 組鈣鈦礦泄漏梯度為36 mg·kg-1Pb,第2 組增加到72 mg·kg-1Pb,第3 組增加到130 mg·kg-1Pb。我國《土壤環境質量農用地土壤污染風險管控標準(試行)》[23]規定農業用地鉛含量上限為170 mg·kg-1Pb,因此第4 組處理鉛含量升高至170 mg·kg-1Pb。為了充分確定鈣鈦礦鉛泄漏導致的生態效應,本研究增設了一個最高鈣鈦礦泄漏梯度260 mg·kg-1Pb。
1.3.2 種植方法 播種前用蒸餾水浸泡種子10 min。土壤過8 mm 篩,揀去石塊和根系,混合均勻。取一部分進行土壤理化性質的測定。
稱取一定量混合均勻的土壤,倒入花盆(2 L)中,再按照鉛離子濃度梯度稱取碘化鉛固體粉末倒入花盆,與土壤混合均勻。取大小相對一致的種子50 顆,均勻地撒播在土壤表面,再輕輕將其覆蓋。
播種開始于2022 年5 月31 日,對照組、第1 組(36 mg·kg-1Pb)、第2 組(72 mg·kg-1Pb)、第3 組(130 mg·kg-1Pb)和第4 組(170 mg·kg-1Pb)分別于6 月3 日、6 月4 日、6 月3 日、6 月4 日和6 月3 日出苗,最大組(260 mg·kg-1Pb)于6 月3 日出苗,但迅速死亡。之后15 d 記錄出苗數量,連續3 d 出苗數量均無變化后,于6 月17 日開始每2 d 測量幼苗高度。
1.4 測定指標
1.4.1 植物數據采集和指標計算 從播種(記為試驗開始第1 天,T1)直到出苗數達到最大值N,即試驗結束,當天記為Tn,每日記錄每盆種子出苗數(Ni),當天記為Ti。其中,第一次出苗的前一天記為Tj。由于在260 mg·kg-1Pb 濃度下,所有幼苗均已死亡,因此以下分析均不包含該梯度的數據。
在出苗后期Tn,測量每株幼苗高度(cm)。之后拔出所有幼苗,小心掃去附著的泥土,測量鮮重;然后108 ℃殺青后,60 ℃烘干48 h 至恒重,測量其干重。
1.4.2 土壤理化性質的測定 將每個花盆內土壤取樣,采用電位法[24]測定土壤pH,采用氯化鉀浸提法[25]測定土壤銨態氮和硝態氮含量,使用碳氮分析儀(TOC-L CPH, 日本)測定土壤全碳含量。
1.4.3 鉛含量測定 植物收獲后洗凈沙土,105 ℃殺青30 min,80 ℃烘干8 h 后磨碎。用四酸電熱板消解土壤,用硝酸-高氯酸消解植物樣品。土壤和植物樣品消解后用電感耦合等離子體質譜儀(ICAP 6300, 美國)測定Pb 含量。植物Pb 富集指數=植物鉛含量/土壤鉛含量[26-27]。
