生防菌株DZY6715在不同生長期的代謝差異分析
2023-10-25 05:11:36周嬡婷彭睿琦王芳伍建榕馬煥成
生物技術(shù)通報(bào) 2023年9期
周嬡婷 彭睿琦 王芳 伍建榕,2 馬煥成
(1.西南林業(yè)大學(xué)林學(xué)院,昆明 650224;2.西南林業(yè)大學(xué)生物多樣性保護(hù)學(xué)院,昆明 650224)
特基拉芽孢桿菌(Bacillus tequilensis)作為芽孢桿菌屬(Bacillus)中具有較好拮抗作用的一類生防微生物,可以通過產(chǎn)生抗生素、細(xì)胞壁水解酶、揮發(fā)性物質(zhì)、次級(jí)代謝產(chǎn)物等來抑制靶標(biāo)病原菌或細(xì)菌的生長繁殖,促進(jìn)植物生長[1-3]。多方面的研究已經(jīng)得到證實(shí),Kiran等[4]發(fā)現(xiàn)從海洋中分離得到的B.tequilensis MSI45對耐藥性極強(qiáng)的金黃色葡萄球菌具有較強(qiáng)的抗菌及抗氧化活性。Li等[5]從白芷中分離到了一株對稻瘟病菌具有較高抑制活性的內(nèi)生菌B.tequilensis GYLH001,是能夠在農(nóng)業(yè)上用于防治稻瘟病的潛力生防治劑,這對于提升綠色糧食質(zhì)量具有重要作用。Zhou等[6]從健康水稻植株中獲得了一株能夠抑制多種植物病原菌發(fā)生發(fā)展的菌株B.tequilensis JN-369,除了具有較好的拮抗作用外,還能促進(jìn)水稻種子萌發(fā)和幼苗植株生長,提高植物防御酶活性,顯著減少水稻葉瘟病和穗瘟病,并且能夠提高田間水稻的產(chǎn)量,是一種潛在的稻瘟病生物防治劑和水稻栽培生物肥料。Shultana等[7]在沿海受鹽影響的水稻種植區(qū)收集并篩選到了潛在耐鹽植物生長促進(jìn)菌B.tequilensis UPMRB9,它能夠增強(qiáng)滲透保護(hù)特性,提高抗氧化酶活性,是一種潛在的生物增強(qiáng)劑。綜上,可見特基拉芽孢桿菌(B.tequilensis)在一定程度上可以減輕農(nóng)林業(yè)生產(chǎn)中化學(xué)農(nóng)藥的使用劑量,提高作物對脅迫因素的抵抗力,增加生物量等。
代謝組學(xué)是生物學(xué)科中的新興組學(xué),研究的是生命體內(nèi)的代謝產(chǎn)物種類、數(shù)量及其變化規(guī)律的科學(xué),能夠更直接、更準(zhǔn)確地反映生物體的生理狀態(tài),揭示其參與的生命活動(dòng)機(jī)制[8-9]。其中,非靶向分析模式能夠?qū)⑸矬w內(nèi)代謝水平的擾動(dòng)情況最大程度地反應(yīng)出來,側(cè)重于系統(tǒng)性和整體性的分析。作者發(fā)現(xiàn)B.tequilensis DZY6715能夠有效防控油茶炭疽菌,且該菌株在生長前期和生長后期對油茶炭疽菌的抑菌活性存在差異,尤以后期階段的抑制活性高,并且隨著培養(yǎng)時(shí)間的遞增,防效越明顯。所以,本文擬通過非靶代謝組的方法研究菌株DZY6715在生長前期和生長后期發(fā)揮主要抑菌活性的物質(zhì),探究這兩個(gè)時(shí)期差異代謝物的種類、調(diào)控機(jī)制及功能作用等,為生防菌株DZY6715的抑菌活性分子機(jī)制提供一定的參考價(jià)值。
