廣西外來入侵植物豚草的分布調查
2023-11-20 13:35:18黃小榮何峰彭玉華陳始貴
雜草學報 2023年2期
黃小榮 何峰 彭玉華 陳始貴
黃小榮,何? 峰,彭玉華,等. 廣西外來入侵植物豚草的分布調查[J]. 雜草學報,2023,41(2):29-34.
doi:10.19588/j.issn.1003-935X.2023.02.0004
摘要:2019—2022年,對廣西111個縣(市、區)的豚草分布進行樣帶調查。結果表明,橫州、興賓、上林、武宣、象州、鹿寨、賓陽、灌陽和隆林9個縣(市、區)有豚草發生;除了在興賓區持續蔓延之外,橫州市和上林縣成為豚草的新增暴發熱點,新熱點的傳播風險更大。裝卸場是豚草發生率和歸化率最高的生境,車輛傳播和污染泥土傳播是廣西豚草擴散的主要渠道。環境的物種豐富度與豚草蓋度呈負相關,但與豚草發生率和歸化率均無顯著關聯。桂中平原周邊的崇山峻嶺對豚草傳播起到隔離帶作用。南部沿海地區的北海、欽州和防城港均未發現豚草,這種情況可能與冬季氣候溫和有關。
關鍵詞:豚草;外來入侵物種;廣西;分布;春化
中圖分類號:S451;Q948.2? 文獻標志碼:A? 文章編號:1003-935X(2023)02-0029-06
Distribution of Alien Invasive Species Ambrosia artemisiifolia in Guangxi Area
HUANG Xiao-rong,HE Feng,PENG Yu-hua,CHEN Shi-gui
(Guangxi Academy of Forestry Sciences,Nanning 530002,China)
Abstract:A field survey was conducted in 111 counties of Guangxi area to investigate distribution of Ambrosia artemisiifolia. The results showed that A. artemisiifolia occurred in 9 counties,including Hengzhou,Xingbin,Shanglin,Wuxuan,Xiangzhou,Luzhai,Binyang,Guanyang and Longlin. In addition to the continued spread in Xingbin County,Hengzhou and Shanglin became the other two new outbreak hotspots of