認知地圖及其內在機制*
2023-10-09 02:53:48吳文雅
心理科學進展 2023年10期
吳文雅 王 亮
·研究前沿(Regular Articles)·
認知地圖及其內在機制*
吳文雅 王 亮
(中國科學院心理健康重點實驗室(中國科學院心理研究所), 北京 100101) (中國科學院大學心理學系, 北京 100049)
空間導航對于人和動物的生存而言都十分重要, 有效的空間表征或認知地圖是空間導航的基礎。認知地圖的典型屬性包括選擇性、靈活性以及層級性, 海馬、場景選擇區域以及前額葉等多個腦區都參與認知地圖的構建。認知地圖的表征形式存在歐式地圖和拓撲圖兩種理論, 但各自單獨都不能全面解釋實際導航中的行為表現, 因此有研究者提出了標簽圖等理論試圖調和二者間的矛盾。未來研究還需要關注在認知地圖構建過程中層級性的變化, 空間范疇的擴展, 以及認知地圖假說的局限。
認知地圖, 歐式地圖, 拓撲圖, 層級性, 空間表征
1 引言
對于人類和其他生活在復雜環境中的動物(如蝙蝠)而言, 空間導航是一項重要的認知功能(Genzel et al., 2018; Goldshtein et al., 2022)。高效的空間導航需要對空間信息進行有效的心理表征(mental representation), 這種表征是對外界信息和客觀知識的內部主觀形式, 不同于常見的外部客觀形式, 如制圖師制作的平面地圖以及目前被大眾廣泛使用的智能手機導航軟件(如高德地圖)。空間知識的內部表征對于人們在熟悉環境下的導航行為至關重要, 可以通過調用已存儲的空間知識靈活設計出行路線(Harten et al., 2020), 進而節約導航時間。有研究表明, 長期依賴于導航軟件可能會導致空間記憶和導航能力的退化(Dahmani & Bohbot, 2020), 因此空間知識的內部表征機制具有重要的理論意義和研究價值。
由于空間知識的內部表征能夠為路線規劃、捷徑選擇等高級認知行為提供幫助, 在功能上類似于地理學意義上制作的地圖, 因此也被稱為認知地圖(cognitive map)。認知地圖這一概念最早由Tolman (1948)提出, Tolman發現隨著練習次數增加大鼠在迷宮任務中的空間導航行為會逐漸優化, 能夠選擇學習階段未經歷過的新異捷徑抵達目標, 這表明大鼠并非僅僅建立刺激?反應的簡單聯結序列, 而是理解了環境的整體空間結構, 主動選擇重要線索從而建構出獨立于自身而存在的環境全景地圖(allocentric comprehensive map)。
盡管Tolman提出的認知地圖概念非常粗糙, 僅僅在功能上闡述了動物超越親身經歷而建立起環境全局認知的必要性, 并沒有對認知地圖的諸多細節做出詳細說明和驗證, 但是仍然催生了后續空間導航研究的大量成果, 在理論和實驗研究方面都引發了突破性的進展。此外, 雖然認知地圖概念誕生于心理學領域, 但卻因其深刻性和普適性而吸引了其他學科的研究興趣, 其內涵也在不斷擴展, 不再局限于認知心理學和認知神經科學, 而是逐漸延伸到計算機科學與工程學、經濟管理學、行為地理學等諸多領域(阮曉鋼等, 2021; 何詩, 陰劼, 2022)。
鑒于目前對于認知地圖的構建規則、精確程度、表征形式等方面的了解仍然有限, 并且不同研究者持有迥異的見解, 本研究通過文獻回顧對認知神經科學領域的認知地圖相關理論和實驗研究進行梳理, 以期為多學科的認知地圖研究者提供啟發。本研究首先基于行為現象和功能視角歸納認知地圖所表現出的典型屬性, 進一步闡述參與認知地圖構建的相關腦區及其作用機制, 隨后總結在認知地圖存在形式方面既有的理論爭議, 最后對認知地圖這一思想進行反思并展望未來空間表征和認知地圖領域的重要問題和研究方向。
2 認知地圖的典型屬性
認知地圖并非對空間知識進行完全保真的復制, 而是具有突出的心理屬性, 具體表現為根據任務需求對不同空間信息進行選擇性的加工和存儲, 這種片面的信息加工進一步導致了扭曲性的存在。其次, 為了應對復雜多變的環境, 空間表征還具有靈活性, 對同一信息容忍表征形式上的冗余性。此外, 由于可導航空間通常具有嵌套結構, 這類嵌套空間的認知地圖會表現出層級性, 不過隨著在嵌套環境中的導航經驗增加, 認知地圖的層級性也會下降, 逐漸變得連貫。
2.1 選擇性和扭曲性
認知資源是有限的, 所以空間表征在內容上需要具有選擇性, 從而提高認知的經濟性。以往心理學、經濟學以及人工智能等多學科的標準觀點是人對包括導航在內的任務形成完整而固定的表征, 并使用啟發式來規劃后續行為, 然而Ho等人(2022)發現針對具體問題人們會靈活地對環境形成特定的簡化表征, 以此來平衡任務表征的復雜度與實用性。
雖然位置、路徑關系、方向等多方面的信息都可以歸入空間知識的范疇, 但是在具體的實用情境下, 認知地圖的內容可能只涵蓋其中一部分。正如Borges (1971)在《論科學的精確性》這篇科幻小說中所揭示的, 當制作的地圖詳盡地包含了真實地理環境中的所有信息時, 這幅巨大的地圖實際上卻是無用的。不加選擇地表征所有信息, 既浪費認知資源, 又無法高效地解決特定問題。比如在一座城市旅行時, 交通工具的種類決定了合適的地圖種類, 騎行需要非機動車道的地圖, 而自駕游則需要機動車道的地圖, 顯然衛星拍攝的城市詳細地圖則是無法直接使用的。
選擇性的空間表征源于功能需求, 而不同的生存環境(如文化環境和地理環境)為導航者提出了不同的表征需求。在地圖繪制任務中, 被試會按照其所住城市的特有屬性去組織手繪地圖, 并且個人的路線使用也會在地圖上突出反映出來, 表明空間表征具有較強的社會功能意義(Appleyard, 1970)。一項納入多國家樣本的大規模研究發現, 人們更擅長在與成長環境具有相似拓撲特征的環境中完成導航任務, 城市的街道網絡設計會對居民的空間表征傾向及導航能力產生影響(Coutrot et al., 2022)。以上發現表明, 認知地圖的焦點會受到日用需求的塑造, 處于不同環境下的人所選擇性表征的內容也不同。
另一方面, 有選擇地表征就意味著所表征的信息是有局限的, 即相比于真實信息, 可能會存在一定的扭曲, 這種扭曲可以視為表征為了發揮特定的功能而不得不付出的代價。路徑距離估計會受到路徑上位置點數量的影響, 在控制實際距離的情況下, 位置點數量越多被試的距離估計會越大(Thorndyke, 1981)。靠近城市中心的路徑長度相比于外周路徑會被明顯高估, 角度估計也表現出趨向于90°的偏好(Byrne, 1979)。語言學信息也能夠解釋大尺度地理學地圖表征任務中觀察到的特定扭曲, 表明自然語言經歷會參與編碼和形成認知地圖(Gatti et al., 2022)。
由此可知, 作為對環境簡化而有目的的刻畫, 實用地圖必須以犧牲某些細節為代價來突出所關注的細節。不存在完全精確的實用地圖, 所有地圖都必然存在一定程度的扭曲, 扭曲的情況視其所發揮的功能而定(Hartley, 2017)。同樣, 也不存在完全精確的認知地圖, 空間表征的扭曲也伴隨特定功能而存在(Longo, 2021)。
2.2 靈活性和冗余性
靈活地運用已習得的知識對于動物應對復雜的外界環境和不可預知的挑戰十分重要。動物的生存環境并非一成不變, 比如原來通行的路徑被障礙物阻斷、原有的食物資源不知去向等, 高效導航的重要表現之一是應對環境線索和結構動態變化的能力(Kabadayi et al., 2018), 而認知地圖的靈活性可能是這種能力的基礎。
早在1948年研究大鼠在復雜迷宮中的繞路和捷徑選擇時, Tolman就發現大鼠的行為表現出很強的靈活性, 而不是簡單的“刺激?反應”序列。已有研究發現靈活的認知地圖和空間導航需要心理重放(mental replay), 并且壓力會損害靈活性(Epsztein, 2022; Brown et al., 2020)。
在靈活性的背后是空間表征和認知地圖形式的冗余性, 為了能夠支持靈活的導航行為, 相同的空間信息可能需要同時以多種形式加以組織和存儲(Yousif, 2022)。相同的信息或內容在不同條件或要求下的最佳呈現方式可能有很大差異, 比如電子文件的多種格式屬性不同但都有其獨特功能(PDF文件便于查看, 而Word文檔便于修改等)、問題的表述形式也會對解決問題的效率產生極大影響。同理, 對于同樣的空間信息, 最佳的認知地圖形式會隨具體的功能需求不同而不同, 因此大腦需要以多重格式冗余地表征信息以備調用。
2.3 層級性和連貫性