1 材料與方法
1.1 材料
特基拉芽孢桿菌(B.tequilensis)DZY6715和油茶炭疽菌(Colletotrichum fructicola)是筆者分別從油茶健康葉片和油茶炭疽病葉片中分離獲得的,保存在西南林業(yè)大學(xué)生物多樣性保護(hù)學(xué)院病理學(xué)實(shí)驗(yàn)室。
LB培養(yǎng)基(g/L):蛋白胨10,酵母提取物5,NaCl 10,瓊脂20,pH自然,121℃滅菌20 min,冷卻至室溫,備用;PDA培養(yǎng)基(g/L):馬鈴薯200,葡萄糖20,瓊脂20,pH自然,121℃滅菌20 min,冷卻至室溫,備用。
1.2 方法
1.2.1 菌株DZY6715生長曲線測定 首先將菌株DZY6715在LB固體培養(yǎng)基上培養(yǎng)24 h,然后用無菌接種針把單菌落移至200 mL的LB液體培養(yǎng)基中培養(yǎng),分別取(t=6、12、24、48、72、96 h)的菌液稀釋后涂布,28℃培養(yǎng)箱培養(yǎng)24 h后計(jì)數(shù),根據(jù)稀釋倍數(shù)計(jì)算每毫升菌液中的活菌數(shù)量[10]。
1.2.2 不同時(shí)期培養(yǎng)液對油茶炭疽菌的抑制活性 采用平板對峙法[11]測定菌株DZY6715在生長前期(t=24 h)和生長后期(t=72 h)兩個(gè)階段對油茶炭疽菌的抑菌活性,抑菌率的計(jì)算公式為:抑菌率(%)=[(對照組菌落直徑-處理組菌落直徑)/對照組菌落直徑]×100%。此外,在光學(xué)顯微鏡下觀察經(jīng)處理后的炭疽菌菌絲結(jié)構(gòu)的變化情況。
1.2.3 樣本準(zhǔn)備 菌株DZY6715在LB固體培養(yǎng)基上培養(yǎng)24 h后,取適量轉(zhuǎn)移到LB液體培養(yǎng)基,以28℃,180 r/min在搖床上培養(yǎng)。根據(jù)菌株生長曲線的結(jié)果,收集生長前期(t=24 h)和生長后期(t=72 h)菌液,置于-80℃冰箱保存。菌株DZY6715在24 h和72 h的編號(hào)分別為DZYB24h和DZYB72h。
1.2.4 樣本提取 -80℃冰箱凍存的樣本送至上海中科新生命生物科技有限公司完成菌懸液的樣品制備、QC制備、樣品LC-MS/MS質(zhì)譜分析和生物信息學(xué)分析。
1.2.5 差異代謝物分析 本文通過有監(jiān)督的偏最小二乘法分析(PLS-DA)方法建立判別模型來區(qū)分組間的總體差異,以O(shè)PLS-DA VIP>1和P value<0.05為標(biāo)準(zhǔn)篩選差異代謝物,并在KEGG數(shù)據(jù)庫(https:www.kegg.jpkeggpathway.html)進(jìn)行代謝通路注釋。
1.2.6 數(shù)據(jù)處理 試驗(yàn)每個(gè)處理3個(gè)重復(fù),數(shù)據(jù)和圖表處理采用Excel 2010和Origin 2023軟件,顯著性分析采用SPSS Statistics 20.0,差異水平為P<0.05。