1.4.4 植物鉛耐受能力 計算不同鉛脅迫梯度下垂穗披堿草幼苗高度(HPb)和生物量(BPb)分別與對照組幼苗高度(HCK)和生物量(BCK)的比值,即為高度耐受指數(height tolerance index, HTI)和生物量耐受指數(biomass tolerance index, BTI)。耐受指數反映了植物對重金屬脅迫的耐受程度,當該指數大于1 時,表示該脅迫梯度對植物生物量具有促進作用,當耐受指數等于1 時,該脅迫梯度為測試植物的耐性下限;當高于某個脅迫梯度測試植物無法存活時,即耐受指數為0,此脅迫梯度為測試植物的耐性上限[28-29]。
1.5 統計分析
使用Excel 2019 進行作圖,SPSS 19.0 進行單因素方差分析和Duncan 顯著性檢驗。數據使用“平均值±標準差”表示。采用Pearson 分析土壤理化性質和垂穗披堿草幼苗性狀之間的相關性,使用R 3.6.1 的corrplot 包作相關性熱圖。為了進一步確定鈣鈦礦泄漏對垂穗披堿草幼苗的影響,本研究使用結構方程模型(structural equation model, SEM)分析不同Pb 脅迫梯度如何通過影響幼苗生長進而改變其Pb 富集和耐受能力。
2 結果與分析
2.1 垂穗披堿草幼苗生長情況
不同鉛濃度沒有顯著影響垂穗披堿草的出苗率(P=0.426),但36 和72 mg·kg-1鉛濃度處理下,出苗率表現出增加的趨勢。垂穗披堿草第一次出苗時間,即出苗時滯受到鉛濃度的顯著抑制(P=0.024),尤其是170 mg·kg-1鉛濃度處理使垂穗披堿草第一次出苗時間比對照平均晚了1 d。垂穗披堿草出苗速率(P=0.050)表現為隨鉛濃度增加而顯著降低的趨勢,尤其是在170 mg·kg-1鉛濃度處理下,其出苗速率下降了50.47%,但低濃度鉛處理對出苗速率無顯著影響。垂穗披堿草出苗持續時間(P=0.002)也隨鉛濃度的增加顯著縮短,在170 mg·kg-1鉛濃度處理下,其出苗持續時間平均縮短了4.4 d(表1)。
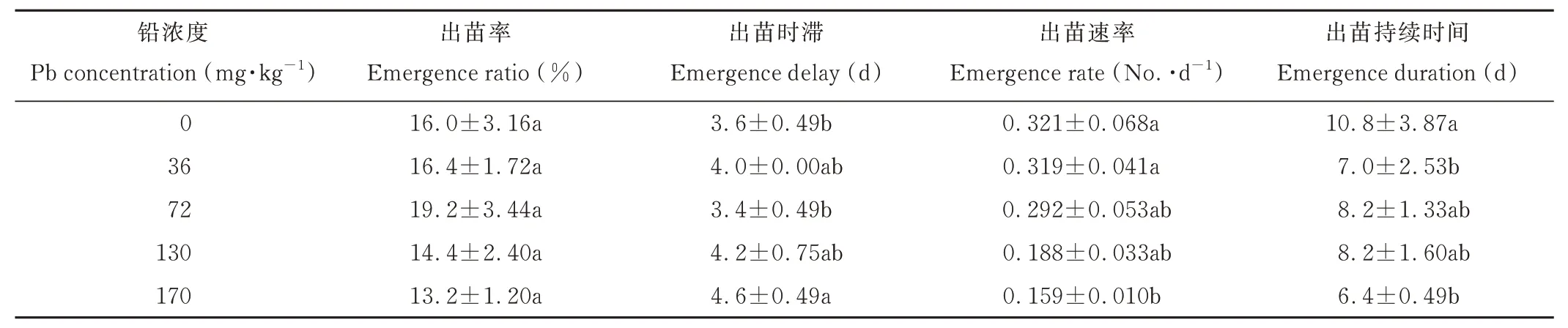
表1 不同鉛濃度對垂穗披堿草出苗情況的影響Table 1 The effects of Pb gradients on the seedlings emergence of E. nutans
