枯草芽孢桿菌BBs-27發酵液性質及脂肽對黃色鐮刀菌的抑菌作用
2023-10-25 05:05:42苗永美苗翠蘋于慶才
生物技術通報 2023年9期
苗永美 苗翠蘋 于慶才
(1.安徽科技學院生命與健康科學學院,鳳陽 233100;2.云南大學生命科學學院,昆明 650504)
鐮刀菌(Fusarium)是一種世界性真菌,已鑒定的有300多種,常以物種復合體存在,其中23種復合體已被研究,近年來,該病原菌因在世界范圍內侵染大量農作物造成巨大經濟損失而備受關注[1]。如棉花枯萎病和香蕉枯萎病均由尖孢鐮刀菌引起[2-3],小麥赤霉病由禾谷鐮刀菌復合種引起[4-5]。鐮刀菌不僅侵染植物,還有70多個種使人類致病,如鐮刀菌性角膜炎菌對免疫力缺陷患者會造成較為嚴重的感染[1]。此外,鐮刀菌代謝毒素如鐮刀酸、伏馬菌素和白僵菌素會污染食品,引起人畜食用急性中毒[6]。鐮刀菌具有多宿主性和強侵染力,通過侵染寄主植物維管束系統破壞輸導組織,引起植物器官腐爛、萎蔫等,導致作物產量損失和品質下降。鐮刀菌通過形成厚垣孢子及與寄主、輪作物、雜草等形成寄生和致病關系,持續存于生產系統中[2],診斷和控制具有挑戰性,尤其是土傳病菌尖孢鐮刀菌的防治。生物防治具有高效、安全、環保等優勢,符合現代農業持續發展趨勢。
脂肽是一種具有獨特兩親性的微生物次級代謝產物,一部分是由多個氨基酸組成的肽鏈,為親水基,另一部分是具有β-羥基或氨基的脂肪酸鏈,為親油基[7]。脂肽因其低耐藥性而被認為是抗生素安全替代品從而受到廣泛關注,具有抗真菌、細菌、病毒、腫瘤和殺蟲等功能。近期研究表明,絕大多數芽孢桿菌菌株都可以產生LPs,如枯草芽孢桿菌、解淀粉芽孢桿菌、蘇云金芽孢桿菌和蠟樣芽孢桿菌。抗菌LPs根據結構分為Iturins、Surfactins 和Fengycins三大家族,其氨基酸殘基序列、肽的環化以及脂肪酸鏈的長度和分支決定其性質和生物活性[8]。不同芽孢桿菌產生的脂肽類型不同,其表面活性劑特性和抑菌活性也存在差異。目前關于脂肽抑菌機制研究大多是從形態上通過電鏡掃描觀察細胞壁和細胞膜損傷、產生質膜離子通道導致離子滲透、ROS導致細胞壁破壞等幾個層面。Li等[9]通過轉錄組分析脂肽中Fengycin和Iturins對黃曲霉的抑制機制發現,主要是通過下調核糖體發生途徑和黃曲霉毒素合成途徑中部分基因。
病原菌MF01(分子鑒定與黃色鐮刀菌F.culmorum相似性高),為實驗室前期從發病棉花植株上分離,黃色鐮刀菌是重要的谷物冠腐病菌,易導致食品污染[10]。作者前期開展石豆蘭(Bulbophyllum sp.)離體培養時發現內生菌豐富,通過平板對峙和發酵液抑菌兩級方法,篩選出一株細菌對MF01有較好抑制效果,分子鑒定與Bacillus subtilis subsp.subtilis strain 168相似性為98.93%,命名BBs-27[11]。為進一步掌握發酵液中抑菌成分及抑菌機制,對發酵液理化性質進行初步分析發現,主要抑菌成分為脂肽(lipopeptides, LPs)。然而,BBs-27分泌的脂肽類型及其對F.culmorum抑菌機制尚不明確。本研究擬通過LC-MS分析抑制F.culmorum的脂肽類型,并從形態、生理和轉錄組3個方面分析其抑菌機理,旨在為鐮刀屬病原菌防治提供新思路,為開發新型、安全殺菌劑提供應用基礎。
1 材料與方法
1.1 材料
枯草芽孢桿菌BBs-27為前期從石豆蘭植物中分離[11];病原菌MF01從發病棉花植株上切取莖段、經表面消毒,接種PDA培養基上,28℃培養,經過多次分離、純化獲得。
