甘藍型油菜酪氨酸代謝關(guān)鍵基因FAH的克隆、功能鑒定和表達分析
2023-11-23 09:11:20支添添周舟陳紀鵬韓成云
生物技術(shù)通報 2023年10期
支添添 周舟 陳紀鵬 韓成云
(1. 宜春學(xué)院生命科學(xué)與資源環(huán)境學(xué)院 宜春學(xué)院江西省作物生長發(fā)育調(diào)控重點實驗室,宜春 336000;2. 宜春學(xué)院化學(xué)與生物工程學(xué)院,宜春 336000)
酪氨酸降解途徑在動物中必不可少,若中斷將導(dǎo)致嚴重的代謝疾病[1-3]。這條途徑首先在動物和細菌中被發(fā)現(xiàn)[4-5],是一條非常重要的代謝途徑。Dixon等[6]證明擬南芥(Arabidopsis thaliana)中存在酪氨酸降解途徑的關(guān)鍵酶,并且和動物中有相同的功能。越來越多的證據(jù)表明,植物具有典型的酪氨酸分解代謝途徑[7-13]。延胡索酰乙酰乙酸酶(fumarylacetoacetate hydrolase, FAH)是酪氨酸代謝途徑的最后一個酶,F(xiàn)AH(SSCD1)基因的突變導(dǎo)致擬南芥在短日照下產(chǎn)生模擬病斑并激活植物防御[11-12]。
模擬病斑突變體是一類在沒有明顯的逆境、損傷或病原物侵害時,在葉片上能自發(fā)地形成類似病原物侵染后的壞死斑的突變體,這類突變體在植物中廣泛存在,如擬南芥、水稻(Oryza sativa)、玉米(Zea mays)、高粱(Sorghum bicolor)、小麥(Triticum aestivum)、大麥(Hordeum vulgare)等中均有報道[14-19]。這些突變體往往能激活植物體的系統(tǒng)獲得性抗性,通常具有與抗病反應(yīng)相關(guān)的細胞學(xué)和生物化學(xué)特征,對許多病原物表現(xiàn)出局部和系統(tǒng)抗性,是研究植物抗病機理的理想材料[20],如bon1(bonzai1)和cpr1/cpr30模擬病斑突變體對丁香假單胞菌有抗性[21-22]。水稻模擬病斑突變體oscul3a對稻瘟病菌和黃單胞菌均有很強的抗性[23],大麥mlo突變體對白粉病具有非特異性抗性[24],小麥模擬病斑突變體Ning7840和lm3分別對葉銹病和白粉病具有抗性[25-26]。
研究表明模擬病斑突變體的抗病性與植物抗病信號中的水楊酸(salicylic Acid, SA)、茉莉酸(jasmonic acid, JA)、乙烯(ethylene, ET)等信號分子均有相關(guān),這些植物激素不同程度地參與了抗病過程中的不同環(huán)節(jié),起著非常重要的作用[27-32]。前期研究證實sscd1突變體產(chǎn)生模擬病斑的同時積累JA,同時激活調(diào)控JA/ET信號途徑的部分病程相關(guān)基因PDF1.2,VSP1和SA病程相關(guān)基因PR1[12]。盡管sscd1中的模擬病斑的產(chǎn)生伴隨著SA誘導(dǎo)基因PR1的上調(diào),但與SA信號轉(zhuǎn)導(dǎo)無關(guān);而JA信號通路的中斷顯著抑制sscd1模擬病斑的產(chǎn)生[12]。這些結(jié)果說明FAH基因突變在短日照下激活擬南芥抗病信號途徑從而調(diào)控植物抗病。