2 結(jié)果
2.1 菌株DZY6715的生長曲線
菌株DZY6715在6-24 h是對數(shù)生長期,24-48 h是生長穩(wěn)定期,48 h以后開始表現(xiàn)出下降的趨勢,72 h進(jìn)入衰亡平穩(wěn)期(圖1)。生長穩(wěn)定期(生長前期)和衰亡期(生長后期)分別是代謝產(chǎn)物生產(chǎn)比較穩(wěn)定的時(shí)期和代謝產(chǎn)物累積較多的時(shí)期,因此,本文將對這兩個(gè)點(diǎn)的代謝產(chǎn)物進(jìn)行后續(xù)分析。

圖1 菌株DZY6715的生長曲線Fig.1 Growth curve of strain DZY6715
2.2 菌株DZY6715在不同生長期對炭疽菌的抑菌活性
圖2和圖3可以看出,t=24 h和t=72 h的菌液對油茶炭疽菌都有抑制作用,但二者的抑菌活性存在差異,表現(xiàn)為生長后期的抑菌活性較高。另外,隨著培養(yǎng)時(shí)間的遞增,抑菌率也隨之增加,并且72 h處理組的抑菌率始終高于24 h處理組,在培養(yǎng)過程中,二者的抑菌率范圍分別為18.05%-55.25%和11.82%-48.88%(培養(yǎng)第3-7天)。另外,對處理后的菌絲形態(tài)觀察發(fā)現(xiàn),對照組的菌絲通直,粗細(xì)均勻,伸展性好,未出現(xiàn)膨大加粗或細(xì)胞空泡化現(xiàn)象,而24 h和72 h處理組的菌絲均出現(xiàn)不同程度的畸形,具體表現(xiàn)為,24 h處理組的菌絲細(xì)胞長度變短,中央細(xì)胞膨大成橢圓形,相比24 h處理組,72 h處理組的菌絲細(xì)胞長度更短,表現(xiàn)為整體均勻膨大,菌絲細(xì)胞側(cè)邊分支產(chǎn)生球狀或念珠狀的數(shù)量較多,整體來說,72 h處理組的畸形現(xiàn)象更突出(圖4)。
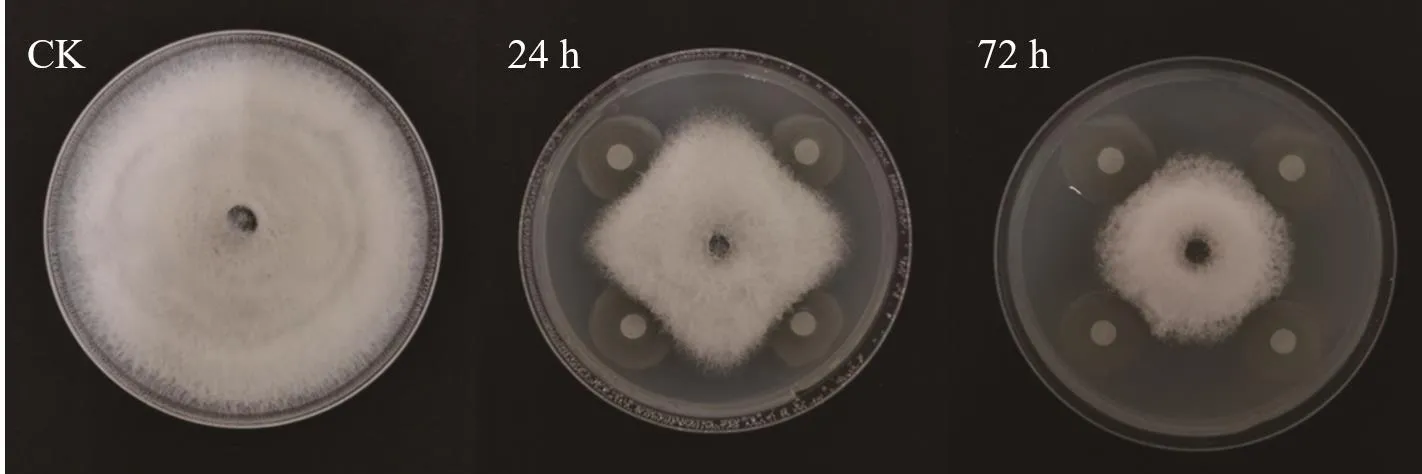
圖2 菌株DZY6715在不同生長期對油茶炭疽菌的抑菌效果Fig.2 Antifungal effects of strain DZY6715 against C.fructicola at different growth stages
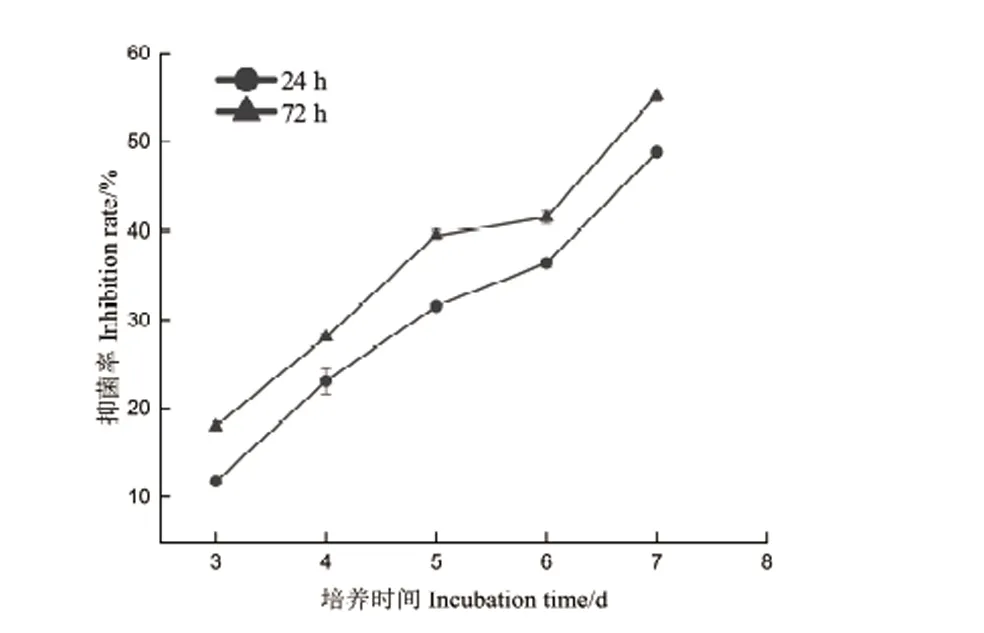
圖3 菌株DZY6715在不同生長期對油茶炭疽菌的抑菌率Fig.3 Inhibition rate of strain DZY6715 against C.fructicola at different growth stages

圖4 不同生長期的菌株DZY6715處理后對油茶炭疽菌菌絲形態(tài)的影響(10×100倍)Fig.4 Effects of strain DZY6715 on mycelial morphology of C.fructicola at different growth stages(10×100 times)
2.3 數(shù)據(jù)質(zhì)控
