后生元調(diào)節(jié)肥胖的研究進展
2023-10-17 07:03:06汪玲娥龐立冬李鴻萱丁弋芯祁雪鶴葉亞明滿朝新姜毓君
食品科學 2023年17期
關(guān)鍵詞:小鼠
汪玲娥,龐立冬,李 譽,李鴻萱,丁弋芯,黃 燕,祁雪鶴,葉亞明,滿朝新,姜毓君,*
(1.東北農(nóng)業(yè)大學食品學院,乳品科學教育部重點實驗室,黑龍江 哈爾濱 150030;2.湖北均瑤大健康飲品股份有限公司,湖北 宜昌 443000)
肥胖概念首次被提出是在1620年,來自英國的醫(yī)生Venner認為肥胖是上層人士的一種“職業(yè)病”[1]。19世紀美國營養(yǎng)學家Ancel Keys提出的體質(zhì)量指數(shù)(body mass index,BMI)被用作衡量肥胖的指標并沿用至今[1]。如今全球超重(BMI≥25 kg/m2)和肥胖(BMI≥30 kg/m2)人群比例成倍增加,主要原因是高能量低營養(yǎng)的飲食習慣以及鍛煉的減少[2]。肥胖可導(dǎo)致人體多個系統(tǒng)發(fā)生病變,許多研究已證實心臟病、糖尿病和中風等各種慢性疾病均與肥胖有著密切關(guān)聯(lián)[3-7]。
2013年,西班牙學者Katerina正式提出“后生元(postbiotics)”概念[8-9]。2021年國際益生菌和益生元科學學會(International Scientific Association for Probiotics and Prebiotics,ISAPP)發(fā)布后生元共識聲明,將其定義為“對宿主健康有益的無生命微生物和/或其成分的制劑”[10]。后生元通過特定的滅活方式,使原本具有活性的微生物喪失活性和生長繁殖能力,但同時一定程度上保留其結(jié)構(gòu)、特性和功效。一般來說后生元可分為菌體成分和菌代謝產(chǎn)物,這些成分包括肽聚糖衍生的胞肽、胞外多糖(exopolysaccharides,EPS)、磷壁酸和構(gòu)成細胞壁成分的表面突出分子(如菌毛、性菌毛或鞭毛)、分泌的蛋白質(zhì)/肽、細菌素(如嗜酸蛋白、羅伊氏菌素等)、細胞-游離上清液、有機酸(如乳酸和乙酸)、維生素、短鏈脂肪酸(short chain fatty acids,SCFAs)(如丁酸和丙酸鹽)、神經(jīng)遞質(zhì)、生物表面活性劑等[11]。
近年來已經(jīng)有不少學者研究了后生元可能具有的健康益處[12-15],目前后生元已經(jīng)被應(yīng)用于普通食品、營養(yǎng)保健品、特殊醫(yī)學食品、嬰幼兒食品等中,隨著后生元研究的不斷深入,其逐漸開始被應(yīng)用于預(yù)防或治療疾病。臨床研究表明,后生元可有效緩解急/慢性腹瀉(Crohn’s disease,CD)、調(diào)節(jié)免疫功能、改善過敏反應(yīng)[16],同時其在預(yù)防肥胖及改善與之相關(guān)代謝紊亂方面的研究報道也越來越多[17-19]。后生元已成為一種潛在的新興治療肥胖手段,部分有關(guān)后生元健康作用的臨床研究匯總?cè)绫?所示。因此,本文對肥胖的危害和形成機制進行介紹,討論后生元對人體發(fā)揮有益作用的潛在機制,綜述后生元改善肥胖作用的研究進展,為推動臨床治療肥胖的研究及相關(guān)后生元藥物或制劑的開發(fā)提供參考。
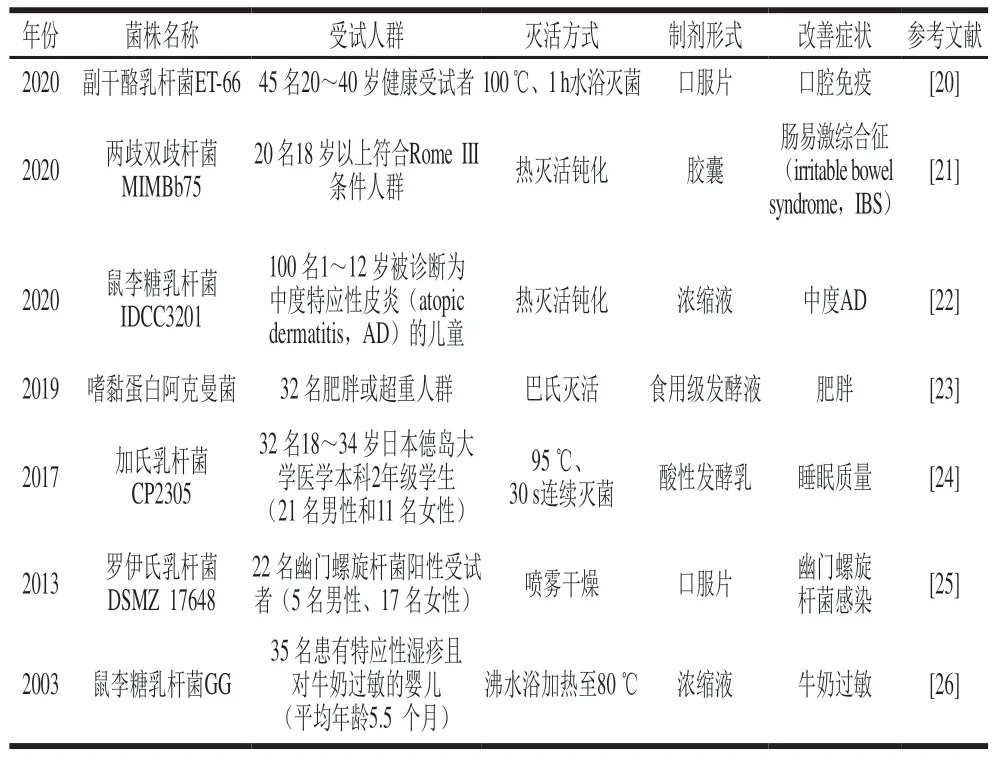
表1 有關(guān)后生元健康作用的臨床研究Table 1 Clinical studies on the health promoting effects of postbiotics
1 肥胖的危害作用及其形成機制
1.1 危害作用
肥胖被世界衛(wèi)生組織(World Health Organization,WHO)認定為一種慢性疾病,患有肥胖的同時常伴隨多種代謝疾病的發(fā)生[27],近年來有關(guān)肥胖危害的研究數(shù)量較多,表2總結(jié)了2017—2022年有關(guān)肥胖危害的研究。根據(jù)《中國居民營養(yǎng)與慢性病狀況報告(2020年)》顯示,我國已有超過一半的成年人處于超重或肥胖狀態(tài)[28]。肥胖不僅會增加2型糖尿病(type 2 diabetes mellitus,T2DM)、心血管疾病、慢性腎病等一系列代謝疾病發(fā)生[3,29-31],還會對大腦小膠質(zhì)細胞造成損傷,加快大腦老化[32]。最新研究發(fā)現(xiàn)肥胖可顯著增加抑郁和焦慮的風險[33]。