甘藍型油菜(Brassica napus L.)是我國重要的油料作物,是第一大國產(chǎn)食用植物油來源和我國第二大飼用蛋白源[33]。然而,各種病害的頻繁發(fā)生嚴重影響著油菜的產(chǎn)量和品質(zhì)[34]。因此,挖掘油菜抗病相關(guān)遺傳資源、提高油菜抗病性對于油菜生產(chǎn)具有重要意義。甘藍型油菜是白菜(Brassica rapa, AA, 2n=20)和甘藍(Brassica oleracea, CC,2n=18)通過自然雜交形成的異源四倍體作物(AACC,2n=4X=38)[35-36],白菜和甘藍的基因組在古老的多倍化事件中都經(jīng)歷了3倍化的過程[37]。因此,甘藍型油菜相比擬南芥,具有更加復(fù)雜的基因組特征并且大多數(shù)基因具有多個同源拷貝,具有冗余或不同的功能[38]。盡管在擬南芥中已證實FAH基因突變在短日照下產(chǎn)生模擬病斑并激活抗病反應(yīng),但鑒于甘藍型油菜相比擬南芥基因組更復(fù)雜,仍需要鑒定不同F(xiàn)AH基因在甘藍型油菜中的功能并分析其各自的表達模式。
本研究從甘藍型油菜克隆了2個與擬南芥高度同源的FAH基因BnaA06g38260D(BnaA06FAH)和BnaC05g49430D(BnaC05FAH),通過生物信息學(xué)分析其編碼的蛋白質(zhì)序列和進化關(guān)系;構(gòu)建過表達載體轉(zhuǎn)化擬南芥突變體sscd1進行功能驗證;進一步對BnaA06FAH和BnaC05FAH啟動子進行克隆,探究其序列特征和潛在的響應(yīng)元件,構(gòu)建含有報告基因GUS的植物表達載體,通過GUS染色揭示其表達模式,為深入研究BnaA06FAH 和BnaC05FAH在甘藍型油菜中的作用和功能奠定基礎(chǔ)。
1 材料與方法
1.1 材料
甘藍型油菜品種為‘westar’、擬南芥(Arabidopsis thaliana)生態(tài)型為哥倫比亞(Columbia, Col?0),突變體sscd1、大腸桿菌菌株DH5α、農(nóng)桿菌菌株GV3101、啟動子分析載體pCAMBIA1301和過表達載體pBI121均由宜春學(xué)院江西省作物生長發(fā)育調(diào)控重點實驗室保存。擬南芥sscd1突變體在短日照下產(chǎn)生模擬病斑[11]。
1.2 方法
1.2.1 DNA、RNA的提取及cDNA合成 待甘藍型油菜長到四葉期,取適量的新鮮的葉片用液氮研磨,用高效植物基因組DNA提取試劑盒(北京天根生化科技有限公司)提取葉片基因組DNA,使用分光光度計(ND?1000, NanoDrop, Thermo Fisher Scientific)檢測濃度后儲存于?20℃,用作克隆啟動子序列的模板;用Trizol(RNAiso Plus, 寶生物工程有限公司)提取總RNA,進一步用DNase I(RNase Free,Thermo Fisher Scientific)去除基因組DNA后利用反轉(zhuǎn)錄試劑盒ReverTraAce qPCR RT試劑盒(perfect real time, Toyobo),將RNA反轉(zhuǎn)錄成cDNA,用作克隆CDS序列的模板。
1.2.2 BnaA06FAH和BnaC05FAH的CDS序列和啟動子的克隆以及載體的構(gòu)建與轉(zhuǎn)化
1.2.2.1 引物設(shè)計及目的片段的擴增 在甘藍型油菜數(shù)據(jù)庫(https://www.genoscope.cns.fr/brassicanapus/)中獲得BnaA06FAH和BnaC05FAH的cDNA預(yù)測序列。利用軟件Primer premier 5.0在2個基因非編碼區(qū)設(shè)計特異引物(表1),用TaKaRa高保真DNA聚合酶進行PCR擴增,以擴增得到的cDNA為模板,在BnaA06FAH和BnaC05FAH上游引物的5′添加Xba I(TCTAGA)限制性內(nèi)切酶和保護堿基(GC)以及下游引物5′添加Sac I(GAGCTC)限制性內(nèi)切酶和保護堿基(C)(引物見表1),再次進行CDS序列的PCR擴增,目的片段大小預(yù)期為1 266 bp,經(jīng)1.0%瓊脂糖凝膠電泳檢測、回收并純化,測序。