2.2 垂穗披堿草幼苗高度和生物量及耐受指數
垂穗披堿草幼苗平均高度對鉛濃度具有極顯著響應(P<0.001)(圖1),對照組(14.26 cm)顯著高于鉛處理組。但不同鉛處理之間無顯著差異,隨鉛濃度增加而降低。36 和72 mg·kg-1濃度下單株生物量與對照差異不顯著,但130 和170 mg·kg-1濃度下則分別顯著降低37.2%和64.2%。
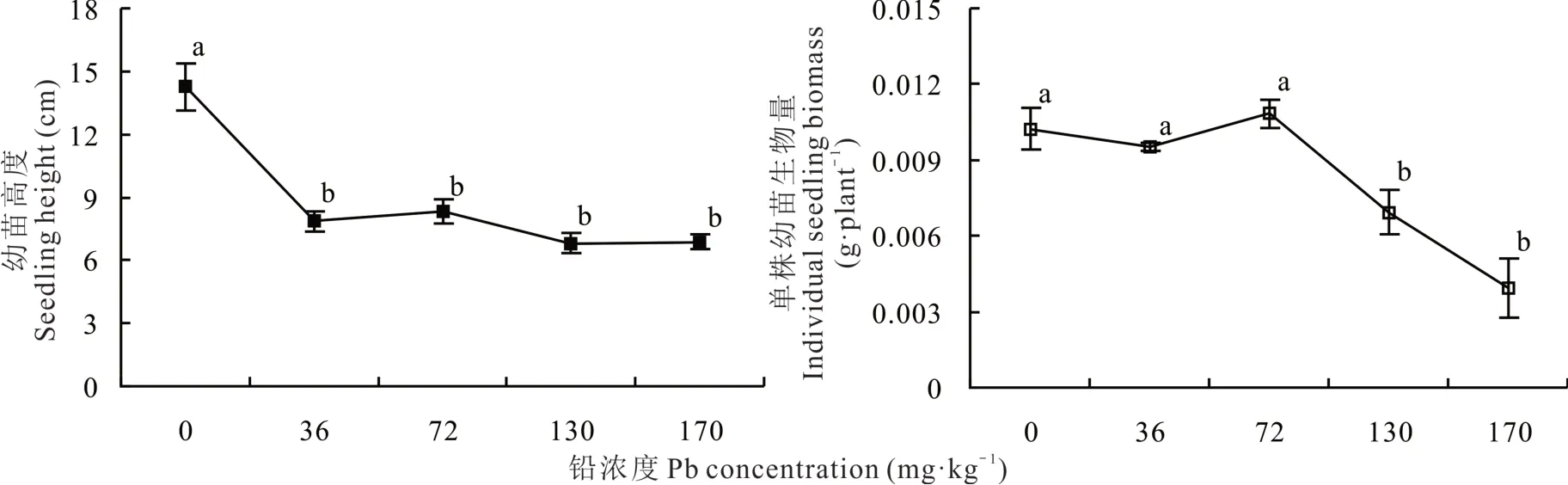
圖1 不同鉛濃度梯度處理下垂穗披堿草幼苗高度和生物量的變化Fig.1 Changes in height and biomass of E. nutans seedling with Pb concentration gradients
垂穗披堿草高度耐受指數(HTI)在所有鉛濃度處理下均小于1。生物量耐受指數(BTI)在鉛濃度為36 mg·kg-1時小于1,在72 mg·kg-1時與1 接近且大于1,當超過72 mg·kg-1時,顯著小于1,說明垂穗披堿草幼苗對鉛的耐性下限位于36~72 mg·kg-1范圍內。由于垂穗披堿草幼苗在170 mg·kg-1鉛脅迫下可以存活,但在本研究最大梯度260 mg·kg-1鉛脅迫下種子無法萌發,因此其耐性上限位于170~260 mg·kg-1范圍內(圖2)。為了進一步確定BTI 耐性下限和上限,本研究將BTI 與鉛濃度(Pb)進行二項式回歸,結果顯示BTI=0.970+0.003Pb-3.744×10-5Pb2(r2=0.827,P<0.001),因此,耐性下限為68.42 mg·kg-1,上限為205.94 mg·kg-1。
2.3 不同鉛濃度處理下植物鉛富集指數
隨著鉛濃度的增加,垂穗披堿草幼苗鉛含量逐漸增加,在低鉛濃度處理間(36~72 mg·kg-1)具有增加趨勢,但不顯著;當鉛濃度超過72 mg·kg-1時,幼苗鉛含量顯著增加(圖3)。鉛脅迫顯著抑制了垂穗披堿草的鉛富集能力,但抑制程度與鉛濃度無關。

圖3 不同鉛濃度梯度處理下植物鉛含量和富集指數Fig. 3 The Pb concentrations of plant and the Pb accumulation index with Pb concentration gradients