可導航空間往往是嵌套的環境, 包含不同的子區域, 如校園的不同建筑以及每個建筑內的不同房間, 導航系統對不同局部區域的表征可能是彼此分離的, 要建立對環境整體的認知地圖, 就需要整合通過離散學習獲得的不同局部區域的信息, 以合理的方式加以組織。不同局部區域的信息之所以是分離的, 是因為可導航空間包括單視點空間(vista space)和環境空間(environmental space)兩類(Meilinger et al., 2016), 前者從同一位置可以統覽全局空間的屬性, 如臥室、曠場(圖1a); 后者由于存在障礙物遮擋所以需要導航者在其間穿梭運動并整合在每個單視點空間中學習到的信息, 如街區、迷宮(圖1b)。
在嵌套的環境空間內(圖1c), 不同局部區域的信息可能是以層級化(hierarchical)的方式加以組織的。McNamara (1986)發現空間啟動、方向判斷以及歐氏距離估計任務表現都會受到不同位置是否在相同區域的影響。在分段化的平行走廊中人的空間記憶會按照每個走廊的局部坐標系統進行組織, 表明對不同局部區域的表征是分離的(Meilinger et al., 2016)。此外, 人在進行空間關系判斷時所表現出的扭曲支持了認知地圖具有層級性(Stevens & Coupe, 1978)。不過層級性并非持續存在, 不同局部區域可能會隨著學習逐漸發生整合, 形成統一連貫的整體。早期就有研究顯示, 不同區域的共同地標可以促進新的局部信息整合到已有的空間知識中(Golledge et al., 1993)。
3 認知地圖的神經機制
認知地圖神經機制研究最早關注的是頂葉皮層, 頂葉受損的患者會表現出空間導航能力的減弱甚至喪失。之后位置細胞的發現讓研究者們聚焦于海馬結構(hippocampus), 并且進一步發現了頭朝向細胞以及內嗅皮層(entorhinal cortex)的網格細胞和邊界細胞, 表明海馬?內嗅系統是認知地圖的關鍵區域。此外, 內側前額葉、眶額葉以及場景選擇性區域也在認知地圖的形成過程中發揮重要作用。
3.1 海馬?內嗅系統
O’ Keefe和Dostrovsky (1971)首次在嚙齒類動物的海馬中記錄到位置細胞(place cell)的存在, 這些細胞在動物處于特定位置時才會發放(圖2a), 由此表明海馬能夠編碼空間位置。除了位置元素以外, 后續研究陸續發現了海馬結構中編碼方向、環境邊界等其他空間信息的多種類型的細胞, 如網格細胞(grid cell)、頭朝向細胞(head direction cell)、邊界細胞(border cell)等(Hafting et al., 2005; Taube et al., 1990; Lever et al., 2009)。其中內嗅皮層的網格細胞在動物處于平鋪環境的等邊三角形頂點處發放(圖2b), 并且表現出六重的旋轉對稱性(sixfold symmetry) (Hafting et al., 2005; Doeller et al., 2010), 即每個網格細胞發放的位置點在曠場中排列成周期性的正六邊形(也可視為6個等邊三角形的組合), 不僅為空間位置提供背景標度, 也能夠計算從當前位置到導航目標的方向(Bellmund et al., 2016)。

圖1 單視點空間(a)、環境空間(b)和嵌套的環境空間(c)示意圖。在單視點空間中, 從一個位置或視點可以總覽空間內的所有物體或位置。在環境空間中, 由于存在遮擋, 需要在障礙物之間不斷移動和穿梭, 分別觀察和學習每個單視點空間中的信息, 隨后整合形成全局空間認知。在嵌套空間中, 每個區域可能包含多個子區域, 如圖中三種顏色背景代表三個不同區域, 每個區域內部又有由彩色線段分隔開的多個子區(類似于不同小區內部進一步分成不同單元)。彩圖見電子版。

圖2 海馬位置細胞(a)和內嗅皮層網格細胞(b)的放電模式示意圖。黑色線代表動物在方形區域內的奔跑軌跡, 紅色點代表細胞發放(spikes)的峰值位置, 疊加在運動軌跡上。位置細胞只有單一的放電位置, 而網格細胞的放電位置構成了周期性的六邊形(或6個正三角形) (改編自Moser et al., 2008)。
海馬?內嗅系統對于歐式度量信息和拓撲連接關系的表征都十分重要。神經影像學研究發現, 海馬和內嗅皮層能夠編碼位置間的歐氏距離和路徑距離, 并且海馬長軸還存在功能分區, 其中前部與歐式距離相關, 后部則與路徑距離相關(Morgan et al., 2011; Howard et al., 2014)。海馬活動對歐式距離的敏感源于內嗅皮層(entorhinal cortex, ERC)的信息輸入, 內嗅皮層網格細胞的多峰發放域為空間表征提供了規則的網格背景(Hafting et al., 2005), 不過內嗅皮層的信息輸入對海馬空間表征的重新組織(或稱重映射, remapping)并不是必要的(Schlesiger et al., 2018)。海馬結構也表征環境的拓撲信息, 在受約束的、由不同通道相連構成的迷宮中, 嚙齒類動物的海馬位置域由相對于迷宮拓撲結構的位置決定, 而非歐式空間中的絕對位置(Dabaghian et al., 2014), 此外在休息期間位置細胞的序列化重激活也反映了迷宮的拓撲結構(Wu & Foster, 2014)。在虛擬街道中導航時人類海馬后部信號與未來可能經歷的路徑連接邊數量相關, 而海馬前部信號則與環境整體的拓撲學性質相關(Javadi et al., 2017)。
對環境尺度的差異化表征主要依賴位置細胞發放域在海馬長軸上的分離(Evensmoen et al., 2013; Brunec et al., 2018)以及網格細胞發放域在內嗅皮層腹背軸上的分離(Stensola et al., 2012), 其中海馬前部表征大尺度全局環境的粗糙信息, 而海馬后部則支持局部環境的精細粒度表征; 對于內嗅皮層, 越靠近腹側的細胞具有更大的網格域, 因而表征更大尺度環境中的粗糙信息, 反之越靠近背側的細胞則更精細地表征小尺度環境中的空間信息。
環境幾何學特征(如邊界形狀)會對網格細胞的位置域產生影響, 將方形環境更改成梯形會高度扭曲嚙齒類動物網格細胞原有的規則發放模式(Chen et al., 2015), 網格模式的下降可能與認知地圖發生扭曲有關。對人類被試的虛擬現實研究也表明, 相比方形環境, 在梯形環境中人的位置記憶準確率會下降, 這種影響在梯形的狹窄部分尤為明顯(Bellmund et al., 2020)。因此, 內嗅皮層網格細胞所提供的幾何標度變形可能是認知地圖扭曲性的主要原因。
海馬結構參與實現認知地圖的靈活性。Widloski和Foster (2022)在迷宮任務中記錄大鼠的海馬細胞活動, 通過改變障礙物的分布不斷變化迷宮的結構和食物位置, 發現大鼠可以靈活調整覓食路線, 并且在休息期間海馬細胞的“重放/重激活(replay/reactivation)”能夠預測未來路線和障礙物規避情況; 在此過程中大部分位置細胞的位置域保持穩定, 只有少部分會隨著障礙物分布變化而變化, 而這部分“不穩定”細胞可能與大鼠靈活應對空間布局變化的行為表現緊密相關。
海馬?內嗅系統在認知地圖的層級性組織中也發揮重要作用。環境中不同的局部區域在新學習時是分離表征的, 人類導航的行為研究和嚙齒類動物導航的電生理研究均支持這一點, 例如人類的空間記憶會受到環境中障礙物的影響, 依據障礙物分割環境的形狀來形成局部的參考系(Meilinger et al., 2016); 嚙齒類動物的內嗅網格細胞和海馬位置細胞會對局部區域分別創建獨立的局部地圖, 具體表現為網格域和位置域局限在局部環境內(Alme et al., 2014)。不過, 長時間在多隔間的空間內導航也會引起隔間邊界影響的減弱甚至消失, 具體表現為網格細胞的發放域擴展到全局空間(Carpenter et al., 2015)。盡管最初學習時嚙齒類動物和人都傾向于將局部表征分離, 然而在具有多個局部區域的環境中經歷長時間、豐富的穿梭體驗之后, 嚙齒類動物的網格細胞發放域會快速移動, 從之前離散的局部網格域轉變為連續的全局網格域, 通過這種方式來逐步整合局部區域表征, 最終形成全局一致的認知地圖(Wernle et al., 2018), 在人類中則沒有發現這種現象, 人類在多區域環境下的豐富導航經驗并不一定會導致局部表征的整合(Zhao, 2018), 但也有可能是研究人類對象的實驗范式不同導致的。
3.2 場景選擇區域