表2 2017—2022年關(guān)于肥胖的危害研究Table 2 Studies on the dangers of obesity in 2017–2022
1.2 形成機制
1.2.1 能量失衡
根據(jù)能量守恒定律的普遍解釋,攝入能量超過消耗能量會導(dǎo)致身體脂肪增加[54],這也就是傳統(tǒng)的能量平衡模型(conventional model,CM)[55]。過量進食會對胰島素分泌產(chǎn)生作用,機體將葡萄糖轉(zhuǎn)化為脂肪組織,影響脂肪細胞中的熱量沉積,這一過程通過對脂肪細胞的直接影響和削弱食物攝入的循環(huán)代謝從而導(dǎo)致肥胖。這一觀點將肥胖主要歸因于食物的過量攝入,認為所有能量對身體來說都是一樣的,沒有考慮神經(jīng)內(nèi)分泌等生理過程機制[56]。實驗性過度喂食動物和人類增加體質(zhì)量可誘導(dǎo)肥胖[57-58]為這一觀點提供了有力證據(jù)。
1.2.2 選擇性代謝
能量守恒定律的第2種解釋是身體脂肪質(zhì)量的增加推動正能量平衡[54]。有別于傳統(tǒng)的CM,碳水化合物-胰島素模型(carbohydrate-insulin model,CIM)[55]提出了飲食與肥胖因果方向的反轉(zhuǎn),即飲食的關(guān)鍵作用是代謝,能量對慢性脂質(zhì)儲存或有獨立效應(yīng),碳水化合物的攝入是導(dǎo)致肥胖的主要原因。高碳水化合物飲食會導(dǎo)致餐后血糖升高,刺激胰島素分泌,促進脂肪儲存,導(dǎo)致血液中糖、脂等供能分子不足,這一過程改變能量物質(zhì)分配,致使饑餓感和能量攝入增加、降低代謝率,從而導(dǎo)致肥胖。這一觀點認為碳水化合物和/或某些特定食物成分的攝入是肥胖的主要原因,胰島素和脂肪細胞在肥胖形成中發(fā)揮著核心作用[56]。相比于傳統(tǒng)模型,CIM納入了人體生理過程和神經(jīng)內(nèi)分泌機制等因素。
CIM假設(shè)了雙向因果關(guān)系(推-拉模型)[54]。在這樣的綜合模型中,人類天生對高能量和高適口性食物的偏好共同促進了正能量平衡。在過度進食后的前幾個小時,機體會將多余能量推入脂肪組織,在進食后期,飲食對激素的影響(選擇性代謝)會引發(fā)一種拉動機制,身體會增加胰島素分泌量,并抑制胰高血糖素的分泌,同時機體向脂肪細胞發(fā)出信號,讓其儲存更多的能量,從而減少肌肉和其他代謝活躍組織所需的能量,當大腦感知到身體沒有獲得足夠能量時,會導(dǎo)致機體饑餓感的產(chǎn)生;同時,新陳代謝也可能會在身體試圖保存能量的過程中減慢。基于這一假設(shè),上述兩種機制共同促進了脂質(zhì)儲存。
2 后生元的潛在作用機制
后生元可通過體內(nèi)系統(tǒng)反應(yīng)與腸腔原位反應(yīng)調(diào)節(jié)機體代謝。體內(nèi)系統(tǒng)反應(yīng)指后生元經(jīng)過熱滅活處理,由于其具有小分子的特性,可以通過腸道屏障更快、更直接地作用于人體,增強人體免疫力、平衡腸道菌群、調(diào)節(jié)生理功能[59]。腸腔原位反應(yīng)指脂壁酸等化合物可以干擾有害菌在腸腔中的生長,減少有害菌彼此間的聯(lián)系,并平衡腸道有益菌和有害菌比例,進而維持人體腸道健康[59]。因此,后生元具有的健康益處可能是由許多不同機制所驅(qū)動的(圖1)。目前,科學家們已經(jīng)發(fā)現(xiàn)了各種形式的后生元,經(jīng)過研究,由益生菌細胞產(chǎn)生的各種后生元成分可以通過不同途徑對機體代謝進行有益調(diào)節(jié),并且有的后生元和益生菌的益生作用相似,在某些情況下,其作用機制也可能與已知的益生菌作用機制類似,但后生元不依賴于細菌的活性,同時這些后生元的益生機制也可以獨立或聯(lián)合發(fā)揮作用[9]。到目前為止,后生元在人類或動物體內(nèi)發(fā)揮益生功效的作用機制尚未完全闡明。