表1 試驗所用引物Table 1 Primers used in the experiment
在NCBI中獲得BnaA06FAH和BnaC05FAH翻譯起始位點ATG上游1.8 kb左右的DNA序列,根據(jù)啟動子序列,利用軟件Primer premier 5.0分別設(shè)計擴增引物(表1),在上下游引物的5′端分別加上Xba I和Bgl II酶切位點及其保護堿基,以‘westar’總DNA為模板進行PCR擴增,目的片段大小預(yù)期為1 980 bp(BnaA06FAH)和1 708 bp(BnaC05FAH),經(jīng)1.0%瓊脂糖凝膠電泳檢測、回收并純化,測序。
1.2.2.2 35S∷BnaC05FAH和35S∷BnaA06FAH重組載體的構(gòu)建 用限制性內(nèi)切酶Xba I和Sac I對已測序驗證的BnaA06FAH和BnaC05FAH的CDS序列和pBI121載體分別進行雙酶切,然后將目的片段和載體進行連接,使目的基因替換原載體上的GUS,構(gòu)建35S∷BnaC05FAH和35S∷BnaA06FAH過表達載體,轉(zhuǎn)化大腸桿菌DH5α感受態(tài)細胞,挑取菌落進行PCR鑒定,對鑒定的陽性菌落進行測序,以確保載體構(gòu)建的準確性。
1.2.2.3 pBnaC05FAH∷GUS和pBnaA06FAH∷GUS重組載體的構(gòu)建 用限制性內(nèi)切酶Xba I和Bgl II對已測序驗證的BnaA06FAH和BnaC05FAH的啟動子序列及pCAMBIA1301載體進行雙酶切,然后進行連接,用BnaA06FAH和BnaC05FAH啟動子替換pCAMBIA1301載體上原有35S啟動子。通過熱激法將連接產(chǎn)物轉(zhuǎn)化至大腸桿菌DH5α,隨機挑選陽性克隆進行菌落PCR及酶切檢測,測序。將測序驗證正確的陽性克隆進行搖菌和質(zhì)粒提取。
1.2.2.4 擬南芥的遺傳轉(zhuǎn)化及篩選 用CaCl2凍融法將已驗證的重組質(zhì)粒轉(zhuǎn)入農(nóng)桿菌GV3101,挑選菌落PCR驗證正確的農(nóng)桿菌單菌落進行培養(yǎng),28℃、220 r/min振蕩培養(yǎng)18-24 h。7 352 r/min離心活化菌液2 min,棄上清,用農(nóng)桿菌懸浮液(5%蔗糖+1/2 MS+0.02% Silwet?L77+0.01% 6?BA)將菌體重懸至OD600為0.6-1.0。利用浸花法轉(zhuǎn)化擬南芥,收取T0代種子,在含有潮霉素(25 mg/L)或卡那霉素(50 mg/L)的MS培養(yǎng)基上篩選成功轉(zhuǎn)入表達載體的轉(zhuǎn)基因植株,將篩選到的轉(zhuǎn)基因植株T0代抗性苗移入土中,后代單株收種。挑選T2代抗性符合3∶1分離比的轉(zhuǎn)基因株系,用GUS組織化學(xué)法檢測啟動子活性。
1.2.3 生物信息學(xué)分析 PLANTCARE在線分析啟動子序列包含的順式作用元件(http://bioinformatics.psb.ugent.be/webtools/plantcare/html/)。用Bioxm2.7軟件分析BnaA06FAH和BnaC05FAH序列的開放閱讀框(open reading frame, ORF)。從NCBI數(shù)據(jù)庫中下載不同物種FAH蛋白的氨基酸序列,用DNAMAN 6.0軟件對氨基酸序列進行比對,用MEGA 11.0軟件對氨基酸序列進行系統(tǒng)進化樹分析。