QC工作是進(jìn)行基于質(zhì)譜技術(shù)的代謝組學(xué)研究時(shí)獲得可靠且高質(zhì)量的代謝組學(xué)數(shù)據(jù)的基礎(chǔ),采用主成分分析方法(PCA)對數(shù)據(jù)進(jìn)行質(zhì)控分析。如圖5所示,紫色QC樣本檢測結(jié)果密集的聚在一起,說明QC重復(fù)性良好,分析系統(tǒng)穩(wěn)定,所采集的樣品數(shù)據(jù)可以進(jìn)一步差異分析。

圖5 QC樣本的PCA得分圖Fig.5 PCA score chart of the QC samples
2.4 主成分(PCA)分析
不同顏色表示不同生長時(shí)期的樣品,同一顏色表示同一生長時(shí)期下的樣品重復(fù)(圖6)。DZYB 24h和DZYB 72h兩組的樣本點(diǎn)均分布集中,說明組內(nèi)樣本重復(fù)性較好,不同處理間樣本點(diǎn)分布明顯分離,說明DZYB 24h和DZYB 72h的代謝物存在差異,本次數(shù)據(jù)質(zhì)量高,檢測性好。
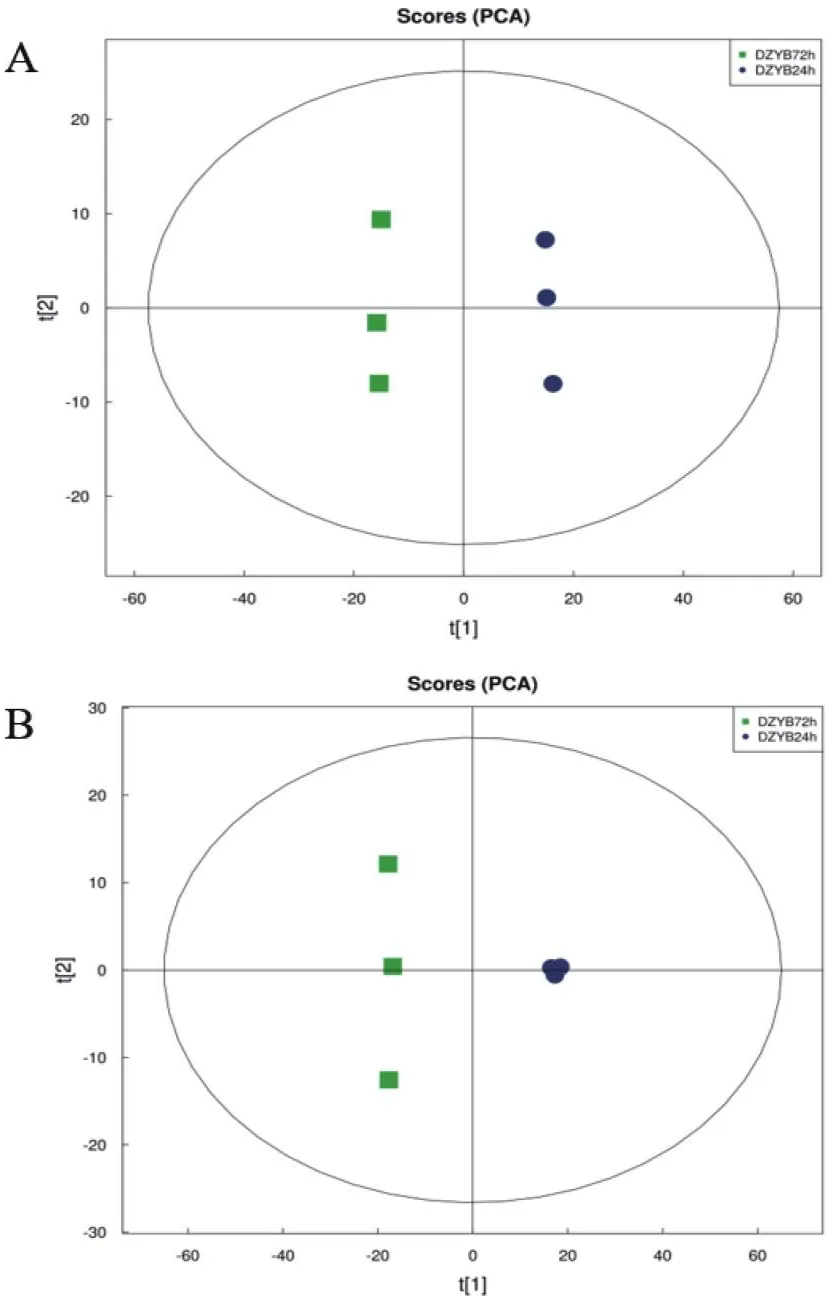
圖6 DZYB72h-vs-DZYB24h的PCA得分圖Fig.6 PCA score chart of DZYB72h-vs-DZYB24h
2.5 偏最小二乘判別(PLS-DA)分析
經(jīng)7-fold cross-validation(7次循環(huán)交互驗(yàn)證)得到的模型評價(jià)參數(shù)(R2和Q2),一般Q2和R2大于0.5,表明模型穩(wěn)定可靠。如圖7所示,正、負(fù)離子模式下,樣本在空間位置上聚類明顯,分成兩組,表明PLS-DA模型穩(wěn)定可靠,組間有明顯的差異。為避免有監(jiān)督模型在建模過程中發(fā)生過擬合,采用置換檢驗(yàn)(Permutation test)對模型進(jìn)行檢驗(yàn),可以看出隨著置換保留度逐漸降低,隨機(jī)模型的R2和Q2均逐漸下降,說明原模型不存在過擬合現(xiàn)象,模型穩(wěn)健性良好。

圖7 DZYB72h-vs-DZYB24h的得分圖(A1, B1)和置換檢驗(yàn)圖(A2, B2)Fig.7 PLS-DA score(A1, B1)and permutation test chart(A2, B2)of DZYB72h-vs-DZYB24h
2.6 差異代謝物篩選
把OPLS-DA VIP>1和P value < 0.05作為差異代謝物的篩選標(biāo)準(zhǔn)。將樣本中正負(fù)離子模式合并后篩選到差異代謝物共239個(gè),100個(gè)上調(diào),139個(gè)下調(diào)(圖8)。
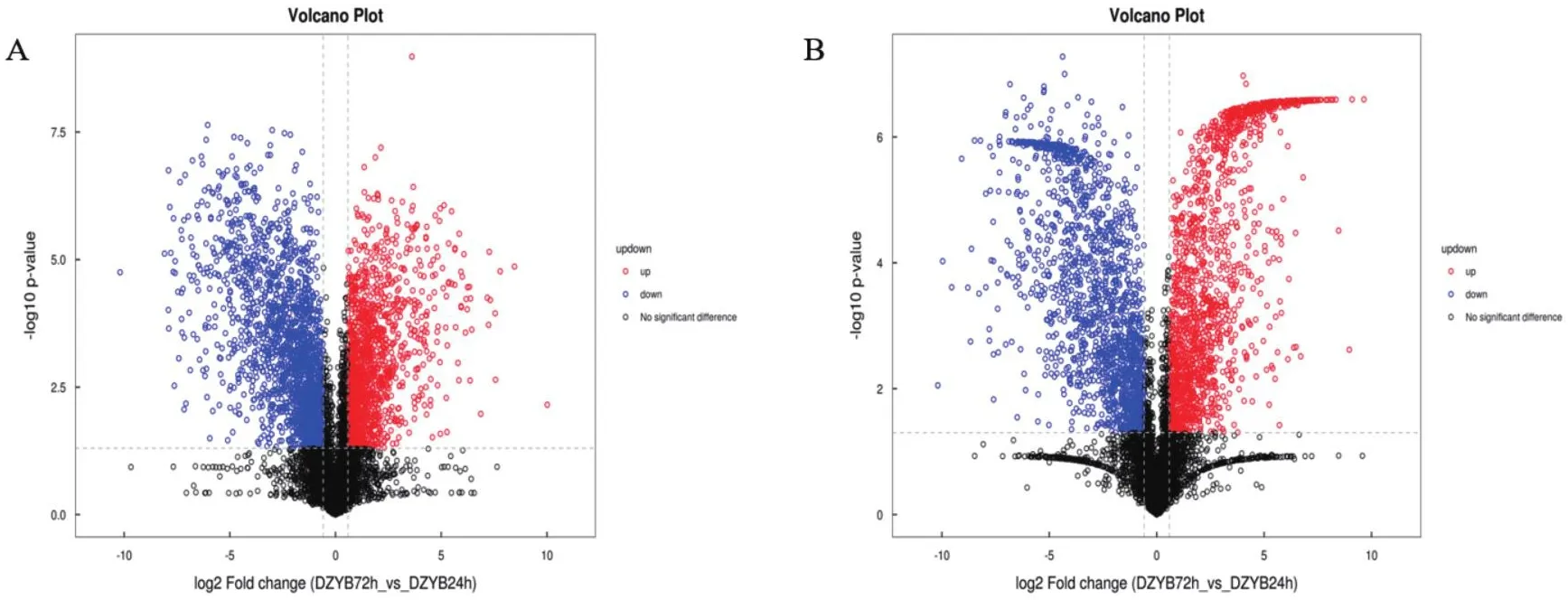
圖8 DZYB72h-vs-DZYB24h的差異代謝物火山圖Fig.8 Differential metabolite volcano plot of DZYB72h-vs-DZYB24h
2.7 KEGG Pathway分析