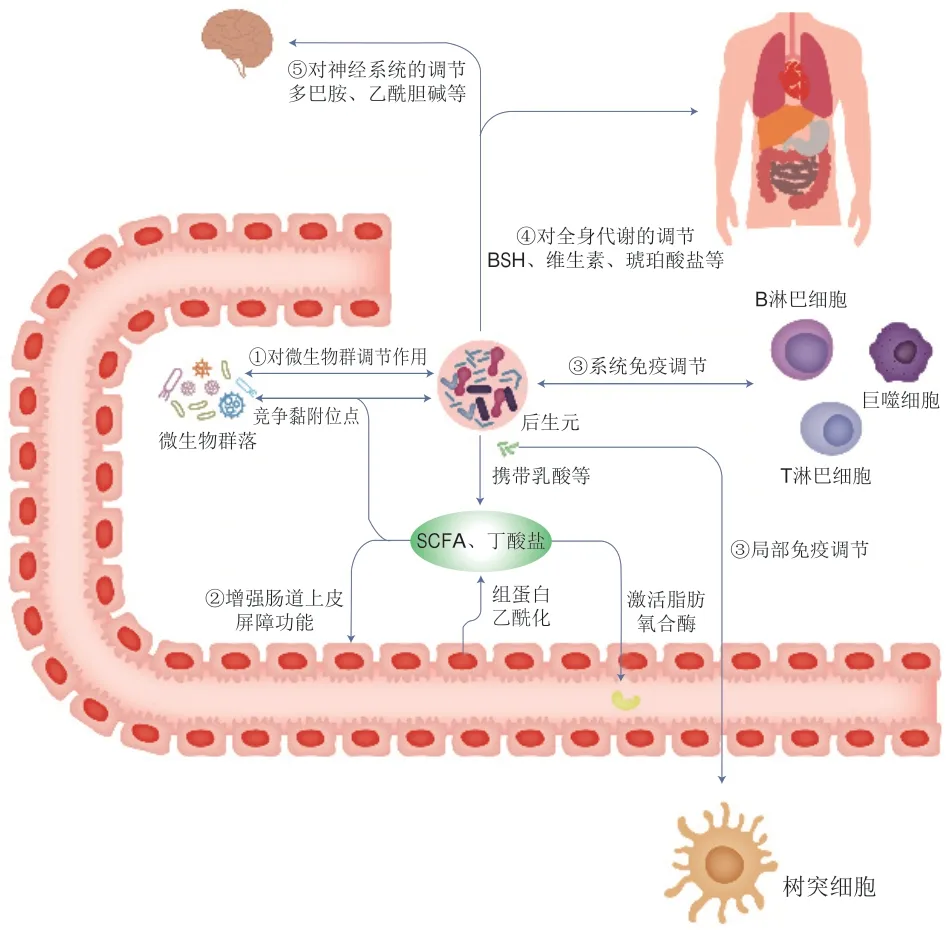
圖1 后生元潛在作用機制Fig.1 Potential mechanism of action of postbiotics
2.1 對微生物群的穩(wěn)態(tài)調(diào)節(jié)
雖然后生元已無生命特征,但其中的許多成分具有生物活性,對腸道微生物具有調(diào)節(jié)改善作用。如細菌素(一種細胞外抗菌肽)可以抑制與致病性密切相關(guān)的微生物,被認為是腸道微生物群落多樣性的重要貢獻者[60]。另一后生元成分乳酸也具有抗菌活性[61-62]。在革蘭氏陽性菌的細胞壁化合物中,磷壁酸與肽聚糖共價連接形成陰離子糖聚合物與磷酸二酯連接的重復(fù)單元,在細胞結(jié)構(gòu)、細胞分裂控制中起關(guān)鍵作用,有利于宿主細菌間的相互作用和細胞生理學中其他關(guān)鍵生化和代謝作用。此外,后生元中可能存在群體感應(yīng)和群體淬滅分子[63],腸道中的某些微生物可以利用后生元中攜帶的乳酸,產(chǎn)生對機體有益的SCFAs和丁酸鹽。如果處理后的后生元中仍含有保持原活性的黏附成分(如菌毛和凝集素),那么后生元也可以與常駐微生物競爭黏附位點[64],發(fā)揮益生功效。
2.2 強化腸道上皮屏障功能
分泌蛋白(如分泌蛋白Msp1/p75和Msp1/p40,或來自鼠李糖乳桿菌GG的蛋白質(zhì)[65])可介導(dǎo)增強腸道上皮屏障功能[9]。研究發(fā)現(xiàn)熱滅活的雙歧桿菌MIMBb75可緩解IBS,可能的原因是滅活后的細菌及其代謝物依然能夠附著在腸道內(nèi)壁細胞上,加強腸道屏障,從而抵御可能導(dǎo)致IBS的有害細菌和毒素[21]。后生元中的SCFAs如果達到足夠的水平,也可能會改變腸道上皮屏障功能,并抵御脂多糖(lipopolysaccharide,LPS)誘導(dǎo)的腸道屏障功能障礙[66]。例如,在Caco-2細胞中,后生元組分丁酸鹽可依賴肌動蛋白結(jié)合蛋白突觸足蛋白(recombinant synaptopodin,SYNPO)促進腸道屏障功能,加速腸上皮細胞損傷修復(fù)[67];或通過組蛋白乙酰化激活脂肪氧合酶,從而改變緊密連接的通透性[68]。
2.3 免疫調(diào)節(jié)
后生元對機體局部和全身的免疫調(diào)節(jié)活動,通常由微生物相關(guān)分子與免疫細胞的特定識別受體相互作用來介導(dǎo),進而導(dǎo)致各種細胞因子和免疫調(diào)節(jié)劑的表達[9]。這表現(xiàn)在副干酪乳桿菌DG的EPS通過增強人類單核細胞系Tamm-Horsfall蛋白1(THP-1)中腫瘤壞死因子(tumor necrosis factor-α,TNF-α)和白細胞介素-6(interleukin-6,IL-6)基因的表達,特別是IL-8和趨化因子配體20(C-C chemokine ligand 20,CCL20)基因的表達,顯示出免疫刺激特性[69]。乳酸等代謝物也可通過腸道CX3CR1+細胞中GPR31依賴的樹突突起,介導(dǎo)免疫反應(yīng)[70]。同樣地,羅伊氏乳桿菌產(chǎn)生的色氨酸衍生物吲哚能激活小鼠腸道中CD4+T細胞的芳香烴受體,誘導(dǎo)其分化為CD4+CD8αα+雙陽性上皮內(nèi)淋巴細胞[71]。Qi等[72]對鼠李糖乳桿菌LGG不同后生元化合物免疫調(diào)節(jié)特性的研究表明,大多數(shù)后生元組分通常抑制LPS刺激的小鼠巨噬細胞RAW264.7中Toll樣受體(Toll-like receptors,TLR)、絲裂原活化蛋白激酶、細胞外調(diào)節(jié)蛋白激酶和核因子-κB信號通路的激活。
2.4 全身代謝調(diào)節(jié)