A. artemisiifolia,and the risk of transmission in the new hotspots was even greater. Loading yard was the habitat with the highest occurrence rate and naturalization rate of A. artemisiifolia,and vehicle and contaminated soil transport were the main channels of spread for A. artemisiifolia in Guangxi area. Species richness in the environment was negatively correlated with the coverage of A. artemisiifolia,but was not correlated with? occurrence rate or naturalization rate. Towering and steep mountains around Guizhong Plain acted as a natural barrier to prevent the spread of A. artemisiifolia. No A. artemisiifolia was found in Beihai,Qinzhou and Fangchenggang of the southern part coastal areas due to warm winter climate there.
Key words:Ambrosia artemisiifolia;alien invasive species;Guangxi;distribution;vernalization
收稿日期:2023-03-07
基金項目:國家林業局生物安全項目(編號:KJZXSA2018012、KJZXSA2019010、KJZXSA202008)。
作者簡介:黃小榮(1964—),女,廣西富川人,碩士,高級工程師,主要從事外來植物調查與研究。E-mail:huangxr2004@sina.com。
豚草(Ambrosia artemisiifolia L.)為雌雄同株、風媒傳粉的一年生草本植物,原產北美洲。近幾十年來,豚草在全球迅速傳播,已經擴散至80多個國家和地區,其中25個國家將其列為外來入侵物種[1]。在歐洲的豚草嚴重暴發區,豚草造成作物大量減產,高致敏花粉引發公共衛生問題[2-3]。
