不同辣椒栽培種的雜交親和性比較及胚胎拯救體系建立
2023-11-27 10:31:38施二榮胡能兵隋益虎袁友志來喜月
中國瓜菜 2023年11期
施二榮,胡能兵,隋益虎,袁友志,來喜月,王 勃
(1.安徽科技學(xué)院農(nóng)學(xué)院 安徽鳳陽 233100;2.安徽金泰種業(yè)有限公司 合肥 230031)
辣椒(Capsicum)為茄科辣椒屬一年生草本植物[1]。目前,全球共有5 個(gè)辣椒栽培種(Capsicum annuumL.、Capsicum chinenseL.、Capsicum frutescensL.、CapsicumpubescensL.、CapsicumbaccatumL.)和20 多個(gè)野生種[2],其中我國種植面積最大的是C.annuum,該栽培種具有產(chǎn)量高、適應(yīng)性強(qiáng)等特點(diǎn),但抗性差異較大;C.frutescens和C.chinense兼具辣度高、抗性強(qiáng)等特點(diǎn),但其栽培條件受區(qū)域限制較大且產(chǎn)量很低。有研究表明,各辣椒栽培種種間雜交均存在不同程度的生殖障礙[3-4]。
種間雜交是拓寬辣椒栽培種遺傳基礎(chǔ)和創(chuàng)制種質(zhì)資源的有效方式[5-6]。為了能有效創(chuàng)制新的辣椒種質(zhì)資源,聚合更多的有益性狀,前人結(jié)合胚胎拯救技術(shù)導(dǎo)入外源基因,拓寬主要栽培種的遺傳背景。Sui 等[7]通過C.annuum7033×C.chinense7020獲得F1雜種,并自交獲得大量分離后代;Yoon 等[8]將炭疽病抗性從C.baccatum滲入到C.annuum,在C.annuum回交后代中篩選出了大量高抗炭疽病的改良植株。
近年來高辣、高抗已成為辣椒育種的重要方向之一[9]。辣度較高、抗逆性較強(qiáng)的C.chinense、C.frutescens由于產(chǎn)量低而在我國栽培面積始終較小。為了把高辣度、強(qiáng)抗逆和高產(chǎn)量相結(jié)合,筆者利用3 個(gè)栽培種辣椒進(jìn)行遠(yuǎn)緣雜交,通過對(duì)雜交坐果率、胚萌發(fā)率、拯救時(shí)期以及F1幼胚生根效果等指標(biāo)的觀測(cè),旨在建立一套系統(tǒng)的胚胎拯救技術(shù)體系,提高遠(yuǎn)緣雜交的成功率。
1 材料與方法
1.1 材料