2.4 不同鉛濃度處理下土壤理化性質
不同鉛濃度處理下土壤pH 呈降低趨勢,但不顯著(P=0.114),僅在最高鉛濃度處理下(170 mg·kg-1)顯著低于對照;土壤銨態氮(NH4+-N)含量在不同鉛濃度處理下差異不顯著(P=0.610),而土壤硝態氮(NO3--N)含量則隨鉛濃度增加呈增加趨勢(P=0.204),僅在最高鉛濃度處理下(170 mg·kg-1)顯著高于對照;土壤總碳含量在72 mg·kg-1鉛濃度處理下最低,在170 mg·kg-1鉛濃度處理下最高,但與對照均無顯著差異(P=0.185)(表2)。
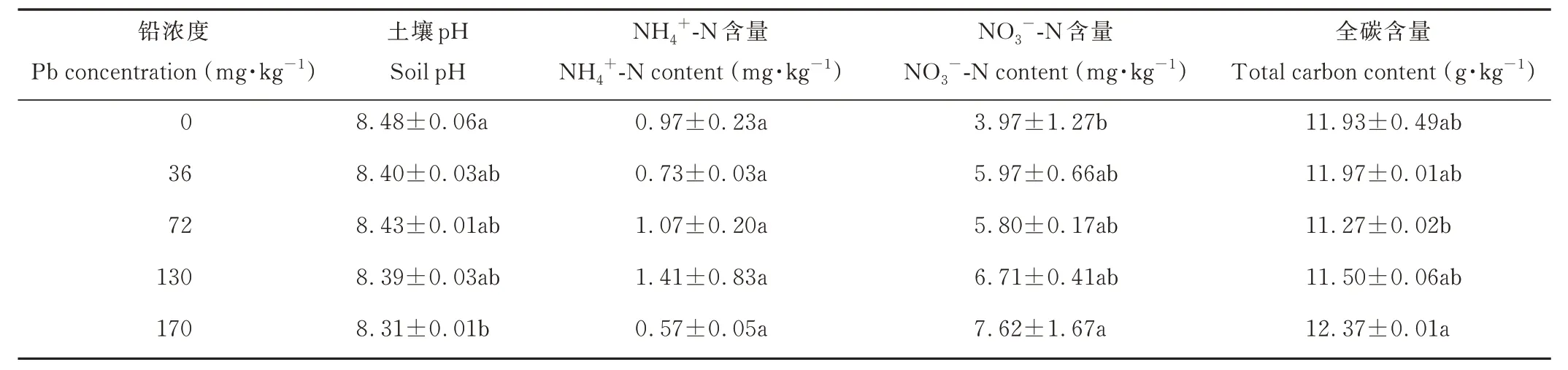
表2 不同鉛濃度梯度土壤理化性質Table 2 Soil physicochemical properties with different Pb concentration gradients
2.5 不同鉛濃度處理下土壤理化性質和垂穗披堿草幼苗性狀的相關性分析
相關性分析表明,垂穗披堿草幼苗相關性狀與土壤pH 和硝態氮(NO3--N)顯著相關(圖4),尤其是土壤pH與出苗率(emergence ratio)、出苗速率(emergence rate)及生物量耐受指數(BTI)顯著正相關,土壤硝態氮則分別與出苗率和出苗速率顯著負相關。土壤銨態氮(NH4+-N)和全碳(TC)與幼苗指標無顯著相關性。

圖4 土壤理化性質和垂穗披堿草幼苗性狀的Pearson 相關性分析Fig.4 The Pearson correlation analysis among soil physicochemical properties and seedling characteristics of E. nutans
2.6 鉛脅迫梯度下垂穗披堿草幼苗的結構方程模型分析
由土壤理化性質和垂穗披堿草幼苗的相關性分析可知(圖4),土壤pH 和硝態氮是主要因子,因此,將兩個因子納入結構方程模型(SEM),探討鉛脅迫對生長情況(seedling growth)的影響。出苗率受鉛脅迫影響不顯著(表1),且出苗速率具有高度共線性(圖4),因此,在SEM 分析中選擇出苗速率、出苗時滯和出苗持續時間共同表示生長情況(圖5)。此外,本研究將生物量耐受指數和高度耐受指數合并指示Pb 耐受能力(圖5),納入SEM 中進行分析。