豚草20世紀40年代傳入我國[4-5],現在在上海、廣州[6]、新疆、黑龍江等地區均有分布[7]。豚草是一種喜熱植物[8]。Essl等發現,入侵奧地利的豚草主要分布在海拔低于400 m的低海拔地區[9]。據Bassett等報道,在恒溫條件下培養28 d的豚草種子萌發率很低(<8.7%),10 ℃ 16 h和30 ℃ 8 h交替溫度培養28 d的種子萌發率最高(75%)[10]。這表明豚草在大陸性氣候下生長最好[11]。廣西大部分內陸地區適合豚草生長發育。豚草于20世紀80年代傳入廣西來賓市興賓區[12-14],廣西其他地區未見豚草分布的報道。2019—2022年,筆者對廣西全區111個縣(市、區)進行了林業外來物種普查,分析了廣西豚草發生區和歸化區的生境特征,可以為清除零星豚草提供信息,有助于防范其蔓延擴散。
1? 材料與方法
1.1? 樣地概況與調查方法
在廣西111個縣(市、區)進行實地調查,側重調查桂中地區來賓市興賓區及相鄰的縣(市、區),以每個縣(市、區)的城區為中心,向東南西北4個方向沿公路行進至縣邊界,每個方向選3~4個地段作樣帶(寬10~50 m,長500~1 500 m)進行調查,樣帶大小以連續行走5 min無新增物種為度,共調查樣帶250個,樣帶間距在5 km以上。記錄樣帶所在地理位置和生境特征,以及樣帶內所有維管植物的種類、豐度、豚草蓋度和發生情況。樣帶內豚草數量大于50株,或零星塊狀分布出現3次的記為歸化;低于該標準的記為偶發或無[15]。公路邊的停車裝卸點、修車場、散貨露天堆場、木楞場等生境類型都歸入裝卸場。
1.2? 豚草識別
豚草植株高5~250 cm,莖直立,上部分枝被糙毛狀柔毛;莖下部葉對生,上部互生,2~3回羽狀分裂;頭狀花序單性,雌雄同株;雄性頭狀花序在枝頂排列成總狀花序[16]。雌頭狀花序2~5組簇生于雄頭狀花序下部葉腋處[11];總苞閉合,倒卵形,頂端有圍裹花柱的圓錐狀嘴部,在頂部以下有5~8個尖刺[17],每個總苞只有1朵小花[11];瘦果倒卵形,藏于堅硬的總苞中[17]。豚草每年11月底開始枯死,雄花脫落;次年3月種子萌發;營養生長期的豚草與伴生的青蒿、艾蒿極形似,但豚草葉片被毛、葉背青綠色;6月底開始頂生雄花穗現蕾,辨識變得容易。
1.3? 數據處理
數量分析是為了揭示環境因子與豚草分布的關系,因此剔除了未發現豚草的102個縣(市、區),只對本次調查發現豚草的9個縣(市、區)進行統計分析,共計82個樣帶。運算采用R 4.1.3的二元廣義線性模型(GLM)和混效模型,響應變量為豚草歸化時,用二元變量1和0分別代表歸化和其他;響應變量為豚草發生時,二元變量的1包括歸化和偶發[18]。解釋變量包括生境類型、縣(市、區)、道路類型、緯度、經度、海拔和物種個數,后4個變量先行對數轉換使其在同一數量級、權重相同。