1.2.4 GUS組織染色 根據(jù)潮霉素抗性在MS固體培養(yǎng)基上篩選pBnaC05FAH∷GUS和pBnaA?06FAH∷GUS轉(zhuǎn)基因擬南芥至T3代,用GUS染色液(GUS Blue KIT, 北京華越洋生物科技有限公司)對MS培養(yǎng)基上9和14 d的擬南芥進行GUS染色,并將14 d幼苗移栽到土中生長至抽薹開花,剪下莖前端進行染色:將擬南芥被配置好的染色液完全覆蓋,抽真空2-3次,每次10 min后,用錫箔紙包好,37℃孵育24 h,70%乙醇脫色3 d,每天更換到脫色完全。染色結(jié)束后將植株置于體式顯微鏡下(Nikon SMZ800N)觀察并拍照記錄。
1.2.5 RT?qPCR 采用SYBR qPCR混合試劑盒(Toyobo)和ABI 7300序列檢測系統(tǒng)(Applied Biosystems, Foster City, CA, USA)按照說明書進行實時定量PCR。RT?qPCR檢測的基因引物見表1,以ACTIN2作為內(nèi)參。每個樣本的基因表達量在3次分析重復(fù)中計算,相對表達量用2-ΔΔCt方法定量。所有試驗至少重復(fù)3次。表中的數(shù)據(jù)表示為RT?qPCR中3次生物重復(fù)的平均值±SE。
2 結(jié)果
2.1 BnaA06FAH和BnaC05FAH的CDS序列擴增
根據(jù)AtFAH氨基酸序列,通過BnIR(Brassica napus multi?omics information resource)進行BLAST比對,獲得同源性最高的2個甘藍型油菜(Brassica napus L.)FAH基因:BnaA06g38260D(BnaA06FAH)和BnaC05g49430D(BnaC05FAH)。根據(jù)基因cDNA序列設(shè)計引物,以甘藍型油菜品種‘westar’葉片cDNA為模板進行擴增,獲得BnaA06FAH和BnaC05FAH的CDS序列(圖1),測序。
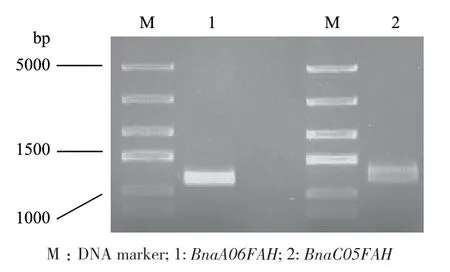
圖1 BnaA06FAH和BnaC05FAH CDS序列的PCR擴增Fig. 1 PCR products of CDS sequences of BnaA06FAH and BnaC05FAH
2.2 BnaA06FAH和BnaC05FAH的CDS序列生物信息學(xué)分析
測序后的CDS序列翻譯成氨基酸序列,與擬南芥AtFAH(NP 172669.2)進行同源比對,同源性分別為93.11%和92.40%(圖2?A)。基于BnaA06FAH和BnaC05FAH的氨基酸序列,與蘿卜、擬南芥、薺菜、亞麻芥、玉米和短花藥野生稻的FAH基因全長預(yù)測氨基酸序列構(gòu)建系統(tǒng)進化樹,結(jié)果(圖2?B)顯示,甘藍型油菜中該基因與白菜和甘藍的親緣關(guān)系最近,其次是蘿卜,再而是擬南芥,與單子葉植物的親緣關(guān)系較遠。

圖2 BnaA06FAH和BnaC05FAH氨基酸序列比對(A)和進化樹分析(B)Fig. 2 Amino acid sequences alignment(A)and phylogenetic tree(B)of BnaA06FAH and BnaC05FAH
2.3 BnaA06FAH和BnaC05FAH的功能驗證
為了驗證BnaA06FAH和BnaC05FAH的功能,構(gòu)建35S∷BnaA06FAH和35S∷BnaC05FAH過表達載體,通過浸花法轉(zhuǎn)入擬南芥sscd1突變體,以T2代純合株系的蓮座葉cDNA為模板,分別選擇4個獨立株系進行熒光定量PCR分析,結(jié)果(圖3)顯示,BnaA06FAH和BnaC05FAH在sscd1突變體中本身無表達,其相對表達量為0;擬南芥突變體sscd1轉(zhuǎn)基因植株中35S∷BnaA06FAH?sscd17?2和35S∷BnaC05FAH?sscd14?11的相對表達量最高,作為后續(xù)試驗的研究材料。