1.2 方法
1.2.1 MF01分子鑒定 擴增18S RNA、28S RNA、tef-1α三個基因片段。引物(NS1: 5′-GTAGTCATATGCTTGTCTC-3′, NS4: 5′-CTTCCGTCAATTCCTTTAAG-3′)擴增18S RNA,引物(NL1: 5′-GCATATCAATAAGCGGAGGAAAAG-3′, NL4: 5′-GGTCCGTGTTTCAAGACGG-3′)擴增28S RNA,引物(EF1: 5′-ATGGGTAAGGA(A/G)GACAAGAC-3′, EF2: 5′-GGA(G/A)GTAC CAGT(G/C)ATCATG-3′)擴增tef-1α[12]。DNA提取、引物合成、擴增、序列拼接等工作委托通用生物(安徽)股份有限公司完成。根據各基因序列系統發育分析結果,從NCBI/GenBank數據庫中搜索并下載相關菌株的各基因序列,Sequencher軟件將下載的原始基因序列串聯拼接,用Clustal X軟件將拼接后的基因序列Alignment比對。用MEGA 7.0軟件,選擇Kimura 2-parameter+Gamma Distributed模型計算進化距離,采用最大似然法(maximum-likelihood)構建系統發育樹,Bootstrap method 1 000次評估系統發育樹各分支置信度。
1.2.2 BBs-27發酵上清液制備 參照苗永美等[13]方法依次進行菌種活化、種子液制備、發酵、收集上清、微孔膜過濾。
1.2.3 發酵液理化性質分析 1.2.2過濾液用4% HCl(V/V)和4% NaOH(m/V)將pH分別調至2、4、6、8、10、12,放置5 h,再調回至初始pH;取10 mL過濾液于直徑9 cm培養皿中,紫外照射5、10、15、20、25、30 min;取9 mL過濾液分別加入1 mL 10 mg/mL的胰蛋白酶、胰凝乳蛋白酶、胃蛋白酶、蛋白酶K,37℃放置2 h;不進行任何處理為對照組(pH 5.5)。各處理液:PDA(1∶10)混合制平板,選用瓊脂柱法進行抑菌試驗,計算抑菌率,具體方法參照苗永美等[13]方法。
1.2.4 粗脂肽提取 根據1.2.3結果,推斷發酵液中主要抑菌物質為脂肽。按照Wu等[14]方法略作修改。具體方法:7 mol/L HCl調至pH 2,過夜、收集沉淀,少量蒸餾水溶解,1 mol/L NaOH調至中性,甲醇多次抽提、40℃減壓濃縮、35℃真空干燥得粗脂肽干品。配制10 mg/mL母液。
1.2.5 粗脂肽的純化與LC-MS鑒定 甲醇溶解粗脂肽,拌硅膠、陰涼處自然風干。濕法裝柱、氯仿壓柱、上樣,用氯仿洗脫,依次用氯仿∶甲醇體積比為98∶2、95∶5、9∶1、8∶2進行梯度洗脫,牛津杯法分析各組分抑菌活性。具抑菌活性的組分采用超高效液相色譜-線性離子阱靜電場軌道阱高分辨質譜儀(型號:Ulitimate 3000- LTQ Orbitrap XL)進行分析。色譜條件:C18柱(Hypersil GOLD,100 mm×2.1 mm),填料粒徑1.9 μm,柱溫27.5℃。流動相:甲醇∶水(0.1%含甲酸)=98∶2,流速0.3 mL/min。質譜條件:HESI離子源,離子源溫度300℃,噴霧電壓3.5 kV,高分辨FTMS模式,分辨率60 000,采集正離子,質核比范圍100-1 200。
1.2.6 電鏡掃描 PDA 培養基中添加500 μg/L脂肽后接種病原菌,培養3 d,挑取菌絲電鏡掃描,具體步驟參見苗永美等[13]方法。