2? 結果與分析
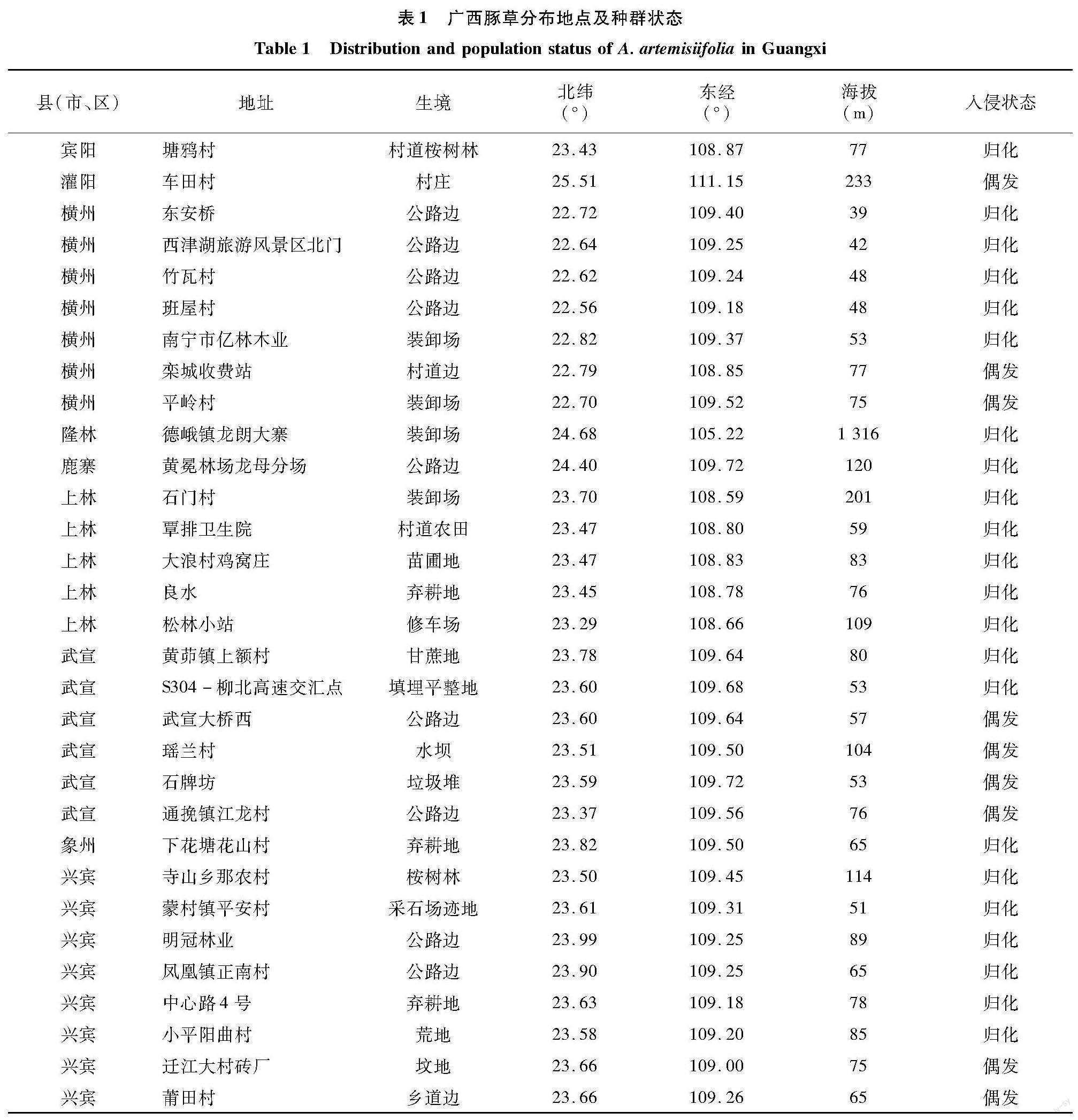
2.1? 廣西豚草分布地點及種群狀態
發現豚草的9個縣(市、區),共計82個樣帶,其中31個樣帶發現豚草,分布在興賓、上林、橫州、武宣、象州、鹿寨、賓陽、灌陽和隆林9個縣(市、區)(表1)。來賓市興賓區東南部仍為豚草危害重災區;蒙村鎮、寺山鄉尤為嚴重,部分地點豚草蓋度超過50%;周邊的武宣縣、象州縣均受影響;興賓區西部豚草分布較少。上林縣和橫州市形成了新的豚草暴發熱點;上林縣大浪村雞窩莊的一個黃花風鈴苗圃豚草生長極為繁茂,豚草蓋度達70%,密度約120株/m2;橫州市東安橋路邊豚草單優群落連片約100 m2,高達2.8 m。本研究發現的大多數豚草種群分布在桂中平原,海拔低于100 m,但在隆林海拔1 316 m的石山區公路邊臺地上(裝卸場)也發現歸化的豚草(表1)。
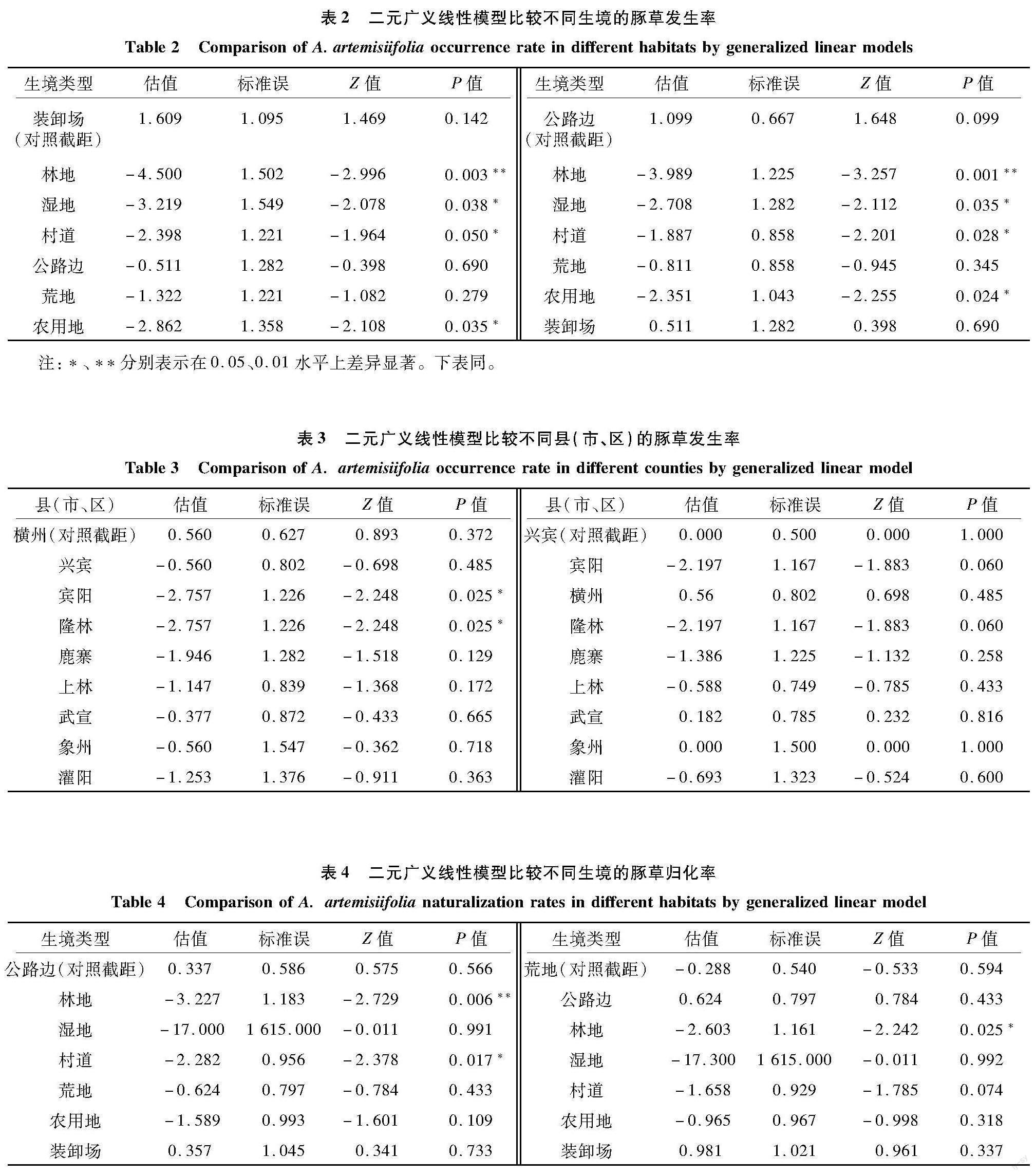
2.2? 不同生境及不同區域豚草發生率
9個縣(市、區)的82個樣地,不同生境的豚草發生率差異顯著,豚草發生率分別為裝卸場5/6,公路邊9/12,荒地8/14,村道5/16,農用地2/9,濕地1/6,林地1/19;裝卸場和公路邊的豚草發生率顯著或極顯著高于村道、農用地、濕地和林地,表2是以裝卸場和公路邊為對照時與其他生境豚草發生率比較結果。橫州的豚草發生率(7/11)顯著高于賓陽(1/10)和隆林(1/10);武宣(6/11)和興賓(8/16)的豚草發生率高于賓陽和隆林,但未達0.05顯著水平。表3是以橫州和興賓區為對照時不同縣(市、區)之間豚草發生率比較結果。
2.3? 不同生境豚草歸化率
調查結果表明,豚草歸化率分別為裝卸場4/6,公路邊7/12,荒地6/14,農用地2/9,村道2/16,林地1/19,濕地0/6,裝卸場歸化率最高,裝卸場和公路邊的豚草歸化率極顯著高于林地,荒地豚草歸化率顯著高于林地。表4是以公路邊和荒地為對照的不同生境豚草歸化率比較結果。象州和橫州樣地的豚草歸化率(1/2、5/11)略高于賓陽和隆林(1/10);其他縣(市、區)豚草歸化率之間無明顯差異。
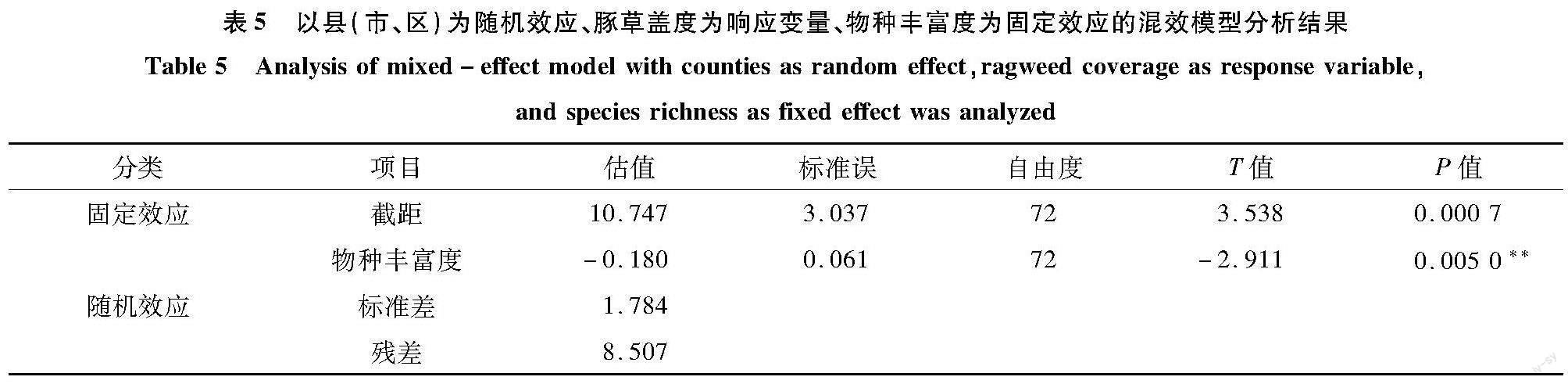
2.4? 物種豐富度固定效應的混效模型分析