后生元對全身代謝的調(diào)節(jié)可直接由滅活微生物內(nèi)部或表面的代謝物以及酶來介導(dǎo)[9]。如BSH負責膽汁酸的解離,從而使微生物進行進一步的生物轉(zhuǎn)化,并使系統(tǒng)循環(huán)的膽汁酸池多樣化[73]。已有的研究表明副干酪乳桿菌細胞壁蛋白組分可通過誘導(dǎo)細胞凋亡從而抑制人結(jié)腸癌細胞系的生長[74]。同時,熱滅活短乳桿菌和副干酪乳桿菌可通過Bcl2關(guān)聯(lián)X蛋白(BCL2-associated X,Bax)/B淋巴細胞瘤-2(B-cell lymphoma-2,Bcl2)和半胱天冬酶途徑誘導(dǎo)HT-29人結(jié)腸癌細胞系的凋亡[74]。另一種后生元物質(zhì)琥珀酸鹽作為一種碳水化合物發(fā)酵的細菌中間體,也是腸道糖異生的底物,已被發(fā)現(xiàn)可改善小鼠的血糖水平[75]。一項關(guān)于線蟲的研究發(fā)現(xiàn),滅活副干酪乳桿菌D3-5菌株的胞壁成分脂磷壁酸(lipoteichoic acid,LTA)可通過促進腸道黏蛋白生成,從而減少衰老相關(guān)的腸道滲漏和炎癥,促進人體健康[76]。
2.5 作用于神經(jīng)系統(tǒng)
微生物可以代謝多種神經(jīng)活性物質(zhì),這些物質(zhì)可以作用于腸道和中樞神經(jīng)系統(tǒng),可能調(diào)節(jié)動物和人類的行為及認知功能。這些物質(zhì)包括幾種神經(jīng)遞質(zhì)(如血清素、多巴胺、乙酰膽堿和γ-氨基丁酸)以及能與大腦中表達受體結(jié)合的各種化合物(例如吲哚和膽汁酸)。微生物酶還可以代謝飲食前體以合成宿主神經(jīng)遞質(zhì),如色氨酸可用于血清素的合成,酪氨酸可用于多巴胺的合成,從而降低營養(yǎng)物質(zhì)的生物利用率[77]。除了酶之外,一些代謝物如SCFA可合成維生素(如VB),對中樞神經(jīng)系統(tǒng)功能有著重要的調(diào)節(jié)作用。
3 后生元改善肥胖研究進展
隨著后生元對人體作用機制研究的深入,利用后生元改善肥胖的研究也取得了初步進展,后生元形式多樣,不僅包括滅活的微生物菌株及其自身的細胞成分,還包括微生物產(chǎn)生的代謝產(chǎn)物等。目前一些高效的抗肥胖后生元在乳桿菌屬、雙歧桿菌屬及擬桿菌屬等[21,24-25,78-79]中均有發(fā)現(xiàn)。已有相關(guān)研究報道了一些具有減肥降脂作用的微生物代謝產(chǎn)物后生元,而目前對微生物細胞自身成分的抗肥胖作用鮮有研究,因此,本文主要從后生元菌株及菌體代謝物的角度綜述后生元在調(diào)節(jié)代謝以改善肥胖癥狀方面的研究進展,并對一些其他形式的減肥后生元進行補充說明。
3.1 后生元菌株改善肥胖癥狀研究進展
3.1.1 嗜黏蛋白阿克曼菌
嗜黏蛋白阿克曼菌(Akkermansia muciniphila,Akk)是一種腸道共生菌,Akk的豐度與肥胖呈負相關(guān),在腸道中占比約為3%~5%[80]。2016年比利時魯汶大學Plovier等[81]首次發(fā)現(xiàn)巴氏滅活(70 ℃、30 min)的Akk比活菌能夠更好地降低小鼠脂肪含量并減輕胰島素抵抗和血脂異常,這是滅活A(yù)kk作為后生元在改善肥胖癥狀方面優(yōu)于益生菌的有力證據(jù);此后的多項研究也證實了Akk的這一特性[82-86]。Akk代謝產(chǎn)物SCFA與腸上皮細胞受體FFAR3-FFAR2(又稱GPR43-GPR41)結(jié)合,刺激腸道內(nèi)胃腸激素胰高血糖素樣肽1(glucagon-like peptide-1,GLP1)和GLP2的分泌,從而有助于糖代謝和腸道屏障功能的調(diào)節(jié)。活菌在70 ℃巴氏殺菌30 min后某些細胞成分的活性會部分或全部保留[87-88],Akk外膜特異性蛋白Amuc_1100的耐熱特性正是Akk發(fā)揮減肥降脂作用的關(guān)鍵[23,81,85,89]。研究人員在比較Amuc_1100和活菌以及巴氏殺菌后的Akk對飼喂HFD小鼠的影響時發(fā)現(xiàn),飼喂Amuc_1100小鼠與飼喂巴氏殺菌后的Akk小鼠觀察到的情況類似,即Amuc_1100激活了TLR2,并單獨發(fā)揮了Akk的部分益生功能[85,89]。Cani[90]和Yoon[91]等發(fā)現(xiàn)了Akk的另一關(guān)鍵蛋白——分子質(zhì)量為84 kDa的P9蛋白[90-91]。