DZYB72h_vs_DZYB24h樣本中,顯著性差異代謝物注釋到了32條代謝通路中,Top20的代謝通路被顯示在圖9中,共富集到差異代謝物183個(gè)。顯著性通路分別為:蛋白質(zhì)消化和吸收,氨酰tRNA生物合成,氨基酸的生物合成,礦物吸收,ABC轉(zhuǎn)運(yùn)蛋白,賴氨酸降解,2-氧代羰酸代謝,各種次生代謝物的生物合成/第3部分,氰氨基酸代謝,纈氨酸、亮氨酸和異亮氨酸生物合成,精氨酸生物合成,精氨酸和脯氨酸代謝,Tropanc、哌啶和吡啶生物堿的生物合成,泛酸和CoA生物合成,苯丙氨酸代謝,mTOR信號(hào)通路,苯乙烯降解,谷胱甘肽代謝,丙氨酸、天冬氨酸和谷氨酸代謝,芥子油苷生物合成,酪氨酸代謝,軸突再生,硫胺素代謝,半胱氨酸和蛋氨酸代謝,組氨酸代謝,β-丙氨酸代謝,嘧啶代謝,甘氨酸、絲氨酸和蘇氨酸代謝,賴氨酸生物合成,神經(jīng)活性配體-受體相互作用,GABA能突觸,cGMP-PKG信號(hào)通路。
Pathway通路分析發(fā)現(xiàn),蛋白質(zhì)消化和吸收、氨酰tRNA生物合成、氨基酸的生物合成、ABC轉(zhuǎn)運(yùn)蛋白、2-氧羰基酸代謝這5條通路在拓?fù)浞治鲋械挠绊戄^大,重要性在0.2174-0.3830之間,差異代謝物的數(shù)量分別為19、14、19、17、15(表1),后續(xù)主要重點(diǎn)分析這些通路。

表1 5條重要代謝通路中的差異代謝物Table 1 Differential metabolites in 5 metabolic pathways
分析這5條代謝通路發(fā)現(xiàn),共涉及27種差異代謝物,主要是氨基酸、有機(jī)酸、高檸檬酸鹽、苯酚、腺苷、N-乙酰-D-氨基葡萄糖、諾氟沙星、硫胺素(表2)。其中,氨基酸占比最高,為61%,其次是有機(jī)酸,占比為18%,說明氨基酸和有機(jī)酸對細(xì)胞生長前期和后期的影響較大,可能調(diào)控著細(xì)胞的生命過程。

表2 5條代謝通路的差異代謝物分析Table 2 Differential metabolite analysis of five metabolic pathways
2.8 差異代謝物表達(dá)量分析
菌株DZY6715生長前期氨基酸總的表達(dá)量顯著高于生長后期,并且各氨基酸在不同生長階段的表達(dá)量也存在差異,總體上生長前期的表達(dá)量高于生長后期。在兩個(gè)生長階段中,DL-纈氨酸、甘氨酸、L-天冬氨酸的表達(dá)量較低。在生長前期,精氨酸、谷氨酸、異亮氨酸、亮氨酸大量表達(dá),分別是生長后期的22.02、2.10、2.02、2.49倍;在后期,賴氨酸、苯丙氨酸、DL-色氨酸、DL-絡(luò)氨酸的表達(dá)量顯著升高,分別是前期的1.48、1.53、1.19、1.88倍(圖10-A)。因此,認(rèn)為精氨酸、谷氨酸、異亮氨酸、亮氨酸在菌株DZY6715生長前期起著主導(dǎo)作用,更側(cè)重于參與細(xì)胞生長、能量代謝等過程,而賴氨酸、苯丙氨酸、DL-色氨酸、DL-絡(luò)氨酸可能主要對后期階段中的脅迫因素起緩解作用。
有機(jī)酸在生長后期的總表達(dá)量明顯高于生長前期。前期,鄰苯二甲酸和丙酸的表達(dá)量明顯高,分別是后期的3.64、8.46倍,而異戊酸、2-氨基己二酸、丁酸在后期的表達(dá)量顯著高于前期,分別是前期的3.00、3.18、13.66倍(圖10-B)。這說明有機(jī)酸都參與了細(xì)胞的不同生長階段,并且后期階段累積的有機(jī)酸更多,這對緩解后期營養(yǎng)、空間不足等的壓力有重要作用。