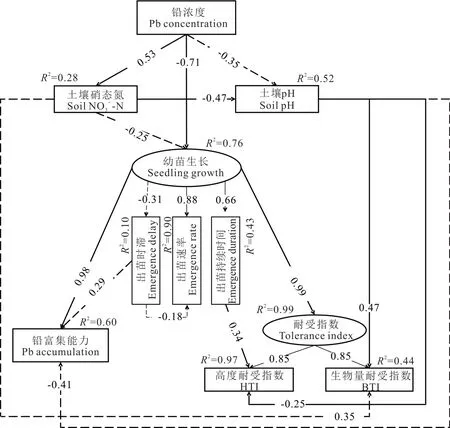
圖5 鉛濃度對垂穗披堿草幼苗生長情況的SEM 分析Fig.5 The SEM analysis for the seedling growth of E. nutans affected by Pb concentration gradients
SEM 分析結果表明,鉛脅迫直接顯著抑制垂穗披堿草幼苗生長情況(P<0.001),尤其是出苗速率和出苗持續時間,進而影響幼苗Pb 富集能力和Pb 耐受能力。鉛脅迫也通過顯著增加土壤硝態氮含量(P=0.020),導致土壤pH 顯著降低(P=0.032),進而影響生物量耐受指數(P=0.004)和高度耐受指數(P=0.017)。生長情況則直接影響植物鉛富集能力(P<0.001)和鉛耐受能力(P<0.001),其中出苗持續時間顯著影響高度耐受指數(P=0.001)(圖5)。
綜合來看,鉛脅迫直接或通過土壤理化性質間接影響了垂穗披堿草幼苗生長情況、鉛富集能力和鉛耐受能力(表3)。其中,鉛脅迫對幼苗生長情況的抑制作用最大(0.847),其次為土壤硝態氮(0.255)。對幼苗高度和生物量耐受能力影響最大的均為幼苗生長情況;其次為土壤pH。但土壤pH 主要提高生物量耐受能力,降低高度耐受能力。對幼苗鉛富集能力具有最大積極影響的仍然為幼苗生長情況,達到0.889,具有最大抑制作用的為鉛脅迫,達到了0.510。
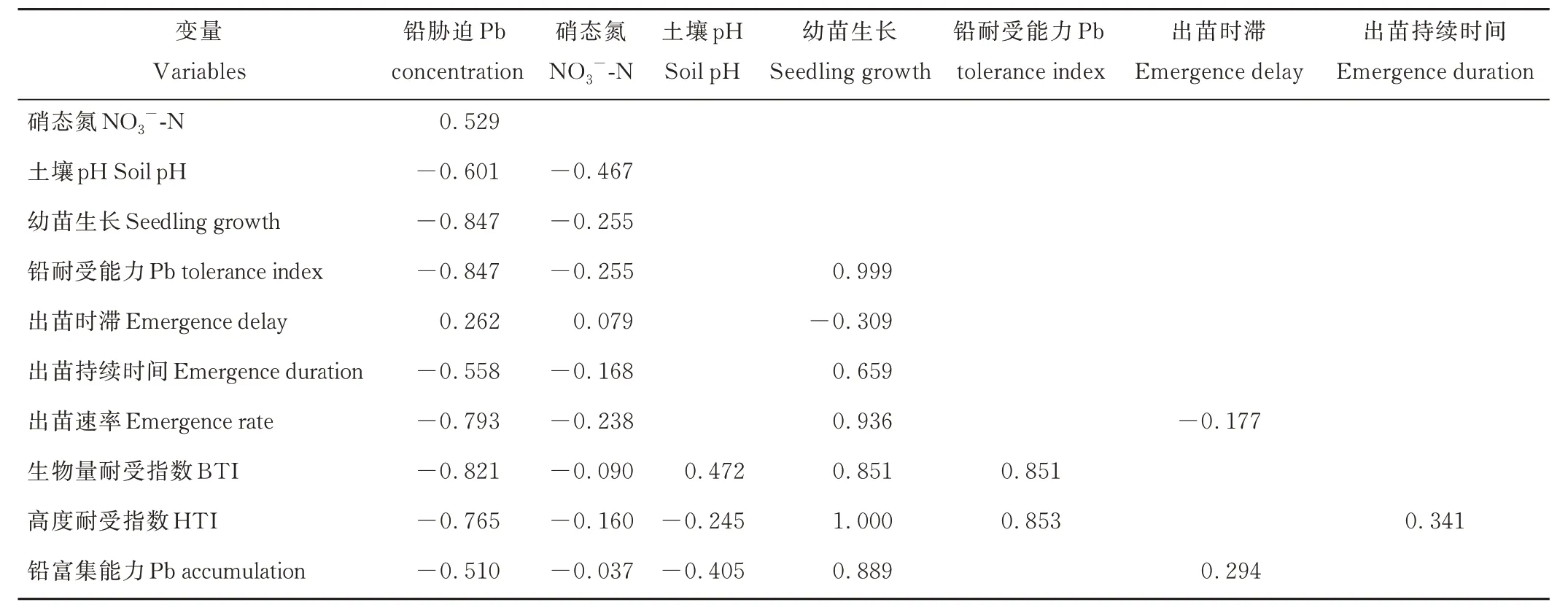
表3 不同變量的綜合影響系數Table 3 The standardized total effects of the variables
3 討論
3.1 鉛脅迫對垂穗披堿草出苗生長情況的影響
重金屬脅迫下,植物種子萌發和出苗情況能夠反映該植物種子活力和芽苗生長能力[30]。植物種子萌發和出苗情況對重金屬的脅迫通常表現為濃度效應,即低濃度促進而高濃度抑制[31]。不同植物種子對同一重金屬濃度的響應不同[19],鉛濃度小于200 mg·L-1時可促進草地早熟禾(Poa pratensis)種子萌發,大于300 mg·L-1則表現為顯著抑制作用[32];而白羊草(Bothriochloa ischaemum)種子對低濃度的鉛脅迫(<100 mg·L-1)反應不敏感,在大于300 mg·L-1鉛脅迫時則表現為促進作用[33]。