1.2.7 生理指標測定 根據1.2.4試驗結果,PDA中添加脂肽使終濃度分別為100、500、1 000 μg/L,蒸餾水為對照。培養3 d時,刮取菌絲于預冷研缽中,加pH 7.8、0.05 mol/L預冷PBS研磨成勻漿,4℃、12 000 r/min離心20 min,上清為待測酶液。超氧化物歧化酶(SOD)活性采用氮藍四唑法測定,以抑制NBT光化還原的50%為一個酶活單位(U);過氧化物酶(POD)活性采用愈創木酚法測定,以每分鐘OD值變化(升高)0.1為1個酶活性單位(U);過氧化氫酶(CAT)活性采用過氧化氫還原法測定,以每分鐘OD值變化0.1為1個酶活性單位(U);可溶性蛋白質采用考馬斯亮藍法測定;可溶性糖含量采用苯酚硫酸法測定。
1.2.8 轉錄組分析 PDA+500 μg/L脂肽,鋪玻璃紙、接種、28℃培養3 d,刮取菌絲液氮速凍后送至美吉生物公司,3次生物學重復(T1-T3),蒸餾水為對照(C1-C3)。依次提取總RNA、Oligo dt富集mRNA、片段化,反轉錄、End Repair Mix將雙鏈cDNA補成平末端,加ployA接頭、擴增、Illumina平臺測序。對原始數據過濾和質控,使用比對工具Hisat2將高質量序列與參照物種禾谷鐮刀菌(F.graminearum)基因組進行mapping,統計每個樣品比對到每個基因上reads數;RSEM軟件以TPM>1為衡量標準計算每個基因表達量;DESeq2軟件分析差異表達基因DEGs,以P-adjust<0.05和|log2FC|≥1為DEGs的篩選條件;用GO數據庫進行基因功能注釋;以顯著性水平P-value≤0.5對功能注釋的基因進行KEGG富集,確定DEGs主要富集的通路及其可能的生物學功能。
1.2.9 數據統計 采用Excel軟件進行數據處理、SPSS19.0軟件進行差異顯著性分析,Origin2018作圖。轉錄組分析及作圖均在美吉生物公司網站完成。
2 結果
2.1 病原菌MF01分子鑒定
18S RNA、28S RNA、tef-1α三個基因片段串聯拼接、比對構建系統進化樹,結果(圖1)表明,MF01與F.culmorum、F.cerealis聚成一個支持強度為78%的末端分支。根據相似性高低,將MF01鑒定為黃色鐮刀菌(F.culmorum)。

圖1 基于三基因序列拼接的MF01系統發育樹Fig.1 Phylogenetic tree of MF01 based on three-gene sequence splicing
2.2 抑菌物質穩定性分析
發酵液經酸堿處理后抑菌活性有明顯變化,pH 6-8時抑菌活性與對照(pH 5.5)差異不顯著,說明抑菌成分能耐一定酸堿,當pH < 4或>10時,活性會顯著降低(圖2-A)。短時間紫外線照射不會影響抑菌成分活性,超過10 min會明顯降低活性(圖2-B)。抑菌物質對胃蛋白酶不敏感,但經胰蛋白酶、胰凝乳蛋白酶和蛋白酶K處理后其抑菌率顯著降低(圖2-C)。

圖2 不同處理后發酵液的抑菌效果Fig.2 Inhibitory effects of fermentation broths after different treatments
2.3 脂肽成分鑒定