諾氟沙星、N-乙酰-D-氨基葡萄糖、腺苷、苯酚、高檸檬酸鹽的后期表達(dá)量遠(yuǎn)高于前期,分別是前期的1.67、45.39、5.97、49.35、5.61倍,而硫胺素前期的表達(dá)量比后期高117.23倍(圖10-C),說明不同生長階段中,各代謝物的表達(dá)量存在差異,抑或是代謝物表達(dá)量的高低因不同生長階段而異。
3 討論
本文生物學(xué)試驗(yàn)發(fā)現(xiàn),與生長前期(t=24 h)相比,生長后期(t=72 h)的菌株DZY6715處理油茶炭疽菌后的菌絲細(xì)胞畸形現(xiàn)象更明顯,說明經(jīng)生長后期(t=72 h)處理的病原菌絲生長受限更嚴(yán)重[12],同時(shí)也表明了菌株DZY6715生長后期(t=72 h)的抑菌活性更強(qiáng)。
氨基酸是許多細(xì)胞生物合成和代謝過程中的主要代謝產(chǎn)物,不僅作為前體物質(zhì)參與細(xì)胞構(gòu)建,還通過形成催化酶調(diào)節(jié)細(xì)胞代謝[13]。本文發(fā)現(xiàn)菌株DZY6715在生長后期和前期的差異代謝物中氨基酸占比最高,并且前期氨基酸的表達(dá)量顯著高于后期,這可能是在細(xì)胞開始生長繁殖的階段,需要大量氨基酸提供能量以維持正常的生命活動(dòng),而隨著生長期的后移,積累和表達(dá)的氨基酸更側(cè)重于緩解生活環(huán)境中營養(yǎng)、空間等不足的壓力。這與本研究中生物學(xué)試驗(yàn)部分的結(jié)果相吻合,進(jìn)一步說明氨基酸既有構(gòu)建細(xì)胞結(jié)構(gòu)的功能,又能發(fā)揮抑菌作用,但是前者可能占主導(dǎo)地位。氨基酸及其衍生物具有較好的抗菌效果,氨基酸的積累有利于細(xì)胞對氧化脅迫、不利因素的抵抗[14]。例如,甘氨酸是抗菌活性較強(qiáng)的自由氨基酸,不僅能抑制病菌的生長繁殖,還可以參與谷胱甘肽抗氧化過程[15]。賴氨酸的生物合成為蛋白質(zhì)的合成和細(xì)菌肽聚糖細(xì)胞壁的構(gòu)建提供了必要的成分,這對于細(xì)胞的生長繁殖來說是必不可少的[16]。也有研究表明,色氨酸在細(xì)胞體內(nèi)具有多種代謝功能,通常整合到酶和蛋白質(zhì)的多肽鏈中,待完全分解后參與代謝過程,B.megaterium 和Rhodococcus erythropolis可以用色氨酸作為生長過程中所需的碳源和氮源[17]。芳香族氨基酸(苯丙氨酸、絡(luò)氨酸、色氨酸)和支鏈氨基酸(異亮氨酸、亮氨酸)等可以通過形成疏水區(qū)域來保護(hù)蛋白質(zhì)免受膽汁壓力[18],苯丙氨酸和色氨酸可以增強(qiáng)水稻根系的耐受性,有效減少逆境下活性氧的大量產(chǎn)生[19]。學(xué)者研究表明,一氧化氮對植物炭疽菌分生孢子的萌發(fā)和附著胞的形成具有調(diào)控作用,而精氨酸作為一氧化氮的前體在此過程中起著關(guān)鍵作用[20]。Takagi等[21]發(fā)現(xiàn)脯氨酸以及谷氨酸、精氨酸、賴氨酸等有助于提高細(xì)胞對低溫的耐受性。