因此,植物在重金屬脅迫下存在一個積極響應和逐漸受損的過程[34]。本研究中垂穗披堿草出苗情況同樣表現為濃度效應。不同的是,垂穗披堿草出苗率對鉛濃度的響應較弱,僅在36 和72 mg·kg-1鉛處理下表現為增加趨勢,且出苗時滯少于對照處理;而隨著鉛濃度的增加,垂穗披堿草出苗速率逐漸降低,出苗持續時間逐漸縮短。這表明72 mg·kg-1鉛濃度可能是影響垂穗披堿草出苗情況的關鍵濃度,該濃度處理下,一些與種子萌發相關的酶活性可能被促進[31],從而導致了其具有較高的出苗率和更快的出苗時間。
3.2 鉛脅迫對垂穗披堿草耐受能力的影響
垂穗披堿草幼苗高度和生物量對鉛脅迫梯度響應不同,單株幼苗生物量對小于72 mg·kg-1的鉛脅迫響應不顯著,對大于該鉛濃度的處理表現為顯著降低。這可能與植物的濃度效應有關[31],超出一定重金屬脅迫濃度,植物體內重金屬敏感基因表達將會受到顯著抑制,導致植物營養元素吸收平衡紊亂,阻礙植物生物量的積累[35]。垂穗披堿草幼苗高度在鉛脅迫時顯著降低,與李慧芳等[36]的研究結果一致。但本研究同時表明,在不同鉛濃度處理之間垂穗披堿草幼苗高度不存在顯著差異,沒有表現出濃度效應。可見,垂穗披堿草幼苗高度和生物量對鉛脅迫的顯著響應,可能與垂穗披堿草自身出苗情況有關[37-38]。出苗情況是影響垂穗披堿草高度和生物量耐受能力最重要的因子,如出苗持續時間一定程度上促進了植物的高度耐受能力。
通過對高度耐受指數和生物量耐受指數研究發現,本研究中測試的垂穗披堿草Pb 耐受范圍為68.42~205.94 mg·kg-1,這遠小于其他研究中的耐受范圍。如劉秀梅等[28]研究認為披堿草(Elymus dahuricus)Pb 耐受范圍為332.32~1315.94 mg·kg-1,徐雅梅等[39]研究認為野生垂穗披堿草的鉛耐受上限達到800 mg·kg-1。這可能是由于植物物種差異[40-41]、Pb 脅迫處理時間不同[42]或評價指標不同[22]導致的。劉秀梅等[28]使用的披堿草與本研究中的不同,且Pb 脅迫處理時間為幼苗長出第一片真葉后的第20 天。盡管本研究和徐雅梅等[39]使用的為同一物種,但本研究使用材料為西南民族大學垂穗披堿草“康北”品種,徐雅梅等[39]使用的為野生垂穗披堿草,且耐性指標主要為葉綠素含量、細胞質膜透性等。
3.3 鉛脅迫對垂穗披堿草幼苗鉛含量和富集指數的影響