供試材料包含3 個(gè)辣椒栽培種(圖1),即8 份C.annuumL.、8 份C.chinenseL.和1 份C.frutescensL.共17 份試驗(yàn)材料(表1),均來自于安徽科技學(xué)院辣椒種質(zhì)資源庫,于2022 年3 月種植于安徽科技學(xué)院種植科技園3 號(hào)溫室,四周覆蓋30 目防蟲網(wǎng),栽培期間進(jìn)行正常的生產(chǎn)管理。
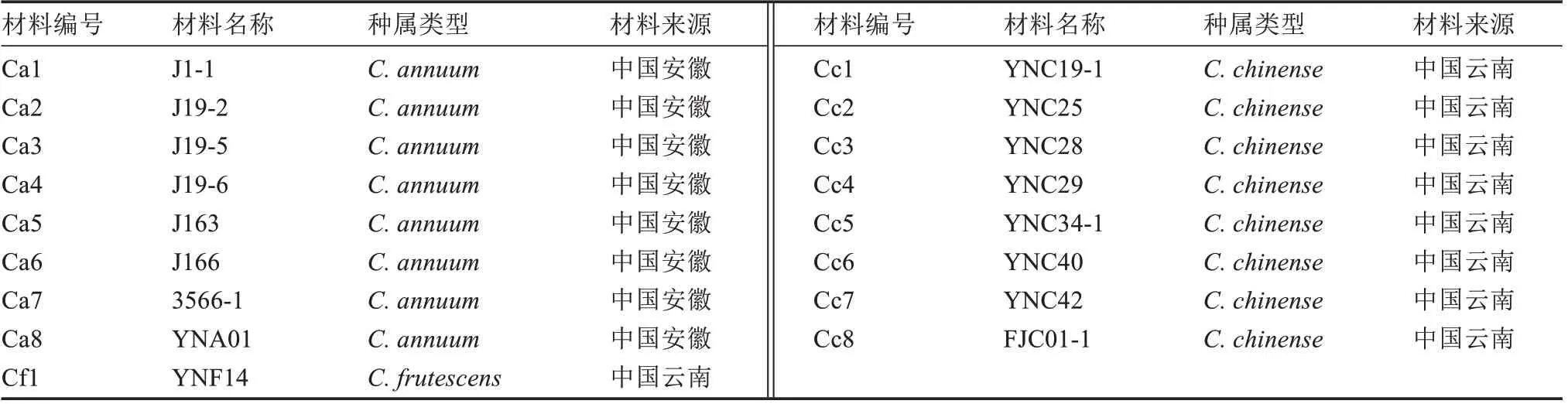
表1 用于遠(yuǎn)緣雜交的辣椒親本信息Table 1 Capsicum parental information for distant hybridization

圖1 3 個(gè)辣椒栽培種的典型圖片F(xiàn)ig.1 Typical pictures of three Capsicum species
1.2 方法
1.2.1 種間雜交 不同辣椒材料進(jìn)入盛花期后選配了18 對(duì)正反交組合(表2)。雜交授粉時(shí)選擇適齡花蕾徒手去雄,授粉在晴天07:00 至10:30、15:30 至18:00 進(jìn)行,授粉后掛牌標(biāo)記,第2 天重復(fù)授粉1 次,授粉后14 d 統(tǒng)計(jì)坐果率。
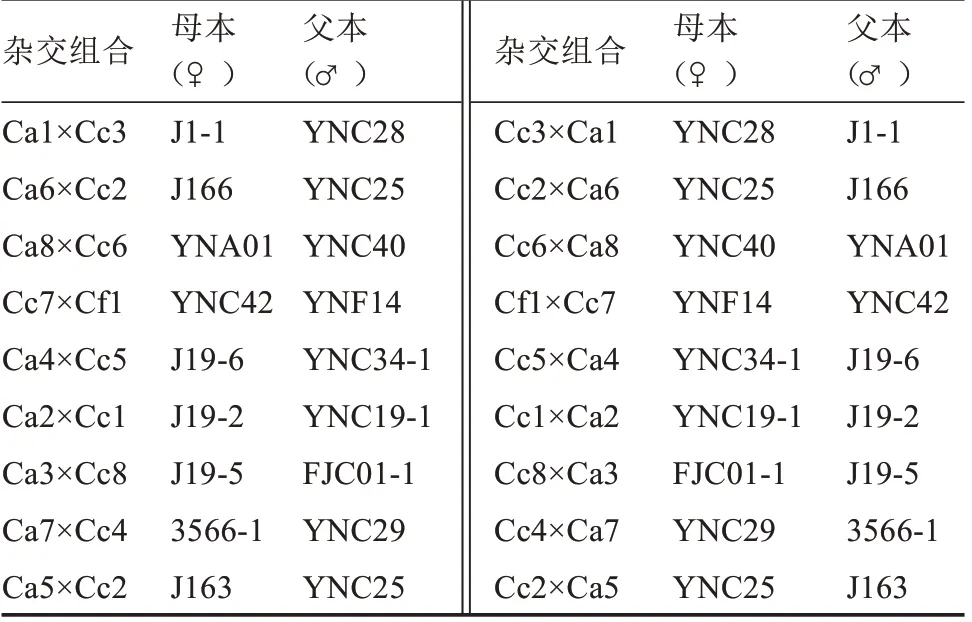
表2 3 個(gè)栽培種辣椒種間雜交的正反交組合Table 2 Reciprocal interspecific hybridization combinations of Capsicum species
1.2.2 消毒及拯救(萌發(fā))培養(yǎng)基篩選 授粉后第34 d,選取Ca4×Cc5 未成熟的F1幼胚在無菌組培室的潔凈操作臺(tái)上進(jìn)行消毒處理,流水沖洗表面灰塵后用75% 的酒精進(jìn)行果實(shí)表面消毒1 min,剖開果實(shí)內(nèi)腔胎座及幼胚消毒30 s,再用0.1% 的氯化汞浸泡3 min,無菌水清洗3~5 次。將剝離的幼胚分別接種到MF0(MS+30 g·L-1蔗糖+6 g·L-1瓊脂,pH 6.0)、MF1、MF2、MF3、MF4、MF5、MF6培養(yǎng)基上(表3),從而篩出最優(yōu)萌發(fā)培養(yǎng)基。每個(gè)處理6 次重復(fù),每瓶接種5 個(gè)幼胚,置于(25±1)℃條件下培養(yǎng),光照度為2500 lx,光周期為12 h/12 h(光照/黑暗),接種30 d 后統(tǒng)計(jì)胚萌發(fā)率。

