通過CD20單抗打破小鼠HBV免疫耐受研究Bregs細胞在慢性乙型肝炎中的作用
2023-11-03 10:47:36劉中天張嘉靜林濤發舒丹王少揚
肝臟 2023年10期
關鍵詞:小鼠
劉中天 張嘉靜 林濤發 舒丹 王少揚
在感染、炎癥和自身免疫性疾病中B細胞可以通過呈遞抗原和分泌細胞因子來調節免疫反應,即便抗原低濃度下仍能精準識別并呈遞[1]。HBcAg特異性B細胞能在慢性乙型肝炎(CHB)患者體內持續產生抗-HBc,而抗-HBc的存在又能破壞肝臟細胞活性和影響HBV的清除,說明同樣是特異性B細胞產生抗體,卻能出現不同的結果[2]。接種疫苗的人群可在體內產生保護性抗體,抗體能阻斷HBV的感染和復制,推斷當B細胞功能障礙會引起體內HBsAg的持續存在和復制。臨床治療中發現使用針對B細胞增生為主的利妥昔單克隆抗體治療淋巴瘤時,如果不同時采取抗病毒措施將會引起HBV再激活甚至引起暴發性肝炎,極易進展為肝功能衰竭[3]。
調節性B細胞(regulatory B cell,Bregs)可以通過產生IL-10細胞因子,負性調控其他免疫細胞同時對效應B細胞也產生抑制作用,能下調炎癥級聯反應。Bregs細胞主要產生IL-10、IL-35和轉化生長因子-β(TGF-β),被認為是先于Tregs細胞發揮免疫作用的細胞[4]。Bregs細胞在系統性紅斑狼瘡、類風濕性關節炎、多發性硬化癥、艾滋病和丙型肝炎中的作用已有較多研究,但對乙型肝炎的作用鮮見報道。本研究探討Bregs細胞在CHB發病機制中的作用。
材料與方法
一、實驗材料
實驗動物為6~8周雄性C57BL/6小鼠20只,飼養于聯勤保障部隊第900醫院動物實驗中心,符合清潔級飼養條件。入組前均排除相關病毒感染、艾滋病、先天性免疫缺陷障礙等。研究方案經過聯勤保障部隊第900醫院醫學倫理委員會批準。
目前已知小鼠和人類都高表達CD1d+和CD19,在另外一種超敏模型中發現表面標志物為CD1d+CD5的B細胞能產生IL-10,抑制超敏反應過程[5, 6]。多種標志物組合檢測能提高檢測成功率,因此擬用anti-mouse CD1d+、CD5、CD19標志物作為檢測指標。
二、實驗方法
將p AAV/HBV 1.2質粒溶于無菌生理鹽水中,通過尾靜脈注射含質粒的鹽水,注射體積為小鼠質量的8%~10%,構建小鼠HBV感染模型。
將20只小鼠隨機分為實驗組和對照組各10只。實驗組小鼠尾靜脈注射CD20利妥昔單抗(Rituximab),對照組小鼠給予同劑量鹽水。比較兩組小鼠的肝功能、凝血功能、HBV DNA、Bregs細胞和IL-10。
Bregs細胞及IL-10分子水平檢測:分離脾臟、外周血、肝臟淋巴細胞,按說明書加入 anti-mouse CD1d+、CD5、CD19進行細胞表面及胞內因子染色,流式細胞儀檢測2組Bregs細胞比例。以前向散射角和側向散射角為坐標設門識別肝臟、脾臟以及外周血淋巴細胞,對CD1d+細胞進行設門,分析CD5+和CD19+Bregs細胞在臟器的分布比例。
三、統計學分析
采用SPSS 26.0 軟件進行統計學分析。偏態分布的計量資料以M(P25,P75)表示,比較采用Mann-WhitneyU檢驗,雙變量相關分析采用Spearman相關系數檢驗。P<0.05為差異有統計學意義。
結 果
一、實驗組與對照組小鼠血清學比較
實驗組和對照組小鼠HBV DNA比較差異無統計學意義(P>0.05)。實驗組小鼠的ALT和AST均高于對照組,而血清IL-10水平低于對照組,差異有統計學意義(P<0.05)。見表1。打破免疫耐受后,實驗組小鼠體內IL-10水平和ALT、AST水平呈負相關(r=-0.66、-0.65,均P<0.05)。

表1 實驗組與對照組ALT、AST以及IL-10水平比較[M(P25, P75)]
二、實驗組與對照組小鼠凝血功能比較
實驗組凝血酶原時間(PT)低于對照組,而纖維蛋白原(FIB)水平高于對照組,差異有統計學意義(P<0.05);活化部分凝血酶原時間、凝血酶時間差異無統計學意義(均P>0.05),見表2。
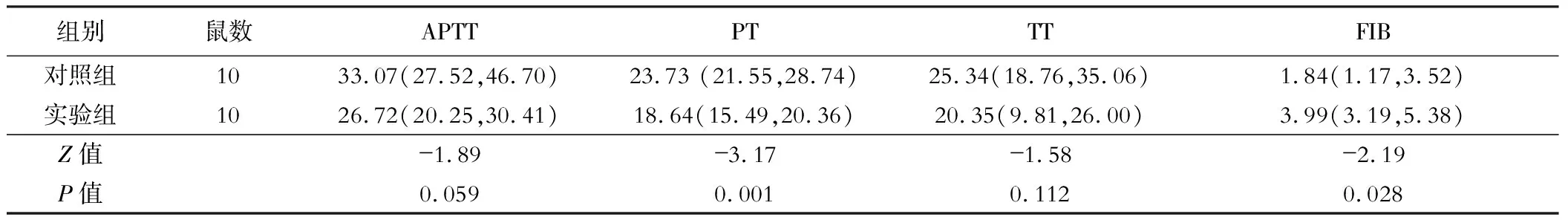
表2 兩組小鼠凝血功能比較[M(P25, P75)]
三、實驗組與對照組小鼠Bregs細胞在不同器官中的分布