硅膠柱共分離到24種組分,其中組分23具有抑菌活性。對組分23進行UPLC-MS分析,0.94 min左右出現一個主要洗脫峰(圖3-A),收集洗脫物進行HESI-MS,結果出現了兩大類物質。第一大類主含物質其m/z值為1 065.54、1 079.55、1 093.57(圖3-B),它們的相對分子量相差14 Da,與一個亞甲基-CH2的分子量大小一致,說明是相差一個脂肪鏈的同系物,對比脂肽類物質的相對分子質量推斷出為伊枯草菌素家族類物質。第二大類主含物質其m/z值為1 433.80、1 447.82、1 461.83、1 463.81、1 477.83、1 491.84、1 505.85、1 519.86、1 533.88,1 485.78、1 499.80、1 513.82(圖3-C),以上兩組物質相對分子量也相差14 Da,根據分子量推測為豐原素同系物。

圖3 脂肽的LC-MS分析Fig.3 LC-MS analysis of LPs
2.4 電鏡掃描結果
電鏡掃描結果顯示,對照組菌絲表面光滑,粗細較為一致,生長正常(圖4-A),脂肽處理組菌絲扭曲、不規則、粗細不均,異常擴張,部分部位膨大為“氣球狀”(圖4-B)。由此看出,脂肽對黃色鐮刀菌的傷害在菌絲形態上體現較為明顯。
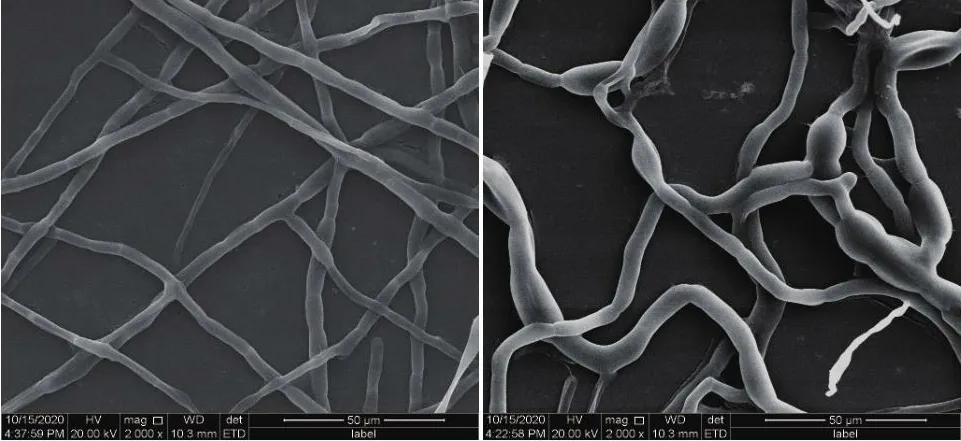
圖4 掃描電鏡下黃色鐮刀菌菌絲形態特征Fig.4 Mycelium morphology of F.culmorum by SEM
2.5 脂肽對黃色鐮刀菌保護酶活性影響
由圖5-A看出,100 μg/L脂肽處理菌絲體內SOD活性與對照差異不顯著,當濃度大于500 μg/L時,SOD活性顯著低于對照,兩個處理濃度下分別降低了49.78%和53.66%,說明此時已對SOD系統造成傷害。3個脂肽濃度會顯著降低菌體POD和CAT活性,POD比對照分別降低了41.05%、62.57%、63.68%(圖5-B),CAT活性比對照分別降低了30.66%、58.64%、74.70%(圖5-C),說明100 μg/L的脂肽濃度就會對POD和CAT系統造成傷害。3種保護酶系統受脂肽影響最大的是POD,其次是CAT,最小的是SOD。

圖5 脂肽對保護酶的影響Fig.5 Effects of lipopeptide on protective enzymes
2.6 脂肽對黃色鐮刀菌滲透調節物質影響
隨著脂肽濃度的提高,菌絲體內可溶性蛋白和可溶性糖含量逐漸降低,顯著低于對照。可溶性蛋白分別降低了25.32%、39.09%、63.44%,但100 μg/L和500 μg/L處理上含量差異不顯著(圖6-A)。3個脂肽處理下可溶性糖比對照分別降低了28.40%、43.85%、78.13%(圖6-B)。說明大于100 μg/L的脂肽已顯著影響菌體中兩種滲透調節物質的合成。
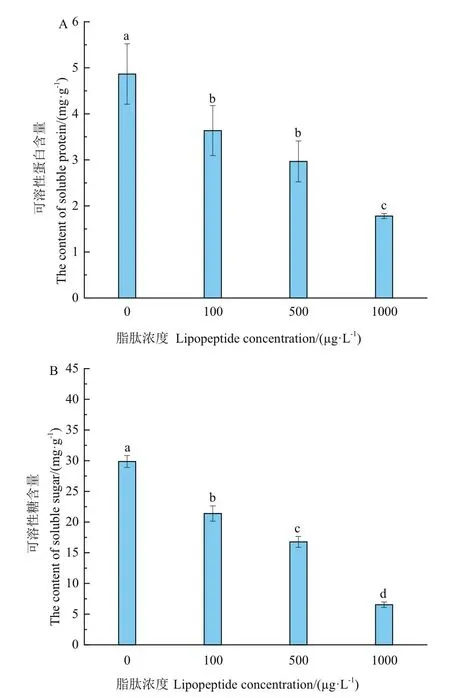
圖6 脂肽對滲透調節物質影響Fig.6 Effects of lipopeptide on osmoregulatory substances
2.7 指標相關性分析
抑菌率高說明菌絲生長緩慢,菌絲受到傷害大。相關性分析顯示(表1),抑菌率與5項指標呈極顯著負相關,說明菌絲體內3種保護酶、可溶性糖和可溶性蛋白受到脅迫后都作出強烈響應,或者說受到嚴重破壞。除SOD活性與其他4項指標、POD與可溶性糖呈顯著相關外,其他指標之間相關性均達到極顯著水平。每項指標與其他5項指標相關系數平均值從大到小依次為:抑菌率>CAT活性>可溶性蛋白>可溶性糖>POD活力>SOD活力。