表3 胚胎萌發(fā)及幼苗生根培養(yǎng)基Table 3 The medium of embryonic germination and seedling rooting
1.2.3 不同胚拯救時(shí)期篩選 試驗(yàn)設(shè)置了5 個(gè)處理時(shí)期,即在授粉后30、32、34、36、38 d 時(shí)剪下幼果,放入篩選出的最佳培養(yǎng)基中進(jìn)行胚胎拯救。具體操作為:在對(duì)應(yīng)時(shí)間分別從每個(gè)雜交組合植株上取2 個(gè)青果,取出內(nèi)部全部F1幼胚,消毒及培養(yǎng)條件同1.2.2,每個(gè)處理均接種3 瓶,每瓶2~9 個(gè)幼胚(視果實(shí)內(nèi)幼胚多少而定)。培養(yǎng)30 d 后統(tǒng)計(jì)胚萌發(fā)率。
1.2.4 生根培養(yǎng)基篩選 試驗(yàn)共設(shè)10 個(gè)處理組合,以不加激素的SG0(MS+30 g·L-1蔗糖+6 g·L-1瓊脂,pH 6.0)為對(duì)照,用萘乙酸(NAA)、吲哚丁酸(IBA)、吲哚乙酸(IAA)3 種生長(zhǎng)類激素各3 個(gè)濃度水平進(jìn)行搭配(表3)。將拯救成功的高2~4 cm 的Ca4×Cc5 幼苗轉(zhuǎn)接至SG 培養(yǎng)基中,培養(yǎng)條件同1.2.2。每個(gè)處理設(shè)置4 次重復(fù),每次重復(fù)接種3 株幼苗,接種30 d 后統(tǒng)計(jì)生根莖段數(shù)、生根率、每莖段生根數(shù)及平均根長(zhǎng)。
1.3 指標(biāo)測(cè)定
1.3.1 坐果指標(biāo)測(cè)定 授粉后記錄各組合授粉總花數(shù),于第14 天調(diào)查坐果情況,計(jì)算坐果率。
1.3.2 胚萌發(fā)指標(biāo)測(cè)定 將幼胚接入培養(yǎng)基后,于第30 天觀察幼胚的萌發(fā)情況,統(tǒng)計(jì)萌發(fā)幼胚數(shù),幼胚萌發(fā)以長(zhǎng)出子葉為標(biāo)準(zhǔn)。
1.3.3 根系各指標(biāo)測(cè)定 將莖段接入不同的SG 培養(yǎng)基后,于第30 天對(duì)各處理的生根數(shù)、生根率、平均每莖段生根數(shù)、平均根長(zhǎng)進(jìn)行統(tǒng)計(jì)分析。