研究發(fā)現(xiàn)Akk能夠增加小鼠肩胛間褐色脂肪組織(brown adipose tissue,BAT)的產(chǎn)熱,而對白色脂肪組織(white adipose tissue,WAT)無影響[91-92],用Akk的無細胞上清液處理人腸分泌L細胞(NCI-H716),觀察到其上清液中的P9蛋白刺激了GLP-1的分泌,而GLP-1是小腸分泌的一種抑制食欲的激素,可以調(diào)節(jié)宿主能量平衡、改善小鼠葡萄糖穩(wěn)態(tài),并且P9蛋白促進GLP-1分泌的作用比Amuc_1100更加顯著,而P9蛋白誘導(dǎo)GLP-1分泌的機制與鈣離子信號蛋白和細胞間黏附分子2(intercellular adhesion molecule 2,ICAM-2)有關(guān)(圖2)。
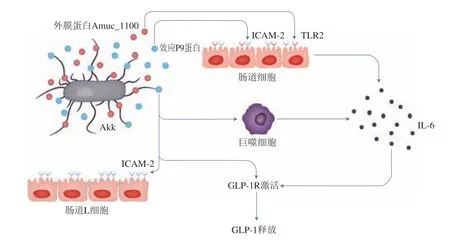
圖2 Akk外膜蛋白激活GLP-1R釋放GLP-1Fig.2 Akk outer membrane proteins activate GLP-1R to release GLP-1
3.1.2 雙歧桿菌屬
雙歧桿菌屬為具有抗肥胖功能的后生元菌株[92]。熱滅活(121 ℃、20 min)的動物雙歧桿菌乳亞種CECT8145(Ba8145)可通過增強碳水化合物氧化以增加能量消耗,從而介導(dǎo)肥胖,能量消耗的增強可能與線粒體功能的改善和BAT產(chǎn)熱相關(guān)基因的激活有關(guān)[93]。此外Ba8145能夠降低飼喂自助餐廳式膳食(cafeteria diet,CAF)肥胖大鼠的腸系膜脂肪含量,并增加其胰島素敏感性,改善血脂異常[94]。Pedret等[95]發(fā)現(xiàn)熱滅活的Ba8145顯著改善腹部肥胖指標、舒張壓和胰島素抵抗指數(shù)的機制可能與其促進腸道Akk豐度的增加有關(guān)。滅活的長雙歧桿菌也能改善小鼠腸道菌群組成,抑制HFD引起的小鼠肥胖[96]。腸道屏障通常會阻止來自微生物或飲食的LPS,腸道屏障的受損會導(dǎo)致肥胖動物或人類血漿LPS水平的長期升高[97]。長雙歧桿菌通過降低小鼠血液中的LPS水平,減少其向腸道的流入,以改變腸道菌群豐度,并促進GLP-1分泌,從而控制小鼠進食欲望來改善肥胖癥狀。Othman等[98]探究了攝入滅活長雙歧桿菌對肥胖糖尿病模型小鼠的影響,進一步證明長雙歧桿菌對體質(zhì)量的增加和總脂肪組織的積累有顯著的抑制作用。
3.1.3 乳桿菌屬
單磷酸腺苷(adenosine 5’-monophosphate,AMP)激活的蛋白激酶(AMP-activated protein kinase,AMPK)參與維持體內(nèi)脂類和膽固醇的細胞能量平衡和穩(wěn)態(tài)[99]。Lee等[100]證實口服植物乳桿菌KTCT13497BP(L-14)提取物可通過AMPK信號通路抑制小鼠前脂肪細胞(3T3-L1)和人骨髓間充質(zhì)干細胞(hBMMSCs)向成熟脂肪細胞分化。L-14提取物中的EPS通過TLR2和AMPK信號通路抑制脂肪形成,口服L-14提取物可改善體內(nèi)肥胖及肥胖相關(guān)疾病。Lim等[101]發(fā)現(xiàn)植物乳桿菌k8制備的后生元LAB-P可以激活能量調(diào)節(jié)因子AMPKα,AMPKα激活后通過刺激脂質(zhì)氧化、葡萄糖攝取和抑制脂質(zhì)合成發(fā)揮減肥降脂作用[102]。近年來也有利用植物乳桿菌L-137調(diào)節(jié)脂代謝以改善肥胖的研究,如熱滅活L-137可減輕大鼠脂肪組織肥大和炎癥、改善胰島素抵抗[103],臨床研究發(fā)現(xiàn)每日攝入熱滅活L-137能夠改善超重或健康成人的脂代謝[97]。除植物乳桿菌外,熱滅活副干酪乳桿菌N1115對長期HFD導(dǎo)致的脂代謝異常具有改善作用[104]。開菲爾乳酸菌后生元(postbiotic lactic bacteria,PLAB)[105]的細胞成分、表層蛋白和EPS可以修復(fù)HFD引起的生態(tài)失調(diào),改善以Akk為代表的肥胖特征性腸道微生物菌群豐度,其表層蛋白和EPS與葡萄籽粉的多酚類物質(zhì)結(jié)合,能夠協(xié)同改善HFD誘導(dǎo)的脂肪生成。過氧化物酶體增殖物激活受體(peroxisome proliferatorsactivated receptors,PPAR)的激活可改善脂質(zhì)代謝紊亂,研究人員從32 種食源性細菌中篩選出一株高PPARα/γ活性的噬淀粉乳桿菌CP1563,發(fā)現(xiàn)物理破碎后的菌株可以代替有機溶劑萃取來表達CP1563的體外活性[106]。