視覺場景是小尺度局部環境表征的主要部分, 正確識別場景或位置對人們的日常生活具有重要意義, 其表征需要皮層網絡的參與。視覺場景的加工系統并不是表征可導航空間的海馬?內嗅系統, 而是場景選擇區域(scene-selective regions)。此區域在觀看場景時發生強烈反應, 包括旁海馬位置區(parahippocampal place area, PPA)、枕葉位置區(occipital place area, OPA)和壓后皮層復合體(retrosplenial complex, RSC)這三個功能子區域(Dilks et al., 2022)。
在功能上, 主流觀點認為這三個子區域都直接參與空間導航。OPA表征局部空間布局、環境約束邊界和邊界距離(Henriksson et al., 2019; Julian et al., 2016; Park & Park, 2020); PPA則參與加工場景的視覺和幾何特征等(Marchette et al., 2015)。而RSC負責整合來自上面兩個區域的局部表征, 形成統一的全局表征(Epstein et al., 2007), 功能核磁研究發現路徑位置點的依序出現會引發RSC更強烈的活動(Schinazi & Epstein, 2010), 表明RSC的活動與路徑拓撲結構有關。
總體而言, 場景選擇區域參與“縫合”多個離散的單視點空間, 為導航者的空間表征編織一個360°全景視圖, 包括當前視野內外的空間信息, 此外當前場景的信息還會啟動全景環境內相關聯的表征(Robertson et al., 2016)。通過這種方式, 場景選擇區域的空間表征實現了短暫的空間知覺與相對持久的空間記憶之間的動態交互, 促進全局認知地圖的形成。也有研究者將上述“縫合”過程稱為“映射”, 即壓后皮層根據單視點空間的初級感覺信息提取出認知地圖, 再根據識別出的環境地標對認知地圖的布局進行調整和實時更新, 從而完成認知地圖與現實環境的映射, 進而保持認知地圖與現實空間布局的一致性(梁群君等, 2022)。
3.3 前額葉
以往研究一般認為, 認知地圖和空間導航的核心腦網絡由海馬?內嗅系統和場景選擇區域共同構成, 但是卻忽略了前額葉(prefrontal cortex, PFC)在適應環境變化、追蹤和提前規劃以規避風險等方面的關鍵作用, 可能有如下三個原因導致大部分研究者并不重視前額葉對空間表征的重要性。首先, 前額葉損傷一般會嚴重損傷執行功能(Shallice & Burgess, 1991), 這自然會導致大多數任務都表現更差, 而不特異于空間任務; 其次, 早期研究中損毀大鼠的內側前額葉似乎并不影響在水迷宮等導航任務中的表現(Lacroix et al., 2002; Sloan et al., 2006); 最后, 前額葉存在位置細胞和網格細胞相似編碼的實驗證據總體上更少并且出現也相對較晚(Park et al., 2021; Jacobs et al., 2013)。
已有研究發現前額葉的不同子區域表現出功能上的差異。背外側前額葉(dorsolateral PFC, dlPFC)和腹外側前額葉(ventrolateral PFC, vlPFC)在由障礙物導致的繞路過程中出現活動, 因此可能參與路線的重新規劃(Javadi et al., 2019)。內側前額葉(mPFC)與自我中心的空間表征有關, 在導航者記住自我中心的目標位置時能夠編碼基于物體間相對關系的認知地圖(Zhang & Naya, 2020), 也能產生有效行為序列的重激活(Kaefer et al., 2020), 參與對環境空間拓撲學上有組織的表征(Sauer et al., 2022), 背內側前額葉還會參與分離任務相關和任務無關的空間背景(Mahmoodi et al., 2023)。眶額葉(orbitofrontal cortex, OFC)表征任務的狀態空間(Schuck et al., 2016) (如價值空間、特征空間), 把對環境的空間表征與行為的價值整合起來, 形成可能的決策地圖, 也參與相同空間背景內表征內容的壓縮(Muhle-Karbe et al., 2023)。此外, 背側前扣帶回(dorsal anterior cingulate cortex, dACC)參與路徑網絡的層級編碼(Anggraini et al., 2018), 并且能抑制錯誤的擇路傾向(Javadi et al., 2019)。
鑒于前額葉空間表征內容的多樣性, Patai和Spiers (2021)總結指出, 前額葉的不同區域可能分別在導航不同階段發揮作用:在導航的開始, 內側前額葉回憶出導航目標的位置, 交由背內側前額葉進行路線規劃, 隨后若需繞路則由背側前扣帶回進行可選路線的搜索, 接著外側前額葉會進行路線的重新規劃并避免錯誤的捷徑, 在此過程中OFC會選擇特定路徑并推理其潛在結果, 內側前額葉則表征路徑距離的遠近, 當超出目標范圍時背側前扣帶回會協助導航者原路返回。
綜上可知, 作為高級腦區, 前額葉的主要角色是認知地圖的運用者和操作者, 接收來自海馬的空間信息(Nardin et al., 2021), 隨后靈活運用于規劃、推導和預測, 進而實現高效的空間導航。
3.4 海馬?皮層共同作用
海馬結構、場景選擇區域以及前額葉在認知地圖的構建中各自發揮獨特的功能, 但是認知地圖的某些高級屬性需要多個腦區的共同作用, 而無法僅由單個腦區就能實現。多個腦區之間的共同作用可能是協同交互, 也可能是平行競爭。
認知地圖的選擇性可能需要海馬結構與眶額葉的協同交互。在近期一項結合fMRI和計算建模的研究中, 由于刺激被嵌套在多重關系結構中, 被試在學習階段同時會形成環境刺激的空間位置關系和預測性關系(即刺激間的轉移關系)兩種認知地圖, 然而在隨后的選擇任務中僅有空間位置關系會決定獎賞額度, 建模結果表明被試的選擇會越來越受到空間位置關系的影響, 行為建模得出的空間權重逐漸增大, 而預測性權重則逐漸減小, 由此表明任務需求會驅動空間位置認知地圖的選擇性強化和預測性認知地圖的選擇性弱化。而fMRI結果則表明空間權重變化的敏感腦區主要在海馬, 而眶額葉則根據獎賞價值驅動海馬更新對當前任務關鍵的認知地圖(Garvert et al., 2023)。
然而, 近期另一項大鼠單細胞記錄研究則提出了相反的觀點, 認為海馬結構和眶額葉雖然都參與認知地圖的形成, 但二者之間并不是上述的簡單前饋關系(即眶額葉根據任務需求驅動海馬更新認知地圖), 而是平行運作的, 甚至還存在競爭關系, 二者分別提取不同的特征從而形成不同的認知地圖, 其中眶額葉傾向于形成反映當前任務相關性和動機性目標的表征, 而海馬的信息輸出則會阻礙眶額葉中的“圖式細胞(schema cells)”參與構建新表征(Zong et al., 2023)。即海馬更傾向于“守舊”, 提取既有的任務圖式, 抑制新圖式的形成; 而眶額葉則傾向于動態反映任務相關的目標, 形成或靈活選擇新的認知地圖。因此, 未來研究需要進一步探索眶額葉和海馬在實現認知地圖的選擇性上分別發揮什么作用、二者之間的關系以及是否還有其他腦區參與。
海馬和皮層的信息交互對認知地圖構建過程中的分段環境表征也十分重要。有證據表明, 在有多個隔間的環境中導航時, 跨越邊界會引起海馬活動出現峰值, 而在此之前會先出現皮層活動模式的變化(Baldassano, 2017)。在虛擬現實的三維多室建筑內, 相比于不同房間, 當被試在同一房間內進行空間判斷時反應更快, 表現出同一水平空間表征的行為啟動效應, 而fMRI結果表明, 三維空間信息的分層編碼受到海馬和皮層的協同作用, 其中左側海馬前部表征房間內的局部信息, 而壓后皮層、旁海馬皮層和海馬后部則表征更廣泛建筑內的房間信息(Kim & Maguire, 2018)。此外, 位于紋狀體的尾狀核也會和海馬一同與前額葉發生交互, 參與編碼環境的轉移結構, 共同支持靈活導航(Brown et al., 2012; Gahnstrom & Spiers, 2020)。
由于多個腦區共同參與認知地圖的構建, 所以從神經結構網絡的角度整合這些腦區的功能有助于更深刻地揭示認知地圖的本質。認知地圖的構建需要海馬和皮層共同參與形成的三條功能神經環路(王琳, 王亮, 2017), 其中第一條功能環路負責將外部信息(包括空間信息和非空間信息)從皮層投射到海馬進而形成內在表征; 第二條功能環路負責將海馬?內嗅系統的內在表征經由壓后皮質投射到后頂葉, 指導前額葉和輔助運動區, 進而轉化為行為上的動作輸出; 第三條功能環路負責目標獎賞, 由海馬和腹側紋狀體聯合表征。
4 認知地圖的存在形式