1.4 數(shù)據(jù)統(tǒng)計(jì)與分析
采用Microsoft Excel 2016 進(jìn)行數(shù)據(jù)處理和圖表制作,采用DPS7.05 進(jìn)行完全隨機(jī)單因素試驗(yàn)方差分析。
2 結(jié)果與分析
2.1 不同種間雜交組合的坐果率與種子數(shù)統(tǒng)計(jì)
由表4 可知,18 個(gè)正反交組合,共授粉花數(shù)1451 朵,平均坐果率總體較低,僅為7.58%,平均每果種子數(shù)為14.82 粒。但不同栽培種做母本時(shí),坐果數(shù)(坐果率)存在差異。C.annuum做母本時(shí)共授粉899 朵花,坐果數(shù)為72 個(gè),平均坐果率為8.01%,共獲得種子1336 粒,平均每果種子數(shù)18.56 粒;C.chinense為母本時(shí)共授粉520 朵花,坐果數(shù)為28個(gè),平均坐果率為5.38%,共獲得種子223 粒,平均每果種子數(shù)7.96 粒;C.frutescens做母本時(shí),共授粉32 朵花,坐果數(shù)為10 個(gè),坐果率31.25%,獲得種子數(shù)為71 粒,平均每果種子數(shù)7.10 粒。3 個(gè)栽培種分別做母本時(shí),C.frutescens的坐果率最高,但其產(chǎn)生的種子數(shù)最少;C.annuum做母本時(shí)坐果率次之,其產(chǎn)生種子數(shù)最多;C.chinense做母本時(shí),坐果率最低,所產(chǎn)生的種子數(shù)介于C.frutescens和C.annuum之間。
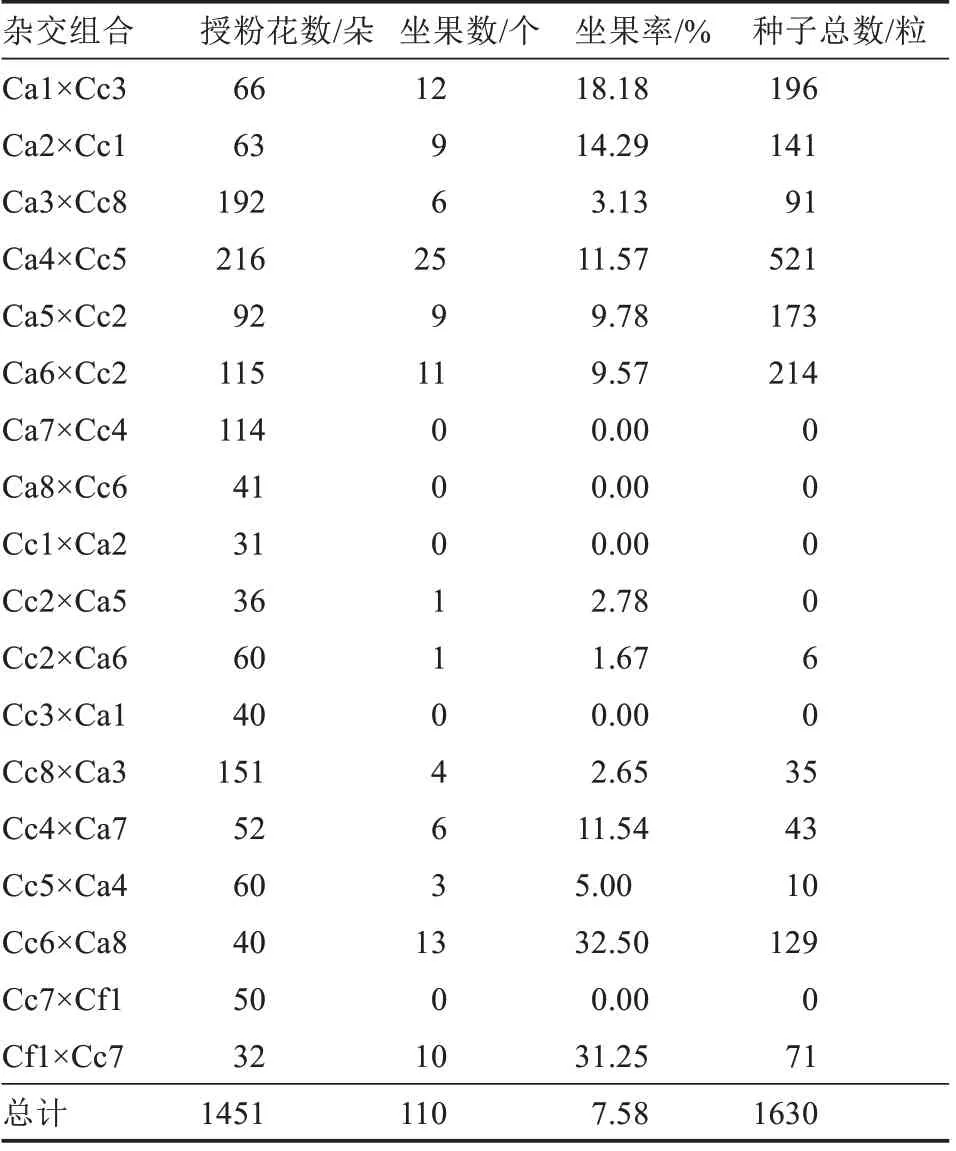
表4 正反交授粉及坐果統(tǒng)計(jì)Table 4 Pollination and fruit-setting of reciprocal interspecific
同時(shí),不同的雜交組合,正反交坐果率也不同。雜交組合Ca1×Cc3、Ca2×Cc1 正交時(shí)均能產(chǎn)生雜交果實(shí),坐果率分別為18.18%、14.29%,反交時(shí)無果實(shí)產(chǎn)生;Ca8×Cc6、Cc7×Cf1、Ca7×Cc4 正交時(shí)均無果實(shí),反交則有果實(shí),坐果率分別為32.50%、31.25%、11.54%。Ca6×Cc2、Ca4×Cc5、Ca3×Cc8、Ca5×Cc2 正反交時(shí)均有少數(shù)雜交果實(shí)產(chǎn)生,正交坐果率為3.13%~11.57%,反交坐果率為1.67%~5.00%。