圖3 過表達擬南芥株系中BnaA06FAH和BnaC05FAH的相對表達量Fig 3 Relative expressions of BnaA06FAH and BnaC05?FAH in overexpressed A. thaliana line
當在擬南芥sscd1突變體中過表達甘藍型油菜BnaA06FAH和BnaC05FAH,sscd1突變體在短日照下的模擬病斑完全被抑制(圖4)。結(jié)果表明,甘藍型油菜BnaA06FAH和BnaC05FAH的功能都與擬南芥AtFAH相似。
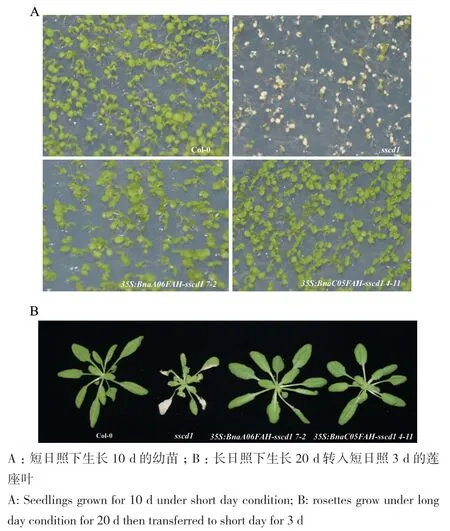
圖4 短日照下過表達BnaA06FAH和BnaC05FAH抑制擬南芥突變體sscd1的模擬病斑Fig. 4 Overexpressions of BnaA06FAH and BnaC05FAH repress the simulated disease spot in Arabidopsis mutant sscd1 under short-day condition
2.4 BnaA06FAH和BnaC05FAH啟動子序列的擴增和生物信息學(xué)分析
在甘藍型油菜數(shù)據(jù)庫和NCBI中獲得BnaA?06FAH和BnaC05FAH對應(yīng)的基因組序列,選取ATG上游1.8 kb左右的序列。以甘藍型油菜‘westar’基因組DNA為模板,設(shè)計特異性的引物進行PCR擴增(表1),分別獲得BnaA06FAH和BnaC05FAH的啟動子片段(圖5)。在TAIR(http://www.arabi?dopsis.org/)網(wǎng)站獲得擬南芥AtFAH基因(AT1G12050)ATG上游1.8 kb的啟動子序列,通過PlantCARE(http://bioinformatics.psb.ugent.be/webtools/plantcare/html/)數(shù)據(jù)庫在線分析,并與BnaA06FAH和BnaC?05FAH啟動子順式作用元件進行比較。結(jié)果發(fā)現(xiàn),BnaA06FAH、BnaC05FAH和AtFAH啟動子序列除了都含有基本轉(zhuǎn)錄元件CAAT?box、TATA?box外,還含有多種應(yīng)答元件。應(yīng)答元件又可分為生長發(fā)育、光響應(yīng)、脅迫響應(yīng)和激素響應(yīng)4類元件(圖6),說明甘藍型油菜BnaA06FAH和BnaC05FAH和擬南芥AtFAH的表達都可能響應(yīng)光、植物激素和脅迫的誘導(dǎo)。生長發(fā)育類別中,涉及胚乳表達(AAGAA?mo?tif)、晝夜節(jié)律(circadian)和玉米醇溶蛋白代謝(O2?site)等,其中O2?site占比例最高,為43%。光響應(yīng)有12種順式作用元件,包括AE?box、AT1?motif、ATCT?motif、Box 4、G?Box、G?box、GA?motif、GT1?motif、Sp1、TCT?motif和MRE,其中G?box占比最大,為21%。脅迫響應(yīng)類別有6種順式作用元件,分別與防御應(yīng)激(STRE、as?1)、厭氧誘導(dǎo)(ARE)、脫水(DRE core)、創(chuàng)傷(WRE3和WUN?motif)和病原誘導(dǎo)(W?box)相關(guān),其中as?1和WRE3所占的比例最高,為32%。激素響應(yīng)類別中的元件包括脫落酸(ABRE、ABRE3a、ABRE4)、生長素(TGA?element、TGA?box)、赤霉素(GARE?motif)、茉莉酸甲酯(CGTCA?motif、TGACG?motif)應(yīng)答元件,其中響應(yīng)茉莉酸甲酯的順式作用元件占比最大,為25%。另外,這些應(yīng)答元件還參與抗病響應(yīng),如W?box、ERE、GT1?motif、G?box、as?1、TGACG?motif和CGTCA?motif,其中BnaA06FAH、BnaC05FAH和擬南芥AtFAH共有的啟動子順式作用元件有as?1、GT1?motif、TGACG?motif和CGTCA?motif,說明無論擬南芥還是甘藍型油菜FAH基因的表達很可能與植物抗病性相關(guān)。