表1 指標之間相關系數Table 1 Correlation coefficient among indexes
2.8 黃色鐮刀菌響應脂肽的轉錄組分析
2.8.1 轉錄組測序數據分析 T組和C組6個cDNA庫共產生316 765 242條reads,質控后獲得305 671 078條reads,各樣品測得數據過濾后所占比例超過96%,樣品誤差率< 0.03%,Q20>96%,Q30>91%,GC含量在52.27%-52.87%,比對率>78%,說明測得數據準確度較高,利于后期數據分析(表2)。

表2 質控數據統計結果Table 2 Quality control data statistics
2.8.2 基因表達及DEGs分析 利用TPM>1為衡量指標計算基因表達量,對照組中共鑒定出9 015個表達基因,特有表達基因187個;處理組中共有9 219個基因,特有表達基因391個;共同表達的基因有8 828個(圖7)。與對照相比,處理組中篩選出712個DEGs,其中上調表達基因393個,下調表達基因319個(圖8)。

圖7 不同樣品間表達量Venn分析Fig.7 Venn analysis of gene expressions between samples
2.8.3 DEGs的GO功能注釋分析 GO功能注釋分類結果表明,712個DEGs主要涉及到6個生物學過程、11個細胞組分和12分子功能,基因數量前20位GO分類見圖9。3個大類分別有300個(上調基因130個,下調基因170個)、453個(上調基因214個,下調基因239個)、570個(上調基因275個,下調基因295個)DEGs。根據富集DEGs數量,主要生物學過程有代謝進程(metabolic process)和細胞進程(cellular process),主要細胞組分有膜組分(membrane part)和細胞組分(cell part),主要分子功能有催化活性(catalytic activity)和結合功能(binding)。
2.8.4 DEGs的KEGG通路富集分析 對所有DEGs進行KEGG通路富集分析,結果表明,712個DEGs中有185個富集到78條代謝通路中,涉及到代謝、細胞進程、生物系統、人類疾病、環境信息處理、遺傳信息處理6個一級分類,其中89個上調DEGs參與41個代謝通路,96個下調DEGs參與60個代謝通路(圖10)。P-value<0.05顯著富集的有6條(表3)。

圖10 差異表達基因KEGG富集前20位代謝通路Fig.10 Top 20 metabolic pathways of differentially expressed genes enriched by KEGG
顯著富集到碳水化合物途徑共14個DEGs,參與半乳糖代謝(Map00052)、氨基糖和核苷糖代謝(Map00520),已知的有半乳糖氧化酶(galactose oxidase, GAO)前體、內切幾丁質酶(endochitinase,CHIT)前體,幾丁質合成酶3(chitin synthase 3,CHS3)、甘露糖-1-磷酸鳥苷酰轉移酶(mannose-1-phosphate guanyltransferase, MPG1)基因,其他均為未知假設蛋白(hypothetical protein)(表3)。脂肽處理會使菌絲體中GAO和MPG1上調表達,而CHS3和CHIT都下調表達。