2.2 胚胎拯救的萌發(fā)培養(yǎng)基篩選
將Ca4×Cc5 授粉后34 d 的幼胚分別接種于7種培養(yǎng)基上,結(jié)果如表5 所示,幼胚在MF0、MF1、MF2及MF4上萌發(fā)率相同,MF5次之,MF6的胚發(fā)育率較低,MF3胚萌發(fā)率為0,推測(cè)可能是激素濃度過高,抑制了胚的萌發(fā)。因此,從試驗(yàn)成本方面考慮,選擇MF0作為最適合幼胚萌發(fā)的培養(yǎng)基,即不需要添加激素,胚胎拯救在MF0培養(yǎng)基上萌發(fā)情況見圖2。
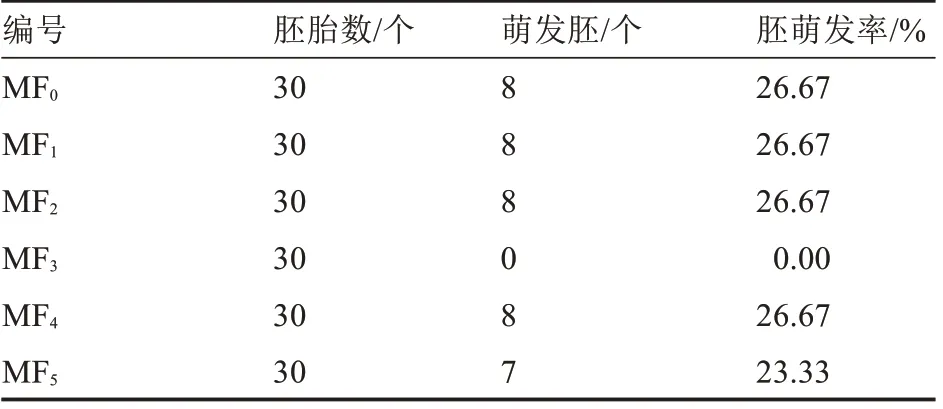
表5 培養(yǎng)基對(duì)辣椒幼胚胚胎拯救的影響Table 5 Effect of culture medium on embryo rescue of Capsicum

圖2 不同發(fā)育時(shí)期的辣椒胚胎拯救Fig.2 Embryo germination and plantlet growth in different development stages
2.3 辣椒不同胚齡對(duì)胚胎拯救的影響
由于該部分試驗(yàn)對(duì)果實(shí)數(shù)量有要求,故選擇了果實(shí)數(shù)量大于等于10(每個(gè)拯救時(shí)期需要2 個(gè)青果)的5 個(gè)雜交組合進(jìn)行試驗(yàn)。由表6 可知,除Ca6×Cc2 組合外,其他4 個(gè)組合胚的萌發(fā)率在胚胎拯救期間隨時(shí)間的延長(zhǎng)均呈先上升后下降的趨勢(shì),34 d 的幼胚萌發(fā)率最高,為45.45%,38 d 有3 個(gè)雜交組合的幼胚萌發(fā)率為0,其原因可能為取材時(shí)間過晚,胚已死亡。在Ca1×Cc3、Ca6×Cc2、Cc6×Ca8、Ca4×Cc5 雜交組合中,34 d 胚齡的胚萌發(fā)率均最高,分別為31.58%、34.62%、45.45%、36.00%,僅在Cf1×Cc7 雜交組合中,34 d 胚齡的幼胚萌發(fā)率比32 d 胚齡低20.83%。可見,C.annuum或C.chinense做母本時(shí),34 d 胚齡的幼胚最適合于拯救;而當(dāng)C.frutescens做母本時(shí),32 d 胚齡的幼胚最適合拯救。雜交組合Cf1×Cc7 整個(gè)幼胚拯救期間的胚平均萌發(fā)率最低,為11.43%,不同栽培種做母本時(shí),其胚的萌發(fā)率有所不同。