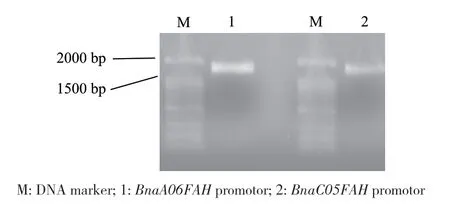
圖5 BnaA06FAH和BnaC05FAH啟動子的PCR擴增產(chǎn)物Fig. 5 PCR products of BnaA06FAH and BnaC05FAH promotors

圖6 AtFAH、BnaC05FAH和BnaA06FAH啟動子應(yīng)答元件分析Fig. 6 Analysis of promoter response elements of AtFAH, BnaC05FAH and BnaA06FAH
BnaA06FAH相比BnaC05FAH與擬南芥AtFAH啟動子共同擁有更多順式作用元件,如激素響應(yīng)順式作用元件(ABRE、ABRE3a、ABRE4)、光響應(yīng)元件(Sp1和G?box)、脫水響應(yīng)元件(DRE core)和創(chuàng)傷響應(yīng)元件(WRE3, 表2)。除了兩者共同的啟動子順式元件,兩者都存在特異的啟動子順式作用元件:BnaC05FAH特異的大部分都是光響應(yīng)元件(如AE?box、GA?motif和AT1?motif);而BnaA06FAH沒有光響應(yīng)元件,有激素防御相關(guān)(GARE?motif、TGA?element、W?box)誘導(dǎo)和與生長發(fā)育相關(guān)的順式調(diào)控元件(表3)。說明甘藍型油菜BnaA06FAH和BnaC05FAH的表達調(diào)控可能存在差異。
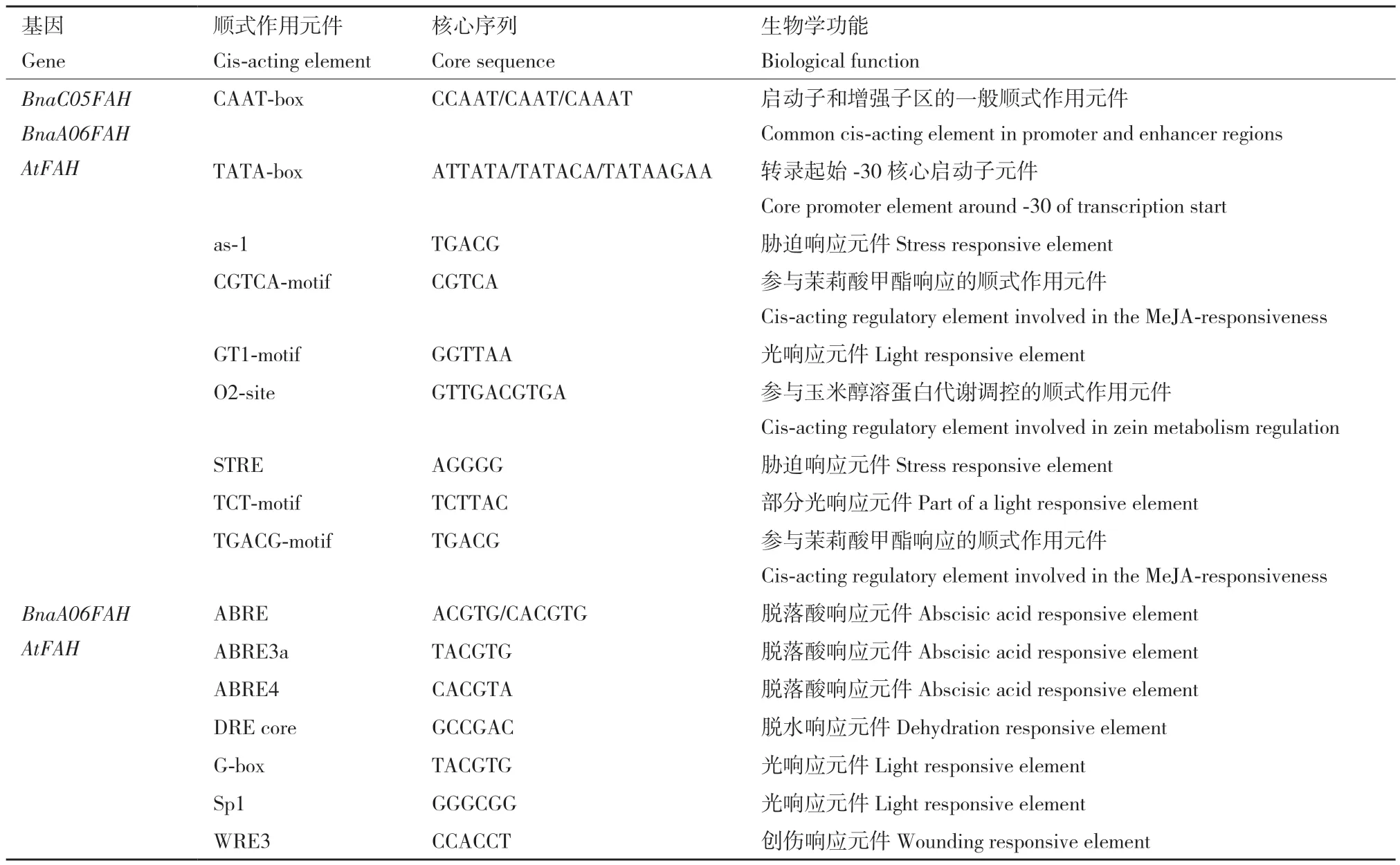
表2 BnaC05FAH、BnaA06FAH和AtFAH啟動子區(qū)共同順式作用元件Table 2 Common cis-acting elements in BnaC05FAH, BnaA06FAH and AtFAH promoter regions

表3 BnaC05FAH、BnaA06FAH和AtFAH啟動子區(qū)包含的不同順式作用元件Table 3 Different cis-acting elements in BnaC05FAH, BnaA06FAH and AtFAH promoter regions
2.5 BnaA06FAH和BnaC05FAH表達模式分析
為了探明BnaA06FAH和BnaC05FAH的表達模式,構(gòu)建BnaA06FAHPro∷GUS和BnaC05FAH?Pro∷GUS融合表達載體,通過電擊法轉(zhuǎn)入根癌農(nóng)桿菌GV3101中,并通過農(nóng)桿菌介導(dǎo)的浸花法轉(zhuǎn)化野生型擬南芥Col?0,利用潮霉素抗性篩選轉(zhuǎn)基因擬南芥至T3代,同時以未轉(zhuǎn)入任何載體的GV3101菌株侵染的擬南芥作為陰性對照(CK)。分別對生長9和14 d的T3代陽性和陰性植株幼苗、花和角果進行GUS組織化學(xué)染色,在實體顯微鏡下觀察染色情況。結(jié)果(圖7)顯示,在9 d的幼苗中,轉(zhuǎn)化BnaC05FAHPro∷GUS和BnaA06FAHPro∷GUS的陽性植株都在子葉、葉柄、下胚軸和根部位檢測到GUS的表達;待植株生長至14 d,BnaC05FAHPro驅(qū)動的GUS幾乎在所有部位都有表達,而BnaA06FAHPro∷GUS轉(zhuǎn)基因植株中GUS表達集中在子葉、幼嫩的真葉、下胚軸以及根。