物種豐富度與豚草蓋度呈負相關(r=-0.926);在響應變量為豚草蓋度、縣(市、區)為隨機效應、物種豐富度為固定效應的混效模型(表5)中,物種豐富度的斜率為-0.180,物種豐富度每增加10,豚草蓋度降低1.80%。但是,物種豐富度與豚草發生和豚草歸化變量均無顯著關聯。
3? 討論與結論
3.1? 豚草主要由人類活動傳播
豚草瘦果中央末端突喙可達2 mm[10],頂端有4~7個刺狀突起[19],附著力較弱[20];豚草種子較重,中國種群的平均質量為4.65 mg[11],只能靠重力傳播[20];大多數種子落在母株附近1 m之內[11];但豚草的土壤種子庫持久,被豚草種子污染的土壤可通過車輪實現遠距離遷移。在隆林,豚草發生在偏僻的桂西石漠化山區,海拔很高,且遠離來賓豚草高發地區;附近的石山是一個造林示范點。豚草種子和幼苗很容易隨樹苗一起傳播[15],林木苗圃很可能是隆林豚草的傳播源。
裝卸場是豚草發生率和歸化率最高的生境。裝卸場是貨物暫時堆放、受各種運輸工具頻繁干擾的場所,為豚草種子隨車輛遠距離傳播創造了機會;橫州和上林的豚草發生區大都與河沙、泥土、磚瓦等建材和農林收獲物的運輸有關。有研究認為,豚草傳播的主要途徑是車輛,輪胎、篷布等攜帶種子沿交通線路傳播[8,20]。
來賓市興賓區是廣西最早報道豚草入侵的地區,20世紀80年代該地區從新西蘭引進牧草種子時帶入[12];目前,豚草在興賓區的多個鄉鎮歸化,最大種群出現在興賓區東南部的寺山鄉和蒙村鎮。在興賓區城區及周邊5 km內的道路綠化帶、公園等多個地點均未發現豚草。
頻繁的人類活動干擾和當地豚草管理觀念淡薄共同導致橫州和上林成為新的豚草暴發熱點。橫州的豚草發生率(7/11)和歸化率(5/11)最高,廣西形成了橫州、興賓、上林三大豚草傳播中心。豚草新熱點比舊熱點的傳播風險更大,因為民眾往往對新熱點缺乏防范意識。賓陽縣北邊毗鄰興賓區,西邊緊連上林縣;靠近興賓區的賓陽農戶多知曉鄰鄉有豚草,須嚴防傳入。本研究在賓陽的10個調查樣帶中有9個沒有豚草,只在賓陽塘鴉村Y035鄉道旁發現豚草,該鄉道直通上林大浪村雞窩莊;賓陽成功地阻擋了興賓區方向的傳入,但豚草已從上林悄然入境。
3.2? 山嶺的物理阻隔作用明顯
大多數豚草種群分布在桂中平原,即駕橋嶺—大瑤山—蓮花山以西、都陽山—大明山以東的地區。在緊鄰武宣縣、興賓區的覃塘區、港北區和桂平市,均未發現豚草。蓮花山突兀橫亙在武宣與港北區之間,對豚草傳播起到物理阻隔作用。筆者在大明山西坡的馬山縣也進行了密集調查,未發現豚草。大明山東坡是新增豚草肆虐地區上林縣。崇山高嶺往往意味著經濟活動和人為干擾較少,限制了豚草隨人類活動的擴散。桂北和桂西除灌陽和隆林有零星短暫分布外,其他地方未發現豚草。
3.3? 氣候限制豚草向桂南沿海地區擴散