DEGs顯著富集到脂代謝通路是麥角甾醇合成途徑(Map00100),有具體名稱的3個DEGs分別是Δ(14)-甾醇還原酶基因(Delta(14)-sterol reductase, ERG24)、C-5甾醇去飽和酶基因(C-5 sterol desaturase, ERG3)、C-4甲基甾醇氧化酶基因(C-4 methylsterol oxidase, ERG25)[15]。ERG24、ERG25、ERG3和FGSG_06215均上調,而FGSG_02016下調表達,說明所用脂肽處理濃度對細胞膜傷害較小,菌體通過積極上調部分麥角甾醇合成相關酶表達來調整細胞膜流動性以抵抗外界脅迫。
氨基酸代謝通路中有具體名稱的3個DEGs分別是3-異丙基蘋果酸脫水酶(3-isopropylmalate dehydratase, 3-IPDH)、酮酸還原酶(ketol-acid reductoisomerase, KARI)、鳥氨酸脫羧酶(ornithine decarboxylase, ODC),該4種酶參與支鏈氨基酸纈氨酸、亮氨酸和異亮氨酸的生物合成(Map00290)。IPDH呈下調、KARI呈上調變化,另兩種未知酶分別呈上調和下調變化,其變化可能會影響3種支鏈氨基酸合成,尤其是變化幅度較大的下調基因如IPDH可作為BBs-27脂肽靶標重點研究。脂肽處理組菌體中參與谷胱甘肽代謝的酶基因有3個上調、2個下調,下調表達且有具體功能描述的是ODC,該酶催化鳥氨酸形成的尸胺參與到谷胱甘肽代謝(Map00480),通過下調表達可能會影響生物體的谷胱甘肽代謝,從而影響抗氧化系統。
在輔助因子和維生素代謝(Map00770)中差異表達的DEGs有4個,包含1個下調和3個上調,具體名稱描述的是KARI,呈上調變化。
3 討論
3.1 脂肽的理化性質及類型
目前研究表明有些枯草芽孢桿菌菌株的代謝物質能抑制農業病原菌[16]、動物病原菌[17]或食品源致病菌[18]。不同菌株產生的活性物質不同,抑菌效果也不同,目前報道的活性成分多是蛋白質類[19-21]和脂肽類[3,7,22],也有揮發性物質研究的少量報道[23]。吳越等[19]研究表明,枯草芽孢桿菌HAINUP40分泌的能夠抑制羅非魚無乳鏈球菌的活性物質對蛋白酶K敏感,不耐高溫,pH < 4或> 8時失去抑菌活性,可以耐受短時間(< 180 s)紫外線照射,表明抑菌物質為大分子蛋白質,并非小分子脂肽。鄧陽等[20]研究枯草芽孢桿菌J-4菌株產生的抑菌物質在100℃水浴60 min、pH為2.8-8.0范圍內均具有較高活性,CTAB和CaCl2還能增強其活性,但會被胃蛋白酶或胰蛋白酶分解,據此推測為抗菌蛋白類物質。李麗等[21]也從枯草芽孢桿菌S6的發酵液中分離到抗棉花枯萎病的拮抗蛋白。關于枯草芽孢桿菌產脂肽類抑菌物質的研究更多,脂肽是非核糖體途徑合成,具有活性強、穩定性高、安全等特點。與抗菌蛋白質不同的是多數脂肽耐受性強,曾國洪等[22]研究表明,枯草芽孢桿菌產生的脂肽在25-80℃下抗菌活性幾乎保持不變,具良好熱和pH穩定性,對胰蛋白酶和胃蛋白酶不敏感。BBs-27發酵液理化性質分析結合前期耐熱性研究[13],推測抑菌物質是以脂肽類為主,且包括少量蛋白質。不同菌株產生脂肽類型不同,LC-MS分析證實BBS-27抑制F.culmorum的抗菌肽主要是伊枯草菌素和豐原素兩大類,每類也都是復合物。B.velezensis產生的脂肽主要是伊枯草菌素和豐原素[18],B.amyloliquefaciens能產生表面活性素、泛革素和伊枯草菌素三類脂肽類化合物以及Bacillibactin、Difficidin和Bacillaene三類聚酮類化合物[24]。