對花的染色結(jié)果顯示,BnaC05FAHPro∷GUS轉(zhuǎn)基因植株中GUS在莖稈、花萼、花絲,雌蕊的柱頭、花柱和子房中表達,相比之下,BnaA06FAHPro∷GUS轉(zhuǎn)基因植株的表達部位只集中在莖稈、花萼、花柱和子房,而柱頭、花藥、花絲和花瓣都沒有表達;二者幼嫩角果里的胚珠都可以染色,但BnaC05FAHPro∷GUS轉(zhuǎn)基因植株角果的心皮上也有染色。表達強度上,轉(zhuǎn)化BnaA06FAH啟動子的陽性苗組織藍色斑點顏色在所有時期均比轉(zhuǎn)BnaC05FAH啟動子的陽性苗淺,說明BnaC05FAH啟動子對GUS的驅(qū)動強度明顯比BnaA06FAH強。綜上所述,BnaA06FAH和BnaC05FAH啟動子驅(qū)動GUS在擬南芥中的表達在無論其組織部位還是強度上都存在明顯差異。

圖7 甘藍型油菜BnaC05FAH、BnaA06FAH啟動子驅(qū)動GUS在擬南芥中的表達Fig. 7 GUS expressions driven by BnaC05FAH and BnaA?06FAH promoters from B. napus in Arabidopsis seedlings
3 討論
油菜是僅次于玉米和油棕的第三大重要油料作物[39]。甘藍型油菜具有株型高大或較高大,單產(chǎn)高和適應(yīng)性強等優(yōu)良特性,成為我國和全世界主要栽培和種植的油菜類型[40]。但是,在甘藍型油菜的生長過程中會遭受很多病害的威脅,目前,生產(chǎn)上主要釆用栽培和化學(xué)的方法防治病害,但其效果十分有限[41-42]。因此,選育抗病品種無疑成為油菜抗病育種最根本也是最有效的方法。然而,當前栽培的甘藍型油菜品種缺少相關(guān)抗性基因,嚴重制約著油菜抗病育種進程。近年來,植物功能基因組學(xué)和基因工程的研究發(fā)展迅猛,特別是在模式植物擬南芥中抗病相關(guān)基因以及抗病分子機制研究取得了突破性進展。在與擬南芥同科的甘藍型油菜中,克隆和研究抗病相關(guān)基因,對提高甘藍型油菜的抗病性具有重要的理論和實踐意義。
油菜與擬南芥同屬十字花科蕓薹屬植物,研究證明二者基因組的同源性很高。課題組前期研究證實酪氨酸降解途徑在植物中具有生物功能,編碼酪氨酸降解途徑FAH的基因SSCD1突變后導(dǎo)致擬南芥在短日照下產(chǎn)生模擬病斑[8],并激活JA/ET途徑和病程相關(guān)基因的表達[11]。本研究克隆兩個與擬南芥高度同源的甘藍型油菜FAH基因BnaA06FAH和BnaC05FAH,分析發(fā)現(xiàn)其編碼的氨基酸序列與擬南芥ATFAH的氨基酸序列相似性高達93.11%和92.40%,分子進化分析表明,BnaA06FAH和BnaC05FAH雖然分別與白菜和甘藍同源性最高,進化關(guān)系最近,但與擬南芥同樣具有較高同源性,并且BnaA06FAH和BnaC05FAH在雙子葉植物中親緣關(guān)系較近,而與單子葉植物的親緣關(guān)系較遠。通過構(gòu)建過表達載體,進一步驗證2個甘藍型油菜FAH基因的過表達都能抑制擬南芥突變體sscd1在短日照下模擬病斑的形成,說明FAH基因在甘藍型油菜中的功能很可能與擬南芥非常相似。另外,對甘藍型油菜和擬南芥FAH基因啟動子區(qū)域順式作用元件的分析表明,AtFAH和BnFAH的啟動子中包含7種參與抗病響應(yīng)的順式作用元件(W?box、ERE、GT1?motif、G?box、as?1、TGACG?motif和CGTCA?motif)以及植物激素響應(yīng)元件。