精氨酸在脅迫下可以通過維持細(xì)胞的完整性、穩(wěn)固細(xì)胞壁和細(xì)胞質(zhì)膜來防止細(xì)胞受損傷,參與調(diào)控活性氧的動(dòng)態(tài)平衡,保護(hù)細(xì)胞在脅迫下的正常生長[22]。氨基酸的匱乏會(huì)降低細(xì)胞應(yīng)對脅迫的能力,還會(huì)使蛋白質(zhì)合成缺少原料,即便與蛋白質(zhì)合成相關(guān)的氨酰-tRNA生物合成途徑被激活,也不足以完全解除蛋白質(zhì)的合成阻礙,導(dǎo)致細(xì)胞生長受限,而酵母細(xì)胞通過減少氨基酸代謝途徑降低了能量的需求,維持碳、氮代謝平衡,使其在逆境中得以存活[23]。因此,認(rèn)為氨基酸既是維持細(xì)胞生長和代謝平衡的重要物質(zhì),也是細(xì)胞抵抗脅迫的重要調(diào)節(jié)劑。
多方研究已證實(shí)有機(jī)酸可以抑制病原菌生長,增強(qiáng)有機(jī)體抗性,其主要是通過能量競爭、改變細(xì)胞膜通透性、改變滲透壓、抑制大分子合成和誘導(dǎo)抗菌肽產(chǎn)生等發(fā)揮抑菌作用。有機(jī)酸通過刺激宿主植物產(chǎn)生抗菌肽物質(zhì)作用于磷脂雙分子層,增大病原菌細(xì)胞膜的通透性,導(dǎo)致細(xì)胞內(nèi)容物外泄,殺死病原菌,但是相比之下,有機(jī)酸在細(xì)胞內(nèi)解離出質(zhì)子和酸根離子而發(fā)揮抑菌作用的效果更為顯著,并且抑菌效果與質(zhì)子和酸根離子呈正相關(guān)[24]。另外,有機(jī)酸通過影響細(xì)胞內(nèi)的酸堿性而抑制病原菌菌絲的生長[25]。研究表明,烏梅有機(jī)酸提取液對枯草桿菌、金黃色葡萄球菌、大腸桿菌和四聯(lián)球菌都具有較好的抑制作用[26]。乳酸菌發(fā)酵產(chǎn)生的丁酸對霉菌起抑菌作用[27]。Wang等[28]通過半制備分離獲得了對有害菌具有顯著抑菌作用的物質(zhì)苯乳酸。本文研究也發(fā)現(xiàn)在細(xì)胞生長的不同階段,都涉及有機(jī)酸的參與,尤其是生長后期有機(jī)酸大量積累和表達(dá),推測有機(jī)酸在細(xì)胞抵抗逆境中發(fā)揮著重要作用,這可能也是菌液在生長后期(t=72 h)的抑菌效果高于生長前期(t=24 h)的又一重要因素。
硫胺素(又名維生素B1)在生長前期的表達(dá)量遠(yuǎn)遠(yuǎn)高于后期,推斷硫胺素可能有助于細(xì)胞的生長繁殖。同時(shí),B族維生素對于菌體的生長過程來說是極其重要的一個(gè)生長因子,還是生物體內(nèi)多種酶的輔酶[29-30],硫胺素的大量表達(dá)能夠增強(qiáng)酶的活性,進(jìn)而加速菌體的生長繁殖速度。腺苷能夠間接參與細(xì)胞內(nèi)的能量轉(zhuǎn)移、信號(hào)轉(zhuǎn)導(dǎo)等過程[31],而本文發(fā)現(xiàn)菌株DZY6715生長的后期腺苷的表達(dá)量明顯高于前期,認(rèn)為腺苷在前期階段主要是參與能量代謝的過程,而后期階段大量表達(dá)的腺苷可能是在響應(yīng)壓力過程中傳遞重要信號(hào)的。