面對單一重金屬脅迫時,植物體內重金屬含量隨脅迫程度增加而增加,但重金屬富集能力逐漸降低[35-36]。本研究中垂穗披堿草鉛含量隨鉛濃度增加而顯著增加,但鉛含量始終低于鉛脅迫水平。這是由于本研究中垂穗披堿草只在四葉期進行取樣,因此鉛含量較低。隨著植物生長期的延長,鉛富集能力將逐漸增加到一個最大值,這個時期通常是植物從營養生長向生殖生長的過渡期,此時植物鉛含量和富集能力開始下降[35]。在一項關于同屬披堿草對鉛脅迫響應的研究中顯示,成體時的該披堿草鉛含量顯著高于鉛脅迫濃度[43]。本研究中垂穗披堿草鉛富集能力在受到鉛脅迫時顯著低于對照,且在72 mg·kg-1鉛濃度處理下最低。植物對Pb 的富集能力不僅與自身有關,而且隨土壤理化性質的改變而發生變化[44],如土壤Pb 含量[26]、土壤pH 值[45]等。本研究發現,鉛脅迫梯度對幼苗鉛富集能力具有最大的直接抑制作用,其次為土壤pH。
4 結論
Pb 脅迫并未改變垂穗披堿草的出苗率,但顯著抑制其出苗速率、出苗持續時間、幼苗高度、富集系數和耐受系數等(P<0.05)。單株幼苗生物量則表現為濃度效應,鉛濃度小于72 mg·kg-1無顯著作用,大于72 mg·kg-1時表現為抑制作用。垂穗披堿草幼苗對Pb 脅迫的耐受范圍為68.42~205.94 mg·kg-1。
Pb 脅迫主要通過兩個途徑影響垂穗披堿草幼苗的富集能力和耐受能力:1)直接影響幼苗生長情況,進而抑制其富集能力和耐受能力,尤其是出苗持續時間對高度的影響;2)直接增加土壤硝態氮含量,降低pH 值,顯著抑制幼苗高度和增加幼苗生物量(P<0.05)。
綜上所述,在一定Pb 脅迫梯度下,可以通過提高土壤pH 值,增加垂穗披堿草生物量,同時降低其生長高度和Pb 富集能力。
猜你喜歡
發明與創新(2022年30期)2022-10-03 08:40:56
動漫星空(興趣百科)(2020年12期)2020-12-12 05:31:40
人大建設(2018年6期)2018-08-16 07:23:10
文理導航·科普童話(2017年5期)2018-02-10 19:42:14
無人機(2017年10期)2017-07-06 03:04:36
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
少兒科學周刊·兒童版(2015年10期)2015-11-07 03:42:03