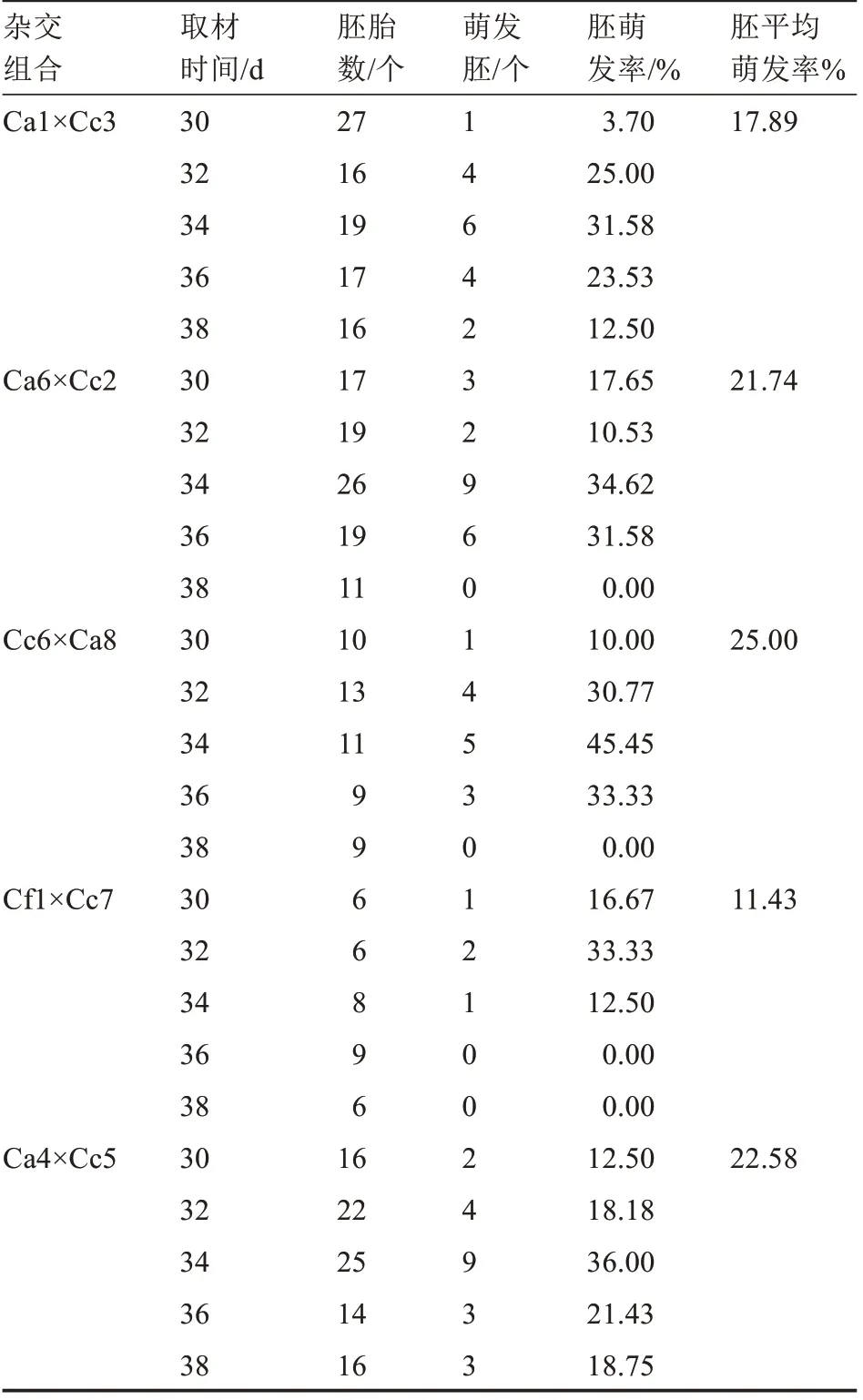
表6 不同胚齡對(duì)辣椒胚胎拯救的影響Table 6 Effects of different embryo ages on embryo rescue of Capsicum
2.4 生根培養(yǎng)基篩選
由表7 可知,以SG0為基本培養(yǎng)基,添加3 個(gè)不同種類3 種不同濃度的生長(zhǎng)類激素的培養(yǎng)基,其生根效果均存在差異。生根莖段數(shù)、生根率、平均每莖段生根數(shù)和平均根長(zhǎng)均以SG3表現(xiàn)最優(yōu),平均每莖段生根數(shù)和平均根長(zhǎng)顯著高于其他處理。由圖3可知,添加相同濃度不同種類的激素誘導(dǎo)的根形態(tài)也存在差異。添加0.5 mg·L-1NAA 的SG3表現(xiàn)為根多且粗壯,添加0.5 mg·L-1IBA 的SG6根較細(xì)且生根率低,添加0.5 mg·L-1IAA 的SG9表現(xiàn)為根少且長(zhǎng)短不均,以MS 為培養(yǎng)基時(shí),SG0表現(xiàn)為根少且細(xì)長(zhǎng)。綜上所述,添加0.5 mg·L-1NAA 的SG3培養(yǎng)基為辣椒組培苗生根最理想的培養(yǎng)基。

表7 不同濃度激素對(duì)辣椒組培苗生根影響Table 7 Effects of different concentrations of hormones on rooting of Capsicum tissue culture seedlings
3 討論與結(jié)論
胚敗育是辣椒種間雜交的一個(gè)共同特征[10]。在多數(shù)遠(yuǎn)緣雜交中,通常會(huì)收獲到癟的或退化的種子,這一現(xiàn)象被認(rèn)為是受精后發(fā)育障礙導(dǎo)致的[11]。Martins 等[4]從C.annuum×C.baccatum組合中得到了9 粒種子,但有7 粒沒有發(fā)育成完整植株;Yoon等[12]觀察到C.annuum×C.baccatum雜交后胚敗育發(fā)生在授粉后15 d,另一個(gè)現(xiàn)象是種子在成熟前便開始干燥失去活力。