W?box是一類植物病原誘導(dǎo)相關(guān)基因的順式作用元件[43-44],ERE元件能夠調(diào)控真菌誘導(dǎo)表達,分別存在于BnaA06FAH和BnaC05FAH啟動子中。G?box能與bZIP蛋白結(jié)合[45],分別存在于BnaA06FAH和AtFAH的啟動子中。三者的啟動子中都含有GT1?motif和as?1元件,GT1?motif的表達由病原體誘導(dǎo),部分與GT?1?like轉(zhuǎn)錄因子相互作用,GT?1?like轉(zhuǎn)錄因子結(jié)合病程相關(guān)PR?1a啟動子,影響水楊酸誘導(dǎo)基因的表達水平[46]。另外,三者啟動子都含有響應(yīng)JA的TGACG?motif和CGTCA?motif。JA響應(yīng)死體型病原菌的侵入,并且能誘導(dǎo)許多植物產(chǎn)生防衛(wèi)應(yīng)答反應(yīng)。在擬南芥中,酪氨酸代謝相關(guān)基因TAT3、HGO、MAAI以及FAH(SSCD1)的表達已證實受到JA的誘導(dǎo)[11]。綜上所述,甘藍型油菜FAH基因BnaA06FAH和BnaC05FAH的功能很可能與擬南芥相似,并且都與抗病和JA相關(guān)。
蕓薹屬是十字花科中包含許多重要經(jīng)濟作物的一個屬,包括白菜、甘藍、油菜等常見的蔬菜和油料作物,是農(nóng)業(yè)育種研究中非常重要的研究材料。甘藍型油菜由二倍體祖先種白菜(B. rapa, 2n=20,AA)和甘藍(B. oleracea, 2n=18, CC)經(jīng)過自然雜交形成的[27]。蘿卜是十字花科植物中與蕓薹屬親緣關(guān)系最近的物種[47],研究者普遍認為蕓薹屬及其近緣屬蘿卜屬起源于一個共同的祖先種,因此在進化樹分析中,甘藍型油菜FAH基因與白菜和甘藍親緣關(guān)系最近,其次是蘿卜,再者是擬南芥。白菜是蕓薹屬第一個完成基因組測序的栽培種,通過與擬南芥的比較基因組分析發(fā)現(xiàn),它們有著很好的共線性關(guān)系。甘藍也是一種重要的蔬菜作物,與白菜比較發(fā)現(xiàn)其基因組局部上存在著大量的染色體重排,特定通路內(nèi)基因的保留以及同源基因的表達也存在顯著差異[48]。甘藍型油菜BnFAH基因一個來源白菜(BnaA06FAH),另一個來源甘藍(BnaC05FAH)。啟動子順式作用元件分析結(jié)果表明,與BnaC05FAH相比,BnaA06FAH和擬南芥擁有更多相同的順式作用元件,此外,它們存在各自的特異順式元件。GUS組織化學(xué)染色結(jié)果顯示,BnaA06FAH和BnaC05FAH啟動子驅(qū)動GUS在不同生長時期擬南芥中表達的部位和強度都存在差異,BnaC05FAH在各種生長時期擬南芥中的表達都比BnaA06FAH強;在14 d蓮座葉、花及果莢中,不僅表達強而且表達的部位更多。綜上所述,甘藍型油菜BnaA06FAH和BnaC05FAH無論是啟動子順式元件還是啟動子驅(qū)動的組織表達上都存在差異,這些結(jié)果為深入研究甘藍型油菜BnaA06FAH和BnaC05FAH的功能奠定基礎(chǔ),為油菜抗病育種工作提供種質(zhì)資源。
4 結(jié)論
克隆獲得與擬南芥AtFAH同源性最高的2個甘藍型油菜FAH基因BnaA06FAH和BnaC05FAH,其編碼的氨基酸序列與擬南芥的同源性高達93.11%和92.40%。過表達BnaA06FAH和BnaC05FAH都能夠完全抑制擬南芥sscd1突變體在短日照下的模擬病斑表型。二者可能都與模擬病斑形成相關(guān)。甘藍型油菜FAH基因BnaA06FAH和BnaC05FAH的功能與擬南芥AtFAH高度相似,但兩者的啟動子在擬南芥中對下游基因的驅(qū)動無論在強度還是部位上均存在顯著差異。