對于認知地圖以什么形式存在, 目前的理論爭議主要包括歐式地圖(Euclidean map)和拓撲圖(topological graph)兩種假設, 二者分別位于表征精確度的兩個極端, 歐式地圖要求完全精確地表征空間位置坐標, 而拓撲圖則不包含任何度量信息, 在解釋真實導航行為上二者都存在一定缺陷。因此, 有研究者試圖調和這兩種理論, 將歐式度量信息和拓撲結構都融合在認知地圖中。
4.1 歐式地圖假設
在Tolman認知地圖思想的基礎上, O’ Keefe和Nadel (1978)進一步提出認知地圖采取絕對、統一的歐式度量結構, 這種全局一致的歐式認知地圖有助于實現靈活的空間行為。支持這種提議的主要生理學依據是在海馬中發現的位置細胞和在內嗅皮層中發現的網格細胞, 如前所述, 位置細胞只在動物處于環境中的特定位置才會發放(O’ Keefe & Dostrovsky, 1971), 而網格細胞周期性地在動物處于平鋪環境的等邊三角形頂點處發放(Hafting et al., 2005; Doeller et al., 2010), 這兩種細胞共同實現對空間環境的背景度量和地標錨定。
歐式認知地圖基于環境中心的參考框架(allocentric reference frame), 不受導航者具身經驗的影響(圖3a, b), 類似于制圖學意義上的地圖, 保留了物理空間中已知位置間包括距離、角度在內的所有幾何特征和信息(圖3c), 便于新異路徑和捷徑的選擇, 從而讓導航過程變得高度靈活(Gallistel, 1990), 而不必拘泥于實際經歷過的有限幾條路徑。Widdowson和Wang (2022)發現無論空間曲率如何(歐式空間、雙曲空間或球面空間), 被試進行指向任務時總會與歐式方向匹配, 據此表明路徑整合和空間更新系統是基于歐式幾何學運作的。
不過以往也有研究發現人類的空間任務表現并不符合歐式幾何的基本假定(Beals et al., 1968; Tversky, 1981), 這對歐式認知地圖假設提出了挑戰。歐式度量空間的必要條件是該空間內的距離度量要滿足如下假定:(1)非負性:點A與其自身的距離為0, 兩個不同的點A、B之間的距離大于0; (2)對稱性:AB = BA; ③可加性:若C為線段AB上的點, 則AB = AC + BC; (3)三角不等性:對于任意三點A、B、C, AC + BC ≥ AC (圖3d)。此外, 若A、B、C三點構成直角三角形, 還應滿足勾股定理。實驗證據顯示, 方向和位置節點的數量都會影響顯式的(explicit)距離估計, 判斷一個地標與非地標之間的距離要比相反的情況更小(Burroughs & Sadalla, 1979; Moar & Carleton, 1982), 因此不符合對稱性; 而路徑上位置點數量越多, 距離估計會更大(Thorndyke, 1981), 因此不符合可加性; 此外距離估計也并不符合三角不等性(Byrne, 1979)。甚至在虛擬“蟲洞”中導航卻沒有意識到空間扭曲和幾何不一致性的存在, 表現出被試對于歐式結構的不敏感性(Warren et al., 2017)。
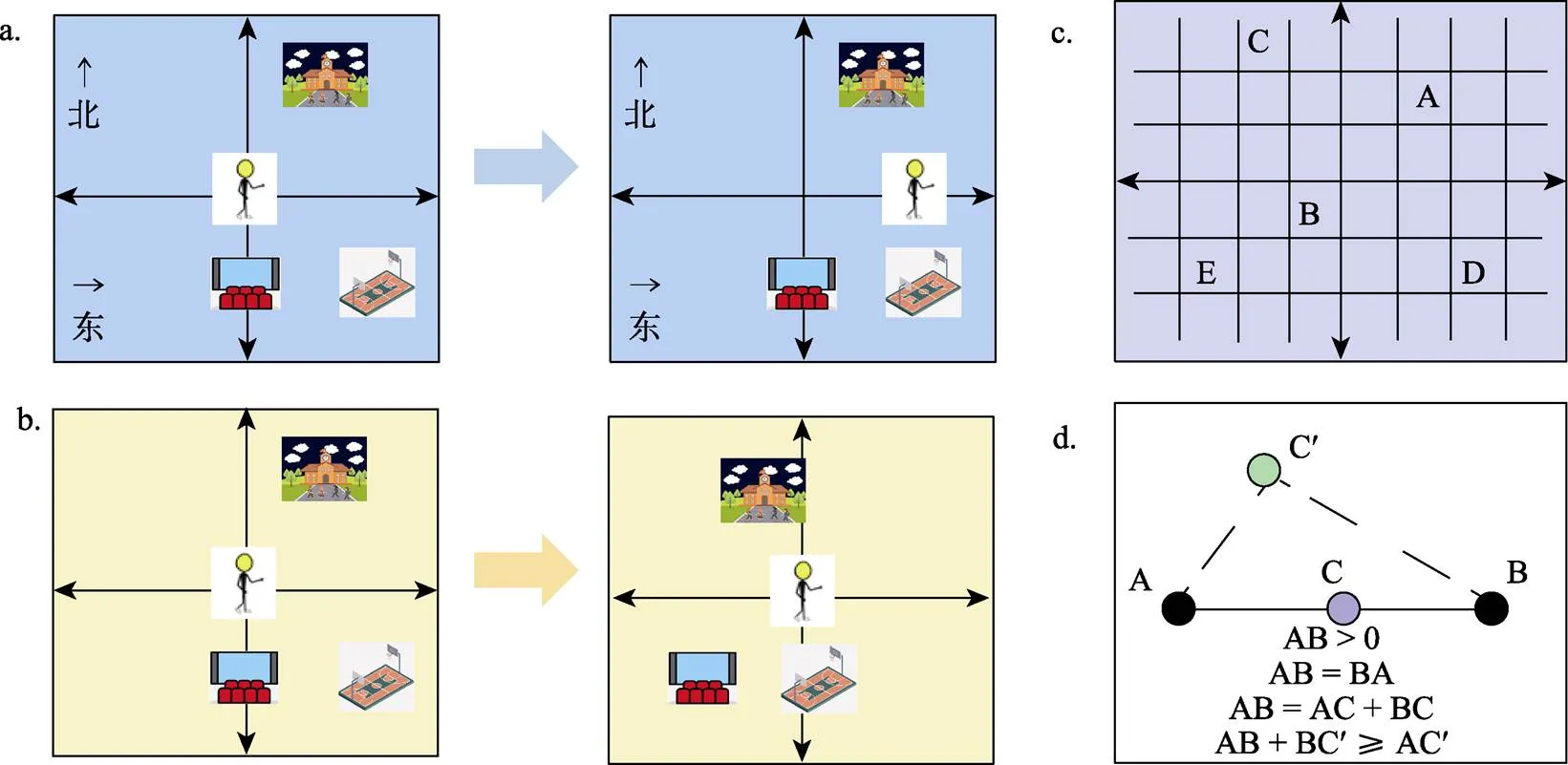
圖3 a. 環境中心的參考框架(allocentric reference frame), 導航者對環境的表征不受導航者自身移動的影響; b. 自我中心的參考框架(egocentric reference frame), 導航者始終以自身為參考中心, 對環境的表征會隨著自身移動而變化; c. 歐式認知地圖的示意圖, 包含具體的位置坐標、方向角度等全面的度量信息; d. 歐式幾何的基本假定。
雖然被試在空間實驗任務中往往會表現出距離估計上的較大誤差, 但是這不足以否定歐式地圖假設, 認知地圖仍然可能是存在一定噪音的歐式地圖。不過這就進一步導致歐式地圖假設可能無法證偽(Warren, 2019), 因為既然度量性地圖無法做到完全精確, 適度的誤差是合理的, 那么就需要考慮究竟多大范圍內的誤差是歐式地圖假設所能容忍的, 而這是一個難以嚴格定義的問題。
4.2 拓撲圖結構假設
對歐式地圖假設的早期反對來自于Eichenbaum (1997), 他認為海馬位置細胞并不足以作為歐式地圖的支持證據, 因為海馬并非專一于空間記憶或空間知識表征, 人類和動物海馬損傷會造成廣泛的記憶缺陷, 所以海馬細胞參與的應該是廣泛記憶空間的表征。Eichenbaum等人(1999)進一步提出記憶空間理論, 該理論認為認知地圖本質上是在記憶空間中對既往經歷的記錄, 首先由不同事件或位置節點的序列構成情節記憶, 其后不同情節記憶經由重復或公共元素(也稱節點)的關聯共同建立起記憶空間(圖4a), 對物理空間的表征僅僅是記憶空間表征的一個特例(Eichenbaum et al., 1999)。
在記憶空間理論的啟發下, 后續研究者提出了拓撲圖假設作為備選的認知地圖存在形式。不同于歐式地圖的精確度量, 拓撲圖假設認為認知地圖僅僅編碼粗糙的拓撲圖結構。環境的拓撲圖結構是由節點和節點間的連接邊共同構成的網絡, 一般而言節點表示位置, 而邊代表位置間的路徑關系(圖4b)。這種“位置圖(place graph)”捕獲了位置間的連接性, 但不需要將位置嵌入到全局一致的坐標系統中(coordinate free)。除了位置外, 節點也可以表示特定視角(view)或局部區域(vista); 相應地, 邊則是在不同視角或區域間進行轉換所需要的動作(Jacobs & Schenk, 2003; Mallot & Basten, 2009)。
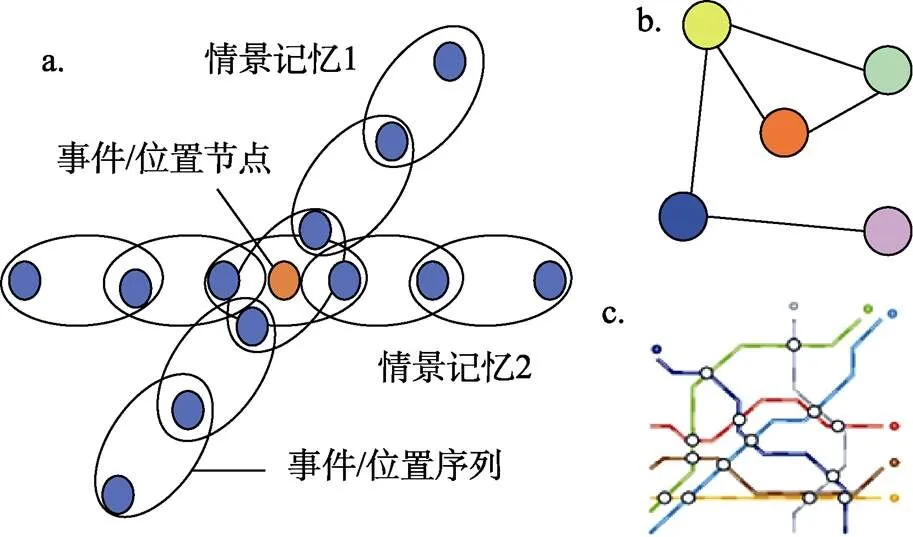
圖4 a. 記憶空間理論示意圖, 每個有色圓點代表特定的事件或位置, 連續發生的事件或連續經歷的位置序列構成一條情景記憶, 不同情景記憶之間可能存在公共元素(紅色圓點), 借此建立記憶空間(圖改編自Eichenbaum et al., 1999)。 b. 拓撲圖的示意圖, 節點代表位置, 節點間的連接邊代表位置間的轉移關系。 c. 地鐵線路圖——拓撲圖在現實生活中的案例, 不同顏色線條代表不同線路, 線路上空心圓代表站點。彩圖見電子版。
拓撲圖的信息含量介于歐式地圖和路線序列記憶之間。一方面, 圖結構知識比路線知識更為豐富。路線是串聯位置點和動作的簡單鏈條, 可能只是路徑網絡的一部分, 只支持沿著熟悉路徑的導航, 缺乏靈活性。而拓撲圖則能包含兩個位置之間的多條路徑, 可以通過路徑片段的重新組合實現新異路徑或繞路。另一方面, 相比歐式地圖, 拓撲圖對記憶資源的占用更少, 節點和邊以壓縮的結構存在, 不包含任何距離度量和角度信息, 不需要對位置坐標進行詳細計算操作, 但與此同時也就無法解釋人類在一定程度上能夠選擇捷徑的行為表現。所以純粹的拓撲圖表征對人的空間導航而言是不足夠的。
拓撲圖表征在人們的社會生活中比較常見, 譬如地鐵線路圖(圖4c), 同一條線路上不同站臺對應的位置名被等間距放置, 只突出強調站臺之間的連通性, 而并沒有關注或展示站臺間的距離, 并且相比于城市外緣區域, 市中心的面積會被放大, 站臺之間距離被高估(Longo, 2021), 側面反映出市中心區域客流量更大, 對站點信息詳細度的需求更高。動物的路線網絡也有類圖屬性, 如卷尾猴在樹枝交叉點通常會選擇不同的樹枝以便覓食, 表明猴子能夠分割路徑片段并組合成有實用價值的新路徑(Presotto et al., 2018)。神經層面上也發現部分海馬位置細胞的位置域會受到環境拓撲結構變化的影響(Dabaghian et al., 2014; Widloski & Foster, 2022)。
4.3 折中或融合假設
如果以度量信息的精確程度作為特征軸來定位有關認知地圖存在形式的不同理論, 那么歐式地圖假設和純粹的拓撲圖假設應該分別位于精確和粗略兩個極端(圖5a)。而另一方面, 空間表征既對環境的拓撲結構敏感, 同時也編碼部分的歐式信息, 盡管可能存在一定程度的誤差。因此, 有研究者試圖調和兩種假設形成折中方案, 即將拓撲圖結構和歐式信息適度融合, 進而形成混合型的認知地圖(hybrid cognitive map)。
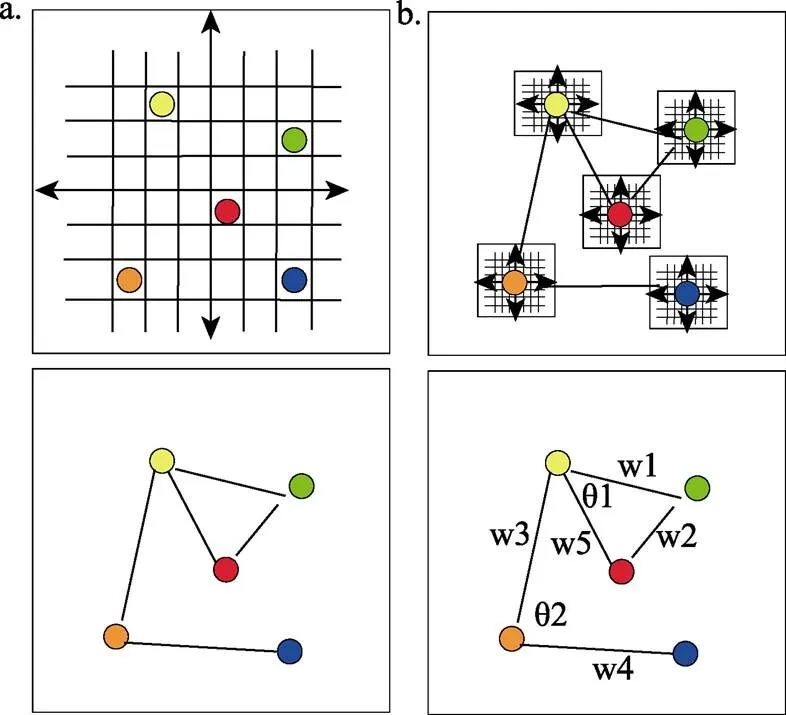
圖5 a. 歐式地圖(上)和拓撲圖(下)的示意圖; b. 參考框架網絡理論(上)在每個局部區域都存在精確的歐式參考框架, 有標簽圖假設(下)在節點間輔以夾角信息和連接邊權重。