實驗組肝臟脾臟、外周血Bregs細胞比例均低于對照組,差異有統計學意義(P<0.05);見表3。實驗組肝臟、脾臟、外周血Bregs細胞和IL-10因子行相關性分析,發現Bregs細胞與IL-10呈正相關(r=0.73、0.74、0.71,均P<0.05)。

表3 實驗組與對照組小鼠Bregs細胞在不同器官中的比例[%,M(P25,P75)]
討 論
本研究發現,隨著小鼠HBV感染免疫耐受的破壞,在第7天時出現了肝功能破壞,實驗組小鼠肝臟、脾臟、外周血Bregs細胞與對照組相比有明顯的下降,與IL-10存在明顯相關性,隨著Bregs細胞的破壞,IL-10的水平也降低。Bregs細胞的功能和數量的下降,伴隨著肝臟炎癥活動的增加。
通過研究打破HBV感染免疫耐受小鼠模型,充分證明在慢性乙型肝炎免疫狀態的激活過程中存在Bregs細胞的數量以及功能的變化,提示Bregs細胞對慢性乙型肝炎的炎癥活動和控制以及病情的進展有主要作用。而在打破免疫耐受過程中沒有發現HBV DNA的變化,提示乙型肝炎通過免疫途徑清除病毒是多方面的,可能包括T細胞、B細胞、NK細胞的參與,以及與γ-IFN、IL-2等多個細胞因子網絡控制。
Bregs細胞在自身免疫性疾病、感染性疾病、預防腫瘤相關生長、抑制免疫移植耐受中能通過分泌細胞因子抑制體內過度的炎癥反應,對Tregs細胞、CD4+、CD8+T細胞、DC細胞水平以及Th1/Th2的比例產生影響,且Bregs細胞數量的增減或功能強弱的變化都能導致疾病向不同的方向發展,尤其體現在后期的病情恢復的快慢,但Bregs細胞的表面標志物、參與激活的信號通路、細胞的分化狀態和不同的作用機制在HBV感染中并不明確[7]。
Bregs細胞在體內的變化能影響炎癥,可能與體內細菌和(或)病毒的負荷有關,將Bregs細胞過繼轉移到重癥免疫缺陷的炎癥性腸病的小鼠模型中后,發現能減輕炎癥程度[8]。Liu等[9]建立的自身免疫性肝炎的小鼠模型發現缺乏Bregs細胞后增加了EAH嚴重性。Gutiérrez等[10]在對HIV患者的研究中發現,未接受治療的新發患者Bregs(CD24hiCD38hi)或CD19+Tim-1+與Tregs呈正相關,而CD19+CD24hiCD27+或CD19+PD-L1+等Bregs亞群并沒有表現出相關性,可能是由于患者之間免疫系統的不同或對Bregs細胞表型的影響。
Bregs細胞通過多種途徑來發揮免疫抑制功能,除了分泌IL-10,還有IL-35和TGF-β等細胞因子,或者能通過細胞間的接觸傳遞信號,調節Tregs細胞的分化,降低炎癥因子的釋放和減弱致病原T細胞的反應[11]。不同組織來源的Bregs細胞收到誘導的信號不同,如在淋巴細胞中的信號是促炎細胞因子或已被激活的CD4+T細胞,而有研究指出脂肪趨化因子和代謝組織中的特異性信號,如CXCL12和游離脂肪酸在脂肪細胞中的重要作用之一就是能促進Bregs的分泌功能[12, 13]。
本實驗發現IL-10的水平與Bregs細胞的數量和功能存在明顯的相關性,提示在乙型肝炎的免疫發病機制中,Bregs細胞通過分泌IL-10可能是發揮功能的重要途徑。近年來對Bregs細胞分泌IL-35以及Bregs細胞是否和其他免疫細胞通過細胞間的接觸而發揮作用,還需要進一步觀察。
利益沖突聲明:所有作者均聲明不存在利益沖突。
猜你喜歡
興趣閱讀·興趣作文與閱讀(低年級)(2025年8期)2025-08-18 00:00:00
東方少年·布老虎畫刊(2023年8期)2023-08-01 15:45:12
科學大眾(2021年6期)2021-07-20 07:42:44
科學(2020年3期)2020-11-26 08:18:30
學苑創造·A版(2020年9期)2020-10-13 09:41:02
娃娃樂園·綜合智能(2019年3期)2019-04-03 09:17:36
中成藥(2018年2期)2018-05-09 07:19:34
小學生學習指導(低年級)(2017年10期)2017-10-10 01:00:05
湖北師范大學學報(自然科學版)(2015年2期)2016-01-10 08:41:55
云南中醫學院學報(2014年3期)2014-07-31 18:57:34