此外,菌株DZY6715在生長后期苯酚、N-乙酰-d-氨基葡萄糖、諾氟沙星、高檸檬酸鹽等差異代謝物高表達(dá),推測這些代謝物質(zhì)更主要的是參與調(diào)控細(xì)胞的抗逆生長過程,這些物質(zhì)在響應(yīng)脅迫中的重要地位已被學(xué)者所證實(shí),N-乙酰-d-氨基葡萄糖是一種特殊的單糖,是生物體內(nèi)絕大部分多糖的組成單位,不僅甜度較高,還有消炎、抗氧化等的功能[32-33]。苯酚是一種芳香族化合物,屬于酚類,有毒性,會(huì)對生物體的生長造成不良影響[34-35]。諾氟沙星是第3代氟喹諾酮類的一種抗生素,會(huì)阻礙消化道內(nèi)致病細(xì)菌的DNA旋轉(zhuǎn)酶的作用,阻礙細(xì)菌DNA復(fù)制[36-37]。所以,本研究認(rèn)為后期階段這些代謝物質(zhì)高表達(dá)的原因,不僅是為有限空間內(nèi)營養(yǎng)的不足提供能量,更偏向于利用這些代謝物來抵抗脅迫,這也印證了本文生物學(xué)試驗(yàn)部分的結(jié)果,即菌株DZY6715在生長后期(t=72 h)抑菌活性較生長前期(t=24 h)高。
綜上,本文通過非靶向代謝組學(xué)技術(shù)對菌株DZY6715在不同生長時(shí)期的差異代謝物及差異代謝物的表達(dá)量進(jìn)行了研究,剖析了該菌株在兩個(gè)不同的生長時(shí)期抑菌活性存在差異的原因,這為后續(xù)這些重要差異代謝物的靶向分析奠定了基礎(chǔ),同時(shí)有利于該菌株有效抑菌物質(zhì)的提取,使該菌株能更有效的應(yīng)用到農(nóng)林業(yè)生產(chǎn)的生物防治中。
4 結(jié)論
本文基于菌株生長曲線和生物學(xué)試驗(yàn)得出,菌株DZY6715生長后期的抑菌活性高于生長前期,并且隨著培養(yǎng)時(shí)間的遞增,效果越明顯,結(jié)合非靶向代謝組學(xué)技術(shù)探討了該菌株生長前期和生長后期的差異代謝物及其調(diào)控機(jī)制,綜合分析認(rèn)為,氨基酸主要是參與細(xì)胞的生長繁殖、能量代謝等,同時(shí)具有抑菌的作用;有機(jī)酸、硫胺素、腺苷、高檸檬酸鹽等差異代謝物是抵抗壓力的重要調(diào)節(jié)劑,是該菌株表現(xiàn)高抑菌活性的關(guān)鍵因子。
猜你喜歡
音樂探索(2022年2期)2022-05-30 21:01:37
小讀者(2021年2期)2021-03-29 05:03:48
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時(shí)刊(2019年13期)2019-11-17 14:59:54
小天使·一年級(jí)語數(shù)英綜合(2019年8期)2019-08-27 02:23:00
中國特種設(shè)備安全(2018年11期)2019-01-08 02:08:32
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
小學(xué)科學(xué)(學(xué)生版)(2018年7期)2018-08-13 09:33:04
紅領(lǐng)巾·萌芽(2016年1期)2016-09-10 07:22:44