Meilinger (2008)最早提出參考框架網絡理論(network of reference frames), 在這個模型中, 每個局部區域都有一個小范圍的度量性參考框架, 這些局部的參考框架作為圖結構中的節點由邊連接起來, 這些連接邊代表在不同參考框架間轉移時的視角轉換(圖5b上)。不過值得注意的是, 所有的局部參考框架無需被整合到統一的全局參考框架中, 新異路徑和捷徑的選擇需要從當前位置逐步轉換視角和參考框架直至到達目標位置。因此這個理論認為, 認知地圖在小尺度的局部環境下是精確的歐式地圖, 而在大尺度的全局環境下則是粗略的拓撲圖。
參考框架網絡理論放松了歐式地圖的全局一致性這一嚴格約束, 因此在減輕記憶負載的同時也容許空間表征存在一定的誤差和扭曲, 從而更加符合以往實際導航任務中人們的行為表現。但是在每一個局部區域都要精確地在度量性參考框架下表征信息仍舊很繁瑣, 并且局部區域間的路徑關系(如長度、方位夾角)似乎也十分不明確。Warren (2019)提出的有標簽圖假設在上述理論基礎上進行了適度的簡化和補充。
有標簽圖假設(labeled graph)沒有過多關注局部區域內的精確表征, 而是在不同的局部區域節點之間輔以度量性信息, 以邊的權重表示路徑的大致長度, 以節點處的標簽指示交叉點處毗鄰路徑間的大致夾角(圖5b下)。與參考框架網絡理論的共同點在于, 有標簽圖假設也無需在一個全局統一的參考框架內表征所有空間信息。
歐式地圖和拓撲圖對于空間表征都是必要的, 但具體建立和使用會依賴于環境特征和任務需求(Peer et al., 2021)。通過將度量信息限制在有限的局部, 一方面避免認知超載, 另一方面也能較好地為空間表征的不精確性或誤差提供解釋。這些融合假設對歐式地圖和拓撲圖二者的理論爭議進行了折中, 確實能夠更完善地解釋已有的實驗證據, 但是本質上并沒有創新。
4.4 統一或整合假設
物理空間、抽象關系空間內的很多環境其潛在結構本質上是一樣的, 只是具體的感覺刺激不同而已, 例如不同物理環境都符合歐式規則但環境內部的具體位置組織卻不同、不同家庭的家譜都有相同的分支結構但具體的家庭成員卻不同。如果能在內在結構相似但具體刺激不同的環境間泛化, 可以提高新環境下空間表征的效率。基于這個思路, Whittington等人(2020)提出TEM (Tolman- Eichenbaum Machine)模型作為統一框架來整合歐式地圖和拓撲圖這兩種形式, 該模型的命名正是為了紀念分別與這兩種表征形式有淵源的學者Tolman和Eichenbaum。
TEM的基本假設是環境或任務內在的抽象結構泛化能為空間表征和推理提供極大便利, 海馬結構的空間記憶和關系記憶都遵循結構泛化這個共同原則, 具體包括結構抽象和特定的感覺信息綁定兩個過程。結構泛化的關鍵在于因子化和重新組合, 即把不同方面的知識(如潛在結構和特定感覺刺激)分開表征形成獨立因子, 以便后續面對新情境可以靈活重組。結構抽象的過程發生在內側內嗅皮層, 抽象出的結構可以在不同的感覺刺激間泛化, 從而實現重映射(remapping); 特定的感覺刺激綁定則發生在海馬。
經過基于上述原則的學習和訓練后, TEM模擬的內嗅皮層確實表現出多種細胞的特性, 如網格細胞、邊界細胞、物體向量細胞等; 而TEM模擬的海馬細胞也表現出位置細胞的性質。這些TEM模擬細胞的活動模式能夠很好地吻合物理空間和抽象空間的實驗結果, 表明結構泛化的基本原則是有效的, 有可能是歐式地圖和拓撲圖共同的底層機制, 由此表明二者在心理表征上可能是一體兩面的。
此外, 后繼表征(successor representation, SR)理論也認為歐式地圖和拓撲圖表征只是相同潛在神經編碼的不同表現。SR指出海馬可能并不只是編碼當前的位置和狀態, 而是側重編碼對未來導航更有價值的后繼狀態(Stachenfeld et al., 2017; de Cothi et al., 2022), 通過編碼后續可能達到的位置及似然度分布, 構建起對所處環境的認知地圖。在拓撲圖結構中, 每個狀態為一個節點, 狀態間的轉移概率為節點間的聯結權重; 在歐式地圖中, 每個狀態為一個關鍵的歐式位置, 狀態間轉移則對應于不同位置間的路徑。
4.5 層級性理論
在上述歐式地圖和拓撲圖理論中, 認知地圖所包含的不同位置都處于無差別的表征水平, 并沒有高低隸屬關系的區分, 然而正如上文“2.3 層級性”這部分提到的, 環境空間往往會存在嵌套結構, 因此在認知地圖上不同區域的表征間可能具有層級性結構(hierarchical organization), 即環境的不同尺度范圍或不同地理單元的信息是按層級加以表征的(趙民濤, 2006)。
具體而言, 早期研究者提出的層級理論(hierarchical theory)認為, 一個環境的不同區域存儲在樹狀結構的不同分支, 即空間知識并不是在同一水平上進行表征的, 而是越細節化的空間知識會被存儲或表征在越低的等級, 反之越抽象概括的空間知識則對應到更高的等級。不同區域間的分界線可能是客觀存在的, 也可能只是主觀存在的(Hirtle & Jonides, 1985), 相比于區域內的空間表征, 跨越區域分界線的物體空間關系或距離表征精確度更低(Wang & Brockmole, 2003)。
層級理論適用的環境一般具有相對較大的空間尺度或相對復雜的內部結構, 環境結構的復雜度可能由物理因素(如墻壁阻隔)或人文原因(如行政區劃)導致, 而簡單的小尺度空間一般并不具備明顯的多區域分支結構(如單視點空間), 因此在這些空間類型中表征一般都并沒有突出的層級性。根據空間關系在記憶中編碼的類型可以將層級理論分成強層級理論和部分層級理論這兩個子理論(McNamara, 1986)。
強層級理論(strongly hierarchical theory)認為處于同一層級不同分支上的位置之間不存在空間關系編碼, 它們的空間關系必須從更高級的空間知識中推導得到(圖6)。比如同一個省內的兩個城市間的空間關系并不會被直接編碼, 而是通過比較二者在該省內的各自位置來間接得到。強層級理論在空間知識的存儲上更節約記憶資源, 但是很明顯喪失了靈活性。
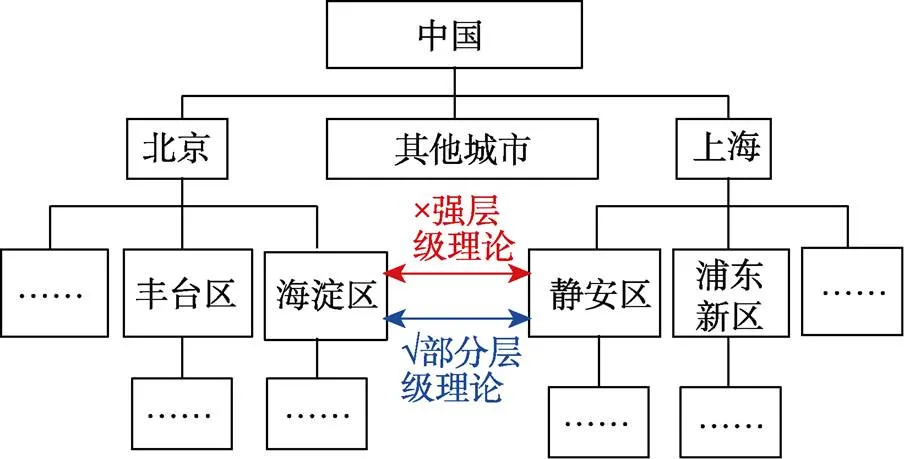
圖6 空間表征層級理論的示意圖。強層級理論認為同一層級不同分支區域內的位置之間不存在空間關系編碼, 而部分層級理論則相反。
不同于強層級理論, 部分層級理論(partially hierarchical theory)認為一個環境中不同區域內的位置間是可以編碼空間關系的(Stevens & Coupe, 1978) (圖6), 在空間知識的存儲上相對更麻煩(要編碼的空間關系更多), 但是這種冗余性存在的優勢是進行空間判斷的速度和準確性都更高, 畢竟直接使用要比推導后才能使用更高效。McNamara (1986)發現空間任務表現都會受到不同位置所在區域的影響, 支持部分層級理論。但是近年來, 層級理論受到的關注較少, 歐式地圖和拓撲圖相關研究也沒有將認知地圖的層級性同時納入考量。
5 研究展望
5.1 認知地圖的層級性變化
認知地圖的層級性會在空間環境具有嵌套結構時表現出來, 而歐式地圖和拓撲圖這兩種形式都沒有考慮層級性的問題。本研究推測, 層級性與空間尺度密切相關, 認知地圖的層級性會在表征大尺度空間時出現, 而歐式表征和拓撲表征則存在于其中尺度相對較小的某一層級內, 但是由于認知地圖的構建是動態的過程, 大尺度空間并非是一成不變的, 隨著認知地圖的逐步擴張, 最初的大尺度空間下不同區域的邊界可能會逐漸重疊, 因此空間表征也相應地發生融合, 從而最初的大尺度空間表征所具有的層級性可能會在這個過程逐步減小甚至最終消失, 形成全局的均一認知地圖, 同時包含歐式表征和拓撲表征兩類信息, 并且建立起新的路徑關系以連接原來隸屬于不同上位等級的子區域(如圖7)。
不同尺度環境下的層級表征是沿著由海馬到前額葉的從后往前順序進行組織的, 前額葉細胞有更大的預測視野, 而海馬細胞的預測視野則相對較小(Brunec & Momennejad, 2022), 因此前額葉可能處于層級表征的上位, 編碼高水平的規劃, 近期的一項地鐵線路圖導航研究也證實了這一點(Liang et al., 2022)。場景選擇區域對場景(如局部位置點)敏感, 將局部區域“編織”形成全局認知地圖, 可能填補了在表征內容上前額葉與海馬?內嗅系統之間的空白。