除上述幾種常見的改善肥胖后生元菌株,研究人員在酵母屬、芽孢桿菌屬、鏈球菌屬及糞桿菌屬[107-109]中也發(fā)現(xiàn)了可用于改善肥胖的后生元菌株。
3.2 后生元菌株代謝產(chǎn)物改善肥胖癥狀研究進展
SCFAs是指碳鏈中碳原子數(shù)小于6的有機脂肪酸,主要包括乙酸、丙酸、異丁酸、丁酸、異戊酸、戊酸[110]。SCFAs是人體腸道中最豐富的微生物代謝物之一[111],這些代謝產(chǎn)物在腸道菌群和宿主之間起中介作用。SCFAs參與體內(nèi)脂肪酸合成、脂肪酸氧化和脂肪分解的調(diào)節(jié)[110],作為后生元活性物質(zhì),其抵御肥胖作用通常涉及3 種機制——能量消耗、抑制食物攝入和調(diào)節(jié)脂質(zhì)代謝[93]。丙酸作為人體脂肪酸生成的抑制劑[112],可降低體質(zhì)量、收縮壓和空腹胰島素水平,并抑制HFD引起的肝臟大泡性脂肪變性、肝細胞肥大[19]。有研究發(fā)現(xiàn)運動后生成的血清乳酸可跨越腸上皮屏障進入小鼠腸道,在腸道菌群作用下代謝為丙酸,從而提高機體運動性能,增大能量消耗[113]。丁酸作為一種組蛋白去乙酰化酶抑制劑,能夠通過調(diào)節(jié)基因表達而調(diào)控相關(guān)細胞代謝[114]。動物實驗表明,攝入丁酸對脂肪組織的代謝和功能、能量和底物的代謝、胰島素敏感度和體質(zhì)量控制有積極影響[115]。Wang Li等[115]發(fā)現(xiàn)丁酸鹽可降低小鼠食欲,促進脂肪氧化,激活BAT。丙酸鹽和丁酸鹽在人體血液中的濃度通常較低,但它們可以通過激活激素和神經(jīng)系統(tǒng)間接發(fā)揮作用[116]。如丁酸鹽通過腸-腦神經(jīng)回路降低食欲并激活BAT[117],丙酸鹽、乙酸鹽也能夠以類似途徑對葡萄糖及脂質(zhì)代謝、食欲及能量消耗產(chǎn)生影響[118]。總體來說,微生物來源的SCFAs能夠通過腸道、中樞神經(jīng)系統(tǒng)、脂肪組織的互作減少食欲[119],SCFAs作為G蛋白偶聯(lián)受體GPR41和GPR4396的信號分子或內(nèi)源性配體,介導(dǎo)SCFAGPR41腸-腦神經(jīng)軸,并通過GPR依賴的方式刺激腸內(nèi)分泌GLP-1和肽YY(peptide YY,PYY),讓大腦產(chǎn)生飽腹感,從而減少食物攝入[111]。瘦素[120]作為飽腹激素,是脂質(zhì)代謝及相關(guān)代謝通路的重要成分,由脂肪組織合成,其合成和釋放也與GPR41的激活以及GLP-1和PYY的表達量增加有關(guān)。
膽汁酸是能量穩(wěn)態(tài)的外周和中樞調(diào)節(jié)因子,在全身脂質(zhì)代謝和葡萄糖代謝中發(fā)揮關(guān)鍵的調(diào)控作用[118],其通過膽汁酸在體內(nèi)的初級膽汁酸代謝和次級膽汁酸代謝來實現(xiàn)[121]。腸道菌群可以生成多種次級膽汁酸,其中脫氧膽酸(deoxycholic acid,DCA)和石膽酸(lithocholic acid,LCA)是腸道菌群代謝最豐富的次級膽汁酸,其通過G蛋白偶聯(lián)受體TGR5調(diào)節(jié)宿主能量穩(wěn)態(tài)和代謝[122]。腸道中的一些特定細菌還能通過7α-脫羥基作用將膽酸和鵝去氧膽酸等初級膽汁酸分別轉(zhuǎn)化為DCA和LCA[123],同時DCA、LCA及其衍生物是膽汁酸池再循環(huán)的主要成分,而膽汁酸池的大小、組成或其生物學作用的改變,也可能會使機體平衡向肥胖偏移[124]。此外,12α-羥基膽汁酸是另一種影響膽汁酸池組成的次級膽汁酸,膽汁酸代謝酶Cyp8b1是產(chǎn)生12α-羥基膽汁酸必需的酶,研究發(fā)現(xiàn)Cyp8b1基因敲除小鼠的膽汁酸水平正常,但膽汁酸組成顯著改變,因此影響了小鼠進食情況[125]。