筆者試驗(yàn)中的18 對(duì)雜交組合,部分授粉后不能坐果,故無法對(duì)其進(jìn)行胚胎拯救。另一部分授粉后可坐果,但因不同栽培種間親緣關(guān)系較遠(yuǎn),且存在雜交障礙,為防止果實(shí)內(nèi)部幼胚在發(fā)育后期死亡而無法獲得雜種植株,故選擇在特定時(shí)間對(duì)其進(jìn)行胚胎拯救。此外,筆者還觀察到多數(shù)辣椒材料雜交后只有微小的種皮,部分種子中途褐化失去活力或沒有種子,但果實(shí)仍能正常生長(zhǎng)。Pickersgill[13]認(rèn)為,受精后胚敗育的合子障礙問題可以通過植物組織培養(yǎng),特別是胚胎拯救技術(shù)來解決。
基因型不同,種間雜交、胚胎拯救技術(shù)難易程度不同。程志芳等[14]采用常規(guī)授粉技術(shù)對(duì)5 個(gè)辣椒栽培品種進(jìn)行了種間正反交,僅從一小部分的辣椒雜交組合中獲得了雜種植株。筆者的研究中總體種間雜交平均坐果率低,但不同栽培種做母本時(shí)仍有相對(duì)較大的差異。C.annuum為母本時(shí),坐果率為0~18.18%,平均每果種子數(shù)為0~20.84 粒;C.chinense為母本時(shí),坐果率為0~32.50%,平均每果種子數(shù)0~9.92 粒;C.frutescens為母本時(shí),坐果率為31.25%,平均每果種子數(shù)7.10 粒。試驗(yàn)中正反交結(jié)實(shí)率差異較大,可見坐果率、果實(shí)內(nèi)種子數(shù)與父母本的基因型有關(guān)。此外,筆者的試驗(yàn)在溫度較高的溫室內(nèi)進(jìn)行,推測(cè)高溫可能會(huì)影響父本的花粉活力。
胚萌發(fā)是胚拯救的關(guān)鍵環(huán)節(jié)。培養(yǎng)基成分對(duì)幼胚萌發(fā)和存活有直接影響。據(jù)報(bào)道6-BA、NAA、GA3、ZT、IAA 對(duì)不同作物的幼胚萌發(fā)具有促進(jìn)作用[15-18],在辣椒胚胎拯救中常用的激素有IAA、GA3
[13,19]。但筆者研究發(fā)現(xiàn)不添加任何激素的MF0與添加IAA、6-BA 的處理無顯著差異,推測(cè)該階段導(dǎo)致幼胚敗育的關(guān)鍵是營(yíng)養(yǎng)虧缺,而不是激素水平的高低。