不過認知地圖的擴張潛力是有限的, 因為人的空間認知資源是受限的, 所能構建的認知地圖必然會存在尺度范圍的上限, 不同個體的上限可能會存在差異。未來研究應當關注認知地圖構建的動態過程中是否存在層級性下降的現象, 并且進一步探究人類認知地圖的尺度限制, 此外歐式地圖和拓撲圖在認知地圖發生的過程中如何實現整合、兩種表征的能力是否存在關聯也是值得研究的問題。
5.2 空間維度和范圍的擴展
傳統的空間表征研究所關注的主要是常規的二維物理空間, 由此導致多數研究者對空間表征的認識也是扁平化的(例如將二維地圖與之類比)。隨著導航認知領域的不斷發展, 更多的創新范式涌現, 實驗中的可導航空間也包含了更多新穎的形式。
一方面, 物理空間出現維度的拓展、曲率的操縱以及扭曲規則的設計。歐式空間的維度上, 從二維拓展到三維, 例如Kim和Doeller (2022)讓被試采取緊貼地面移動或飛行等不同方式在由平面和弧面構成的三維空間中探索學習, 隨后進行距離估計測試, 發現在能鳥瞰全局時(即直接暴露于環境的三維結構中)歐氏距離估計表現更好, 表明空間表征并非局限于二維平面, 而是會適應于行為經驗和任務需求。此外, 在非歐空間如雙曲空間或球面空間內, 被試完成指向任務時仍與歐式規則相匹配, 表明空間表征可能會將日常生活經驗沿用到不同曲率的虛擬空間中(Widdowson & Wang, 2022)。然而, 當歐式空間發生扭曲(如存在“蟲洞”, 可以在兩個位置點間瞬時穿越)時, 被試卻可能在沒有察覺到這種扭曲的情況下產生行為偏差(Warren et al., 2017)。這些研究主要關注行為表現, 沒有考察海馬、內嗅等相關腦區在表征這些不常見的物理空間時發生了何種變化、與常規空間表征是否存在共同機制等, 未來有必要將這些創新的空間范式與神經測量手段結合起來。

圖7 認知地圖的層級性變化假設。圖中A, B和C代表位于上位等級的區域, 而各自之下樹狀分支的三個位置是它們的子區域, 同一上位等級區域內的不同子區域間(圖中不同顏色的1, 2, 3)存在拓撲關系, 并且歐式位置信息也會被同時表征。隨著認知地圖的形成, 空間表征逐漸從局部擴展到全局, 相應負責的腦區也會變化(左側藍色虛線代表認知地圖的形成, 紅色虛線則表示調用認知地圖進行導航規劃的過程)。空間表征的層級性并非一成不變的, 可能會隨著認知地圖的逐步擴大, 而將不同區域的邊界進行融合, 最終有可能所有位置表征之間的層級性會逐漸消失, 即形成圖中A, B和C三個區域內9個位置的歐式表征和拓撲表征整合后的認知地圖。彩圖見電子版。
另一方面, 空間知識開始在廣義上指向抽象的認知空間, 包括社會關系空間、概念空間、價值空間和語義空間等, 而不再僅局限于物理空間(Schafer & Schiller, 2018)。空間隱喻在日常生活中十分常見, 比如社會等級和人際關系的很多描述其實與空間表達密切相關, 如“高”攀、“上”坐、疏“遠”、親“近”等。社會關系的感知反過來也會影響物理空間知識的表征, 更密切的關系會縮短主觀表征的距離(Kerkman et al., 2004)。社會信息的編碼類似于空間信息, 將每個社會個體表示為坐標, 根據特征或屬性確立社會空間的評估維度(如權力、受歡迎度) (Park et al., 2020), 進而可以進行社會推導和決策。人類可以在抽象的特征空間中編碼潛在的社會關系模式, 以靈活的方式構建社交網絡(Son et al., 2021)。包括海馬、楔前葉、背外側前額葉和腦島等多個腦區在內的腦網絡共同參與社會空間中的導航(Zhang et al., 2022)。不過社會空間中的很多元素并不能很清楚地與物理空間進行一一對應, 比如一個社交場合的邊界無法明確定義、人際關系空間中距離的度量維度沒有上限, 未來研究還需要進一步厘清這些模糊點。
5.3 認知地圖假說的局限性
認知地圖是對空間表征的直觀比喻, 具有很高的表面效度, 把復雜抽象的空間表征描述為簡單形象的地圖, 十分易于接受和理解, 但也可能存在 “表征論者謬誤” (representationalist fallacy), 即并不存在這樣一幅獨立于導航者本身而存在的地圖可供參考, 這種錯覺可能源于日常生活中實用地圖的普及和研究者的盲目類比(Warren, 2019)。空間導航相關的神經系統活動沒有必要拷貝物理環境, 而是要幫助人和其他動物能夠更好地保持自身與其所處環境之間的有效關聯, 從而更有利于生存(Sung et al., 2021)。
Tversky (2005)曾提出容忍系統誤差存在的認知拼貼畫(cognitive collages)和簡單環境下的空間心理模型(spatial mental models)來取代認知地圖的比喻。近期Farzanfar等人(2023)進一步提出了空間圖式(spatial schema)的概念, 他們假設有相似環境結構的導航經歷會形成空間圖式(如布局類似的城市)。具體而言, 環境空間信息的主旨和細節是分開表征的, 分別對應著空間概要和認知地圖, 其中空間概要(spatial gist)是指特定環境的核心特征, 表征腦區是海馬前部; 細節信息則在海馬后部表征, 二者都是針對特定環境而言的。而空間圖式則是更上位的空間表征, 從多個空間概要中提煉得到, 通過提取同類環境中可泛化的相似特征(如地理、視覺空間特征), 進而參與優化導航決策, 主要表征腦區是內側前額葉。
認知地圖的理論假設和早期證據都源于嚙齒類動物行為學和電生理的研究成果, 但是人腦表征信息的方式不同于嚙齒類, 這種差異貫穿了空間表征的各個方面, 包括對環境結構和子區域、路線、方向以及距離的表征(Zhao, 2018), 因此未來研究應當更多地關注人類自身的導航行為, 不宜直接進行跨物種的理論遷移。
此外認知地圖概念不清晰導致涵蓋的范圍也不明確, 甚至是過于寬泛。由于認知地圖是源于類比的概念, 有很多模糊的地方, 如地圖的尺度、細節和精確程度以及表征形式等, 所以可能出現兩項研究的內容存在很大差異卻都以認知地圖來冠名的現象, 從簡單的條件化任務(Costa et al., 2023)到復雜的空間導航(Epstein et al., 2017), 不同任務中所涉及的變量和要求可能存在很大區別, 卻都無差別地歸入認知地圖的范疇。未來研究需要審視認知地圖這個概念涵蓋的范圍, 避免擴大化; 此外應將關注點放在空間表征的本質、特征和發生過程的神經動態機制等方面。
5.4 結語
認知地圖思想的哲學起源是康德關于空間表征本性的觀點, 康德(1781/2004)在《純粹理性批判》中提出空間表征是先于經驗而存在的, 因此具有先天性。在康德的空間表征先天性理論和Tolman (1948)認知地圖思想基礎上, O’ Keefe和Nadel (1978)明確指出空間本性是絕對的, 因此認知地圖是歐式地圖, 其生理基礎是海馬中表征絕對位置的細胞(以及后續在內嗅皮層中發現的、為位置表征提供標度的網格細胞), 同時反對相對(或關系)空間理論。然而, 海馬的功能并不局限在物理空間內, 而是參與廣泛的記憶空間表征, 因此后續Eichenbaum等人(1999)提出記憶空間理論, 認知地圖表征相對連接關系的拓撲圖理論隨后出現。
歐式地圖和拓撲圖這兩種認知地圖的可能存在形式都有其合理性和局限之處, 神經機制層面二者都在一定程度上得到了驗證, 并且分別對認知地圖的某些基本屬性十分必要, 因此近期研究者們試圖將二者組合或統一起來, 以更完整地描述認知地圖。這些理論雖然兼顧歐式地圖和拓撲圖的特征, 但是卻忽視了空間層級性在認知地圖構建中的角色和影響。
嵌套空間表征的層級性在行為現象上多有發現, 并且早期也有研究者提出層級性理論, 但對于這類空間如何在大腦中表征的研究卻十分稀少(Kim & Maguire, 2018), 在理論和實證研究上歐式地圖和拓撲圖都未與認知地圖的層級性相聯系。二者都有可能在嵌套的空間中被構建, 因此這可能并不是因為歐式地圖和拓撲圖都不具備層級性表征, 而是因為以往研究者所采用的研究范式更多地關注認知地圖構建完畢后的特征, 而沒有考察構建的動態過程。在通過學習構建認知地圖的過程中, 層級性可能會逐步下降, 未來研究需要進一步探索和檢驗這個推測。
此外, 認知地圖的研究動態也提示, 未來還需要關注空間維度和范圍的擴展, 比較社會空間、語義空間與傳統研究的物理空間中認知地圖的差別和共性。多種行為、建模和神經現象揭示認知地圖具有選擇性、扭曲性和靈活性等基本屬性, 未來需要在這些屬性的基礎上進一步完善理論建構, 反思認知地圖假說可能存在的局限性。
何詩, 陰劼. (2022). 認知地圖的地理學研究進展與展望.(1), 73?85.