革蘭氏陽性細菌和革蘭氏陰性細菌中有一些微生物可以促進色氨酸代謝產(chǎn)生吲哚及各類吲哚衍生物,如吲哚乙酸、色胺、吲哚醛等[13]。有研究人員在HFD誘導(dǎo)的小鼠實驗中發(fā)現(xiàn),吲哚能夠有效預(yù)防肥胖,這是由于腸道吲哚代謝物作為一種信號分子,可以誘導(dǎo)腸內(nèi)分泌L細胞釋放GLP-1,從而抑制食欲、促進胰島素分泌、減緩胃排空[126],這一結(jié)論在吲哚丙酸大鼠實驗中同樣得到了證實[127]。同樣地,多形擬桿菌代謝物色胺能夠激活腸上皮G蛋白偶聯(lián)受體,調(diào)節(jié)小鼠脂代謝[128]。另一腸道菌群(尤其是乳桿菌屬)的吲哚類代謝物3-吲哚乙酸與LPS可分別作用于孕烷X受體及TLR4,促進調(diào)節(jié)性B細胞IL-35+B在脂肪組織中的累積,從而抑制HFD誘導(dǎo)的小鼠肥胖[129]。Virtue等[130]通過動物實驗發(fā)現(xiàn)來自腸道微生物菌群的色氨酸代謝物能夠控制小鼠WAT中RNAmir-181的表達,從而調(diào)節(jié)其能量消耗和胰島素敏感性。一項納入63 名正常和肥胖受試者的人體研究對菌群代謝產(chǎn)物與大腦獎勵網(wǎng)絡(luò)的功能和連接關(guān)系進行分析,發(fā)現(xiàn)腸道菌群吲哚代謝物(吲哚、吲哚乙酸和甲基吲哚)含量的增加與受試者大腦中杏仁核、伏隔核和前腦島等區(qū)域的功能和連接增強相關(guān),且吲哚和甲基吲哚分別與BMI和食物上癮評分呈正相關(guān),這表明腸道菌群衍生的吲哚代謝物可能作用于大腦的獎賞網(wǎng)絡(luò),從而影響食物上癮和肥胖[131]。此外,通過臨床樣本分析以及小鼠和細胞實驗發(fā)現(xiàn),吲哚含量與肥胖相關(guān)的NAFLD存在負相關(guān)性,補充吲哚能減輕小鼠飲食誘導(dǎo)的NAFLD,減少肝臟脂肪含量[132]。
細胞外囊泡(extracellular vehicles,EVs)是微生物釋放的球形結(jié)構(gòu),攜帶各種生物活性成分。越來越多的證據(jù)表明,腸道微生物菌群衍生的EVs可能是微生物-宿主跨領(lǐng)域交流的相關(guān)媒介,在腸道微生物菌群介導(dǎo)的腸道穩(wěn)態(tài)中發(fā)揮作用,并最終參與一些代謝性疾病的調(diào)控。一些研究已經(jīng)證實EVs可以改善肥胖癥,并且能夠輕松穿過黏液層與宿主相互作用,以降低敗血癥感染風險[109]。Chelakkot等[133]探究了EVs與改善HFD誘導(dǎo)糖尿病小鼠腸道屏障完整性和代謝狀況之間的直接關(guān)系,發(fā)現(xiàn)口服Akk的EVs能夠降低小鼠腸道屏障通透性,減少體質(zhì)量增加,并改善葡萄糖耐量。腸道微生物還可以代謝一些具有生物活性的脂質(zhì),常作為后生元代謝物發(fā)揮作用。10-羥基-順式-12-十八碳稀酸(10-hydroxy-cis-12-octadecenoic acid,HYA)是由微生物代謝產(chǎn)生的初始中間共軛亞油酸,HYA能夠激活GPR40和GPR120并促進GLP-1的釋放,直接調(diào)節(jié)葡萄糖穩(wěn)態(tài),并通過G蛋白偶聯(lián)受體家族的EP3受體促進腸道蠕動,從而抑制脂質(zhì)吸收[134]。一些乳酸菌代謝物ω-6多不飽和脂肪酸可以在不引起花生四烯酸介導(dǎo)的脂肪炎和通過游離脂肪酸受體改善代謝條件的情況下減輕HFD誘導(dǎo)的小鼠肥胖[134]。其他代謝物如擬桿菌產(chǎn)琥珀酸可以通過激活腸道糖異生作用降低血糖,從而改善小鼠肥胖和代謝功能障礙[135]。普氏桿菌Prevotella copri可改善葡萄糖代謝與其產(chǎn)生的琥珀酸相關(guān)[136]。除此之外,部分腸道細菌的神經(jīng)遞質(zhì)類代謝產(chǎn)物神經(jīng)酰胺還能夠通過分解代謝來介導(dǎo)環(huán)境低溫誘導(dǎo)機體的產(chǎn)熱活動[137]。
3.3 其他形式后生元改善肥胖癥狀研究進展