廣西南部沿海地區北海、欽州、防城港均沒有豚草發生,這種情況可能與氣候密切相關。豚草種子需要長時間的寒冷春化才能打破休眠。4 ℃是豚草最佳春化溫度[21]。-5 ℃春化效果最差,10 ℃處理效果中等,-5 ℃、10 ℃春化種子連續春化15周才能達到最大萌發率,而4 ℃春化種子12周即達到最高萌發率[22]。豚草種子萌發率隨著春化天數呈線性增加,4 ℃春化7、14、21、28 d后 25 ℃ 溫育催芽的萌發率約為14%、21%、29%、38%[23]。從天氣網(www.tianqi.com)查詢2012—2022年間冬季3個月的平均低溫,北海、欽州和防城港的11年冬季低溫均值高于12 ℃,來賓、上林、橫州的11年冬季低溫均值分別為9.2、9.6、10.2 ℃。沿海地區冬季氣候溫和,不利于豚草種子打破休眠,限制了豚草在沿海地區的擴展。
目前,筆者發現豚草入侵了廣西桂中平原和橫縣平原的9個縣(市、區),形成了橫州、興賓和上林3個暴發熱點。鑒于豚草的惡性生態影響和花粉過敏癥的危害,在向當地農林部門及時反饋的同時,應加大科普宣傳,讓當地人們了解該入侵物種的危害性,群防群治,以有效控制其在廣西的繼續蔓延。
參考文獻:
[1]Montagnani C,Gentili R,Smith M,et al. The worldwide spread,success,and impact of ragweed (Ambrosia spp.)[J]. Critical Reviews in Plant Sciences,2017,36:139-178.
[2]Schindler S,Bayliss H R,Essl F,et al. Effectiveness of management interventions for control of invasive common ragweed Ambrosia artemisiifolia:a systematic review protocol [J]. Environmental Evidence,2016,5:11.
[3]Basky Z,Ladányi M,Simonc[DD(-1*2/3]ˇic[DD(-1*2/3]ˇ A. Efficient reduction of biomass,seed and season long pollen production of common ragweed (Ambrosia artemisiifolia L.)[J]. Urban Forestry and Urban Greening,2017,24:134-140.