康德. (2004).(鄧曉芒譯). 北京:人民出版社. (原著1781)
梁群君, 鄭森寧, 李金惠, 黃瑞旺. (2022). 壓后皮質在空間導航的作用: 認知地圖與環境的映射.(1), 2?8.
阮曉鋼, 柴潔, 武悅, 張曉平, 黃靜. (2021). 基于海馬體位置細胞的認知地圖構建與導航.(3), 666?677.
王琳, 王亮. (2017). 認知地圖的神經環路基礎.(3), 187?197.
趙民濤. (2006). 物體位置與空間關系的心理表征.(3), 321?327.
Alme, C. B., Miao, C., Jezek, K., Treves, A., Moser, E. I., & Moser, M.-B. (2014). Place cells in the hippocampus: Eleven maps for eleven rooms.(52), 18428?18435.
Anggraini, D., Glasauer, S., & Wunderlich, K. (2018). Neural signatures of reinforcement learning correlate with strategy adoption during spatial navigation.(1), 10110.
Appleyard, D. (1970). Styles and methods of structuring a city.(1), 100?117.
Baldassano, C., Chen, J., Zadbood, A., Pillow, J. W., Hasson, U., & Norman, K. A. (2017). Discovering event structure in continuous narrative perception and memory.(3), 709?721.
Beals, R., Krantz, D. H., & Tversky, A. (1968). Foundations of multidimensional scaling.(2), 127-142.
Bellmund, J. L., de Cothi, W., Ruiter, T. A., Nau, M., Barry, C., & Doeller, C. F. (2020). Deforming the metric of cognitive maps distorts memory.(2), 177?188.
Bellmund, J. L., Deuker, L., Navarro Schr?der, T., & Doeller, C. F. (2016). Grid-cell representations in mental simulation., e17089. https://doi.org/10.7554/eLife.17089.001
Borges, J. L., & Casares A. B. (Eds). (1971).. New York: Herder and Herder.
Brown, T. I., Gagnon, S. A., & Wagner, A. D. (2020). Stress disrupts human hippocampal-prefrontal function during prospective spatial navigation and hinders flexible behavior.(10), 1821?1833.
Brown, T. I., Ross, R. S., Tobyne, S. M., & Stern, C. E. (2012). Cooperative interactions between hippocampal and striatal systems support flexible navigation.(2), 1316?1330.
Brunec, I. K., Bellana, B., Ozubko, J. D., Man, V., Robin, J., Liu, Z. X., ... Moscovitch, M. (2018). Multiple scales of representation along the hippocampal anteroposterior axis in humans.(13), 2129?2135.
Brunec, I. K., & Momennejad, I. (2022). Predictive representations in hippocampal and prefrontal hierarchies.(2), 299?312.
Burroughs, W. J., & Sadalla, E. K. (1979). Asymmetries in distance cognition.(4), 414?421.
Byrne, R. W. (1979). Memory for urban geography.(1), 147?154.
Carpenter, F., Manson, D., Jeffery, K., Burgess, N., & Barry, C. (2015). Grid cells form a global representation of connected environments.(9), 1176? 1182.
Chen, X., He, Q., Kelly, J. W., Fiete, I. R., & McNamara, T. P. (2015). Bias in human path integration is predicted by properties of grid cells.(13), 1771? 1776.
Costa, K. M., Scholz, R., Lloyd, K., Moreno-Castilla, P., Gardner, M. P., Dayan, P., & Schoenbaum, G. (2023). The role of the lateral orbitofrontal cortex in creating cognitive maps.,(1), 107?115.
Coutrot, A., Manley, E., Goodroe, S., Gahnstrom, C., Filomena, G., Yesiltepe, D., ... Spiers, H. J. (2022). Entropy of city street networks linked to future spatial navigation ability.(7904), 104?110.
Dabaghian, Y., Brandt, V. L., & Frank, L. M. (2014). Reconceiving the hippocampal map as a topological template.e03476. https://doi.org/10.7554/eLife. 03476.001
Dahmani, L., & Bohbot, V. D. (2020). Habitual use of GPS negatively impacts spatial memory during self-guided navigation.(1), 6310.
de Cothi, W., Nyberg, N., Griesbauer, E.-M., Ghanamé, C., Zisch, F., Lefort, J. M., ... Spiers, H. J. (2022). Predictive maps in rats and humans for spatial navigation.(17), 3676?3689.
Dilks, D. D., Kamps, F. S., & Persichetti, A. S. (2022). Three cortical scene systems and their development.(2), 117?127.
Doeller, C. F., Barry, C., & Burgess, N. (2010). Evidence for grid cells in a human memory network.(7281), 657?661.