目前對抵御肥胖后生元的研究多集中在菌株發(fā)掘和代謝產(chǎn)物方面,而構(gòu)成菌體細胞本身的成分如磷壁酸、肽聚糖、LPS中還鮮有發(fā)現(xiàn)具有確切減肥降脂功效的后生元,其他形式的后生元如細胞裂解物多見于調(diào)節(jié)機體免疫與改善炎癥方面[138-140],一些以無細胞上清液形式存在的后生元被發(fā)現(xiàn)具備改善肥胖的作用。無細胞上清液中含有許多低或高分子質(zhì)量的生物分子和活性代謝物,能夠?qū)C體健康產(chǎn)生有益的作用,多發(fā)現(xiàn)于乳酸菌或酵母菌上清液中[141],鼠李糖乳桿菌GG上清液能夠減少高脂肪/高果糖飲食加間歇性缺氧暴露小鼠的體質(zhì)量增加和肝臟脂肪的沉積,減少脂肪含量,減輕胰島素抵抗和肝臟損傷,大幅減少新脂肪生成,增強脂質(zhì)β-氧化,增加能量消耗[142]。胃饑餓素是由胃部細胞產(chǎn)生的多肽,被稱為“饑餓荷爾蒙”,負責提高食欲并促進脂肪囤積,其受體GHSR-1a遍布人體的腸道和大腦,在維持能量平衡以及對食物攝入動機和情緒的中樞調(diào)節(jié)方面起著關(guān)鍵作用。有研究發(fā)現(xiàn)制備雙歧桿菌(長雙歧桿菌APC1472和短雙歧桿菌APC6331)和乳酸桿菌(鼠李糖乳桿菌DPC6118和加氏乳桿菌DPC6106)上清液能夠調(diào)控GHSR-1a減弱胃饑餓素介導(dǎo)的信號傳導(dǎo)[143],這為上清液形式抗肥胖后生元的功效提供了有力證據(jù)。
4 結(jié) 語
肥胖作為全球性疾病,已成為世界上最普遍的公共健康問題之一,與之相關(guān)的代謝紊亂疾病發(fā)病率在過去的幾十年里一直在穩(wěn)步上升,肥胖的發(fā)生與人體能量失衡和機體選擇性代謝密切相關(guān),但有關(guān)肥胖的病理生理學機制仍存在爭議且尚未完全明確。后生元是不具備生命活性的混合物,其種類豐富、形式多樣,可以通過不同途徑發(fā)揮對機體的有益作用,其不僅可應(yīng)用于食品、保健品等領(lǐng)域,也有越來越多的研究探索其在臨床治療和預(yù)防疾病方面的功效。近年來大量體外和體內(nèi)模型被用來評估后生元在治療、緩解肥胖癥狀方面所具有的潛在益生功能及作用機制,發(fā)現(xiàn)其菌株主要為Akk、雙歧桿菌屬及乳桿菌屬,且后生元在形式上呈現(xiàn)出多樣化特點,揭示后生元在肥胖的預(yù)防和疾病治療中有著良好的發(fā)展前景。
當前肥胖依舊是普遍存在的一種慢性疾病,是多種代謝紊亂病的主要誘發(fā)因素之一,嚴重危害著全球公共衛(wèi)生健康,同時肥胖也是WHO一直非常關(guān)注的疾病之一,控制體質(zhì)量的方法和預(yù)防肥胖的相關(guān)措施一直在隨著科技的進步與研究的深入而多樣化推進。目前,后生元由于無生命特征、菌體成分及其活性成分可以高密度集中、安全系數(shù)高、質(zhì)量穩(wěn)定且可與抗生素同時使用等優(yōu)勢,已被廣泛應(yīng)用于食品、藥品等領(lǐng)域,過去20 年間對肥胖的研究和臨床試驗數(shù)量顯著增加,盡管后生元的部分作用已得到研究證實,但其對人體抗肥胖的作用機制還不完全明確;因此,有必要通過促進生物機動性(后生元)的綜合治療策略來優(yōu)化相關(guān)肥胖綜合征所涉及的不同治療方法。后生元有望成為代替活菌或其他藥物治療肥胖的有效手段。
猜你喜歡
興趣閱讀·興趣作文與閱讀(低年級)(2025年8期)2025-08-18 00:00:00
東方少年·布老虎畫刊(2023年8期)2023-08-01 15:45:12
科學大眾(2021年6期)2021-07-20 07:42:44
科學(2020年3期)2020-11-26 08:18:30
學苑創(chuàng)造·A版(2020年9期)2020-10-13 09:41:02
娃娃樂園·綜合智能(2019年3期)2019-04-03 09:17:36
中成藥(2018年2期)2018-05-09 07:19:34
小學生學習指導(dǎo)(低年級)(2017年10期)2017-10-10 01:00:05
湖北師范大學學報(自然科學版)(2015年2期)2016-01-10 08:41:55
云南中醫(yī)學院學報(2014年3期)2014-07-31 18:57:34