[4]梁巧玲,陸? 平. 新疆伊犁河谷發現外來雜草——三裂葉豚草和豚草[J]. 雜草科學,2014,32(2):38-40.
[5]謝云珍,王玉兵,譚偉福. 廣西外來入侵植物[J]. 熱帶亞熱帶植物學報,2007,15(2):160-167.
[6 ]周? 偉,徐瑞晶,趙? 倩,等. 廣州市花都區豚草種群監測調查[J]. 雜草科學,2010,28(3):9-13.
[7]鄧貞貞,趙相健,趙彩云,等. 繁殖體壓力對豚草(Ambrosia artemisiifolia)定殖和種群維持的影響[J]. 生態學雜志,2016,35(6):1511-1515.
[8]Galzina N,Baric[DD(-1*2/3]' K,c[DD(-1*2/3]'epanovic[DD(-1*2/3]' M,et al. Distribution of invasive weed Ambrosia artemisiifolia L. in Croatia[J]. Agriculturae Conspectus Scientificus,2010,75(2):75-81.
[9]Essl F,Dullinger S,Kleinbauer I.Changes in the spatio-temporal patterns and habitat preferences of Ambrosia artemisiifolia during its invasion of Austria[J]. Preslia,2009,81(2):119-133.
[10]Bassett I J,Crompton C W.The biology of Canadian weeds.11. Ambrosia artemisiifolia L. and A. psilostachya DC.[J]. Canadian Journal of Plant Science,1975,55(2):463-476.
[11]Essl F,Biro K,Brandes D,et al. Biological flora of the British Isles:Ambrosia artemisiifolia[J]. Journal of Ecology,2015,103(4):1069-1098.
[12]周忠實,陳紅松,鄭興汶,等. 廣聚螢葉甲和豚草卷蛾對廣西來賓豚草的聯合控制作用[J]. 生物安全學報,2011,20(4):267-269.
[13]劉曉亮,何金富,李克敵,等. 豚草卷蛾在廣西來賓建群狀況及其對作物的安全性調查[J]. 生物安全學報,2013,22(2):136-139.
[14]林春華,唐賽春,韋春強,等. 廣西來賓市外來入侵植物的調查研究[J]. 雜草科學,2015,33(1):38-44.
[15]Skálová H,Guo W Y,Wild J,et al. Ambrosia artemisiifolia in the Czech Republic:history of invasion,current distribution and prediction of future spread[J]. Preslia,2017,89(1):1-16.
[16]萬方浩,劉全儒,謝? 明,等. 生物入侵:中國外來入侵植物圖鑒[M]. 北京:科學出版社,2012.
[17]中國科學院中國植物志編輯委員會. 中國植物志[M]. 北京:科學出版社,1993.
[18]Pinke G,Kolejanisz T,Vér A,et al. Drivers of Ambrosia artemisiifolia abundance in arable fields along the Austrian-Hungarian border[J]. Preslia,2019,91(4):369-389.
[19]周業盛,吳永健,超材踞,等. 豚草的擴展規律和綜防技術的研究[J]. 雜草科學,1992(4):35-38.[HJ2.3mm]
[20]Vitalos M,Karrer G. Dispersal of Ambrosia artemisiifolia seeds along roads:the contribution of traffic and mowing machines [C]//Pyek P,Pergl J. Biological invasions:towards a synthesis. NeoBiota,2009,8:53-60.
[21]Baskin J M,Baskin C C.Ecophysiology of secondary dormancy in seeds of Ambrosia artemisiifolia[J]. Ecology,1980,61(3):475-480.
[22]Willemsen R W. Effect of stratification temperature and germination temperature on germination and the induction of secondary dormancy in common ragweed seeds[J]. American Journal of Botany,1975,62(1):1-5.
[23]Dinelli G,Marotti I,Catizone P,et al. Germination ecology of Ambrosia artemisiifolia L. and Ambrosia trifida L. biotypes suspected of glyphosate resistance[J]. Central European Journal of Biology,2013,8(3):286-296.