Eichenbaum, H. (1997). Declarative memory: Insights from cognitive neurobiology.(1), 547?572.
Eichenbaum, H., Dudchenko, P., Wood, E., Shapiro, M., & Tanila, H. (1999). The hippocampus, memory, and place cells: Is it spatial memory or a memory space?(2), 209?226.
Epstein, R. A., Parker, W. E., & Feiler, A. M. (2007). Where am I now? Distinct roles for parahippocampal and retrosplenial cortices in place recognition.(23), 6141?6149.
Epstein, R. A., Patai, E. Z., Julian, J. B., & Spiers, H. J. (2017). The cognitive map in humans: Spatial navigation and beyond.(11), 1504?1513.
Epsztein, J. (2022). Mental replays enable flexible navigation.(7908), 35?36.
Evensmoen, H. R., Lehn, H., Xu, J., Witter, M. P., Nadel, L., & H?berg, A. K. (2013). The anterior hippocampus supports a coarse, global environmental representation and the posterior hippocampus supports fine-grained, local environmental representations.(11), 1908?1925.
Farzanfar, D., Spiers, H. J., Moscovitch, M., & Rosenbaum, R. S. (2023). From cognitive maps to spatial schemas.(2), 63?79.
Gahnstrom, C. J., & Spiers, H. J. (2020). Striatal and hippocampal contributions to flexible navigation in rats and humans., 1?7.
Gallistel, C. R. (1990).. Cambridge: The MIT Press.
Garvert, M. M., Saanum, T., Schulz, E., Schuck, N. W., & Doeller, C. F. (2023). Hippocampal spatio-predictive cognitive maps adaptively guide reward generalization.e(4), 615?626.
Gatti, D., Marelli, M., Vecchi, T., & Rinaldi, L. (2022). Spatial representations without spatial computations.(11), 1947?1958.
Genzel, D., Yovel, Y., & Yartsev, M. M. (2018). Neuroethology of bat navigation.(17), R997?R1004.
Goldshtein, A., Harten, L., & Yovel, Y. (2022). Mother bats facilitate pup navigation learning.(2), 350?360.
Golledge, R. G., Ruggles, A. J., Pellegrino, J. W., & Gale, N. D. (1993). Integrating route knowledge in an unfamiliar neighborhood: Along and across route experiments.(4), 293?307.
Hafting, T., Fyhn, M., Molden, S., Moser, M.-B., & Moser, E. I. (2005). Microstructure of a spatial map in the entorhinal cortex.(7052), 801?806.
Harten, L., Katz, A., Goldshtein, A., Handel, M., & Yovel, Y. (2020). The ontogeny of a mammalian cognitive map in the real world.(6500), 194?197.
Hartley, S. (2017).New York: Simon and Schuster.
Henriksson, L., Mur, M., & Kriegeskorte, N. (2019). Rapid invariant encoding of scene layout in human OPA.(1), 161?171.
Hirtle, S. C., & Jonides, J. (1985). Evidence of hierarchies in cognitive maps.(3), 208?217.
Ho, M. K., Abel, D., Correa, C. G., Littman, M. L., Cohen, J. D., & Griffiths, T. L. (2022). People construct simplified mental representations to plan.(7912), 129? 136.
Howard, L. R., Javadi, A. H., Yu, Y., Mill, R. D., Morrison, L. C., Knight, R., ... Spiers, H. J. (2014). The hippocampus and entorhinal cortex encode the path and Euclidean distances to goals during navigation.(12), 1331?1340.
Jacobs, J., Weidemann, C. T., Miller, J. F., Solway, A., Burke, J. F., Wei, X.-X., ... Kahana, M. J. (2013). Direct recordings of grid-like neuronal activity in human spatial navigation.(9), 1188?1190.
Jacobs, L. F., & Schenk, F. (2003). Unpacking the cognitive map: The parallel map theory of hippocampal function.(2), 285-315.
Javadi, A.-H., Emo, B., Howard, L. R., Zisch, F. E., Yu, Y., Knight, R., Silva, J. P., & Spiers, H. J. (2017). Hippocampal and prefrontal processing of network topology to simulate the future.(1), 14652.
Javadi, A.-H., Patai, E. Z., Marin-Garcia, E., Margolis, A., Tan, H.-R. M., Kumaran, D., ... Spiers, H. J. (2019). Prefrontal dynamics associated with efficient detours and shortcuts: A combined functional magnetic resonance imaging and magnetoencenphalography study.(8), 1227?1247.
Julian, J. B., Ryan, J., Hamilton, R. H., & Epstein, R. A. (2016). The occipital place area is causally involved in representing environmental boundaries during navigation.(8), 1104?1109.
Kabadayi, C., Bobrowicz, K., & Osvath, M. (2018). The detour paradigm in animal cognition.(1), 21?35.
Kaefer, K., Nardin, M., Blahna, K., & Csicsvari, J. (2020). Replay of behavioral sequences in the medial prefrontal cortex during rule switching.(1), 154?165.
Kerkman, D. D., Stea, D., Norris, K., & Rice, J. L. (2004). Social attitudes predict biases in geographic knowledge.(2), 258?269.
Kim, M., & Doeller, C. F. (2022). Adaptive cognitive maps for curved surfaces in the 3D world., 105126. https://doi.org/10.1016/j.cognition.2022.105126
Kim, M., & Maguire, E. A. (2018). Hippocampus, retrosplenial and parahippocampal cortices encode multicompartment 3D space in a hierarchical manner.(5), 1898?1909.
Lacroix, L., White, I., & Feldon, J. (2002). Effect of excitotoxic lesions of rat medial prefrontal cortex on spatial memory.(1), 69? 81.
Lever, C., Burton, S., Jeewajee, A., O'Keefe, J., & Burgess, N. (2009). Boundary vector cells in the subiculum of the hippocampal formation.(31), 9771?9777.
Liang, Q., Li, J., Zheng, S., Liao, J., & Huang, R. (2022). Dynamic causal modelling of hierarchical planning.119384. https://doi.org/10.1016/j.neuroimage. 2022.119384
Longo, M. R. (2021). Distortion of mental body representations..(3), 241?254.
Mahmoodi, A., Nili, H., Harbison, C., Hamilton, S., Trudel, N., Bang, D., & Rushworth, M. F. (2023). Causal role of a neural system for separating and selecting multidimensional social cognitive information.(7), 1152?1164.
Mallot, H. A., & Basten, K. (2009). Embodied spatial cognition: Biological and artificial systems.(11), 1658?1670.
Marchette, S. A., Vass, L. K., Ryan, J., & Epstein, R. A. (2015). Outside looking in: landmark generalization in the human navigational system.(44), 14896?14908.
McNamara, T. P. (1986). Mental representations of spatial relations.(1), 87?121.
Meilinger, T. (2008). The network of reference frames theory: A synthesis of graphs and cognitive maps. In: Freksa, C., Newcombe, N. S., G?rdenfors, P., W?lfl, S. (Eds.),(Vol. 5248, pp. 344?360).Springer, Berlin, Heidelberg.
Meilinger, T., Strickrodt, M., & Bülthoff, H. H. (2016). Qualitative differences in memory for vista and environmental spaces are caused by opaque borders, not movement or successive presentation., 77?95.
Moar, I., & Carleton, L. R. (1982). Memory for routes.(3), 381?394.
Morgan, L. K., MacEvoy, S. P., Aguirre, G. K., & Epstein, R. A. (2011). Distances between real-world locations are represented in the human hippocampus.(4), 1238?1245.
Moser, E. I., Kropff, E., & Moser, M.-B. (2008). Place cells, grid cells, and the brain's spatial representation system., 69?89.
Muhle-Karbe, P. S., Sheahan, H., Pezzulo, G., Spiers, H. J., Chien, S., Schuck, N. W., & Summerfield, C. (2023). Goal-seeking compresses neural codes for space in the human hippocampus and orbitofrontal cortex., 2023-01. https://doi.org/10.1101/2023.01.12.523762
Nardin, M., Kaefer, K., & Csicsvari, J. (2021). The generalized spatial representation in the prefrontal cortex is inherited from the hippocampus., 2021-09. https://doi.org/10.1101/2021.09.30.462269
O’ Keefe, J., & Dostrovsky, J. (1971). The hippocampus as a spatial map: Preliminary evidence from unit activity in the freely-moving rat.(1), 171?175.
O’ Keefe, J., & Nadel, L. (1978).. Oxford: Oxford University Press.
Park, J., & Park, S. (2020). Coding of navigational distance and functional constraint of boundaries in the human scene-selective cortex.(18), 3621?3630.
Park, S. A., Miller, D. S., & Boorman, E. D. (2021). Inferences on a multidimensional social hierarchy use a grid-like code.(9), 1292?1301.
Park, S. A., Miller, D. S., Nili, H., Ranganath, C., & Boorman, E. D. (2020). Map making: Constructing, combining, and inferring on abstract cognitive maps.(6), 1226?1238.
Patai, E. Z., & Spiers, H. J. (2021). The versatile wayfinder: Prefrontal contributions to spatial navigation.(6), 520?533.
Peer, M., Brunec, I. K., Newcombe, N. S., & Epstein, R. A. (2021). Structuring knowledge with cognitive maps and cognitive graphs.(1), 37? 54.
Presotto, A., Verderane, M. P., Biondi, L., Mendon?a- Furtado, O. D., Spagnoletti, N., Madden, M., & Izar, P. (2018). Intersection as key locations for bearded capuchin monkeys (Sapajus libidinosus) traveling within a route network.(3