3.2 脂肽的抑菌機制
BBs-27分泌的兩類脂肽會導致菌絲畸形,細胞壁合成受阻,異常腫脹,部分呈“氣球狀”,與B.velezensis和B.amyloliquefaciens粗提脂肽分別對黃曲霉和香蕉枯萎菌絲細胞壁破壞結果非常相似[21,24],也能使曲霉細胞呈“氣球狀”[25]。BBs-27分泌的兩類脂肽使菌體中SOD、POD、CAT酶活性和可溶性蛋白、可溶性糖含量顯著下降,相關性分析表明,脂肽是通過破壞SOD、POD、CAT等保護酶系統,及抑制可溶性蛋白和可溶性糖合成來抑制菌體生長。
細胞壁是鐮刀菌侵染植物過程中最先接觸的結構,是鐮刀菌保持致病力和正常發育所必須結構。真菌細胞壁多糖包括幾丁質、葡聚糖、甘露聚糖和半乳糖等,GAO、CHIT、CHS3、MPG1參與細胞壁形成。GAO 催化半乳糖形成乙醛糖和H2O2,注射D-半乳糖致小鼠衰老的機理是ROS的增加引起機能損傷,干擾細胞正常代謝[26]。脂肽處理會引起F.culmorum中GAO上調表達,推測真菌可能也與動物體內一樣因產生過多ROS損傷機體,此外乳糖的氧化還會導致細胞壁結構遭到破壞。CHS3是參與合成禾谷鐮刀菌細胞壁的基因之一,是其致病和發育的關鍵基因[27],還影響環境適應能力[28]。值得注意的是,本研究中幾丁質合成基因CHS3和降解基因CHIT都下調表達,推測脂肽主要是通過抑制幾丁質合成來影響細胞壁結構,因此,CHS3可作為農藥靶標深入研究。此外,MPG1上調表達會加大催化甘露糖形成GDP-甘露糖的力度,造成細胞壁穩定性和強度減弱,也會使菌體變脆弱。
脂肽能增大Botrytis cinerea和F.oxysporum細胞膜通透性,導致滲透物質流出和電導率上升[6,29]。麥角甾醇是真菌細胞膜主要固醇類物質,本研究中參與麥角甾醇代謝通路的5個DEGs中有4個上調,說明脂肽對F.culmorum細胞膜會有一定影響,菌體能通過積極上調部分麥角甾醇合成相關酶表達來調整細胞膜流動性以抵抗傷害,而FGSG_02016表達下降,后期可對序列信息、調控功能、與細胞膜損傷相關性等進行深入研究。本研究也進一步證實了麥角固醇合成途徑的酶類是唑類抗真菌藥物的主要靶點[15]。
脂肽破壞3種酶系統的分子響應機制,可能是通過改變GAO和ODC等基因表達。方欣等[30]通過基因敲除和外源恢復等方法,證明3-異丙基蘋果酸脫水酶基因(FgLEU1)在禾谷鐮刀菌亮氨酸合成、菌絲孢子形成及產毒致病過程中發揮著重要作用。脂肽導致IPDH下調表達的結果可能會干擾支鏈氨基酸合成,進而影響菌體蛋白合成和生長。由于支鏈氨基酸合成途徑只在植物和微生物中存在,哺乳動物中不存在,從農產品安全角度出發,支鏈氨基酸合成酶成為安全藥劑的靶標和研究熱點。
4 結論
BBs-27產生抑制F.culmorum的物質主要是脂肽類且含有少量蛋白質,抗菌肽包含伊枯草菌素和豐原素兩大類。脂肽使菌絲卷曲、畸形、異常膨大,能破壞SOD、POD、CAT和可溶性蛋白及可溶性糖合成系統,生理變化與生長表現呈極顯著相關性。脂肽通過改變GAO、CHS3、MPG1的表達來干擾幾丁質、半乳糖和甘露糖和麥角甾醇合成以破壞細胞壁和細胞膜,導致細胞生長受阻;脂肽還能上調GAO而產生H2O2損傷機體,下調ODC干擾谷胱甘肽抗氧化系統,降低抗氧化酶活性。此外,脂肽還能通過下調IPDH表達干擾支鏈氨基酸代謝,導致細胞死亡。GAO、CHS3、MPG1、IPDH均可作為脂肽作用靶點用于殺菌劑開發。