適宜的胚齡是胚胎拯救成功與否的最重要的影響因子之一。一般情況下,在雜種胚敗育之前將胚取出進(jìn)行離體培養(yǎng)可避免胚的早衰,而胚發(fā)育的越完全,拯救的成功率越高[20]。筆者的試驗(yàn)以30~38 d 胚齡的幼胚進(jìn)行離體培養(yǎng),其中C.annuum×C.chinense、C.chinense×C.annuum的雜交組合中34 d胚齡的胚萌發(fā)率最高,C.frutescens×C.chinense的雜交組合中32 d 的胚萌發(fā)率最高,而38 d 的胚萌發(fā)率普遍較低,主要表現(xiàn)為幼胚褐化,其主要原因推測(cè)為不同栽培種種間的雜交果實(shí)熟性有差異,導(dǎo)致酚類物質(zhì)積累不同,故胚胎拯救效果不佳。因此,針對(duì)不同的辣椒栽培種,仍需要探究適合幼胚培養(yǎng)的最佳胚齡。
綜上所述,試驗(yàn)中不同辣椒栽培種正反交組合坐果率整體較低但仍有差異。最佳胚萌發(fā)培養(yǎng)基選擇低成本的MF0(MS+30 g·L-1蔗糖+6 g·L-1瓊脂,pH 6.0),胚平均萌發(fā)率可達(dá)到20%。胚拯救適宜的取材時(shí)期應(yīng)在授粉后32~34 d,此時(shí)胚拯救成功率最高。生根培養(yǎng)基應(yīng)選擇SG3(MS+30 g·L-1蔗糖+6 g·L-1瓊脂+0.5 mg·L-1NAA,pH 6.0)。該體系適合本試驗(yàn)中3 個(gè)栽培種種間雜交幼胚的培養(yǎng),研究結(jié)果為加快辣椒新品種選育進(jìn)程奠定了基礎(chǔ)。

- 中國瓜菜的其它文章
- 從《農(nóng)業(yè)法律法規(guī)》看農(nóng)業(yè)法律體系的構(gòu)建
- 農(nóng)業(yè)新聞公共服務(wù)的理論與實(shí)踐
——評(píng)《農(nóng)業(yè)新聞:公共服務(wù)與話語創(chuàng)新的理論與實(shí)踐》 - 基于《農(nóng)產(chǎn)品包裝標(biāo)識(shí)概論》分析傳統(tǒng)文化元素在農(nóng)產(chǎn)品包裝視覺傳達(dá)設(shè)計(jì)中的應(yīng)用
- 開封秋大白菜輕簡(jiǎn)化栽培關(guān)鍵技術(shù)及經(jīng)濟(jì)效益分析
- 我國辛辣類蔬菜價(jià)格波動(dòng)預(yù)警分析
- 世界甜瓜貿(mào)易格局演變及對(duì)我國甜瓜貿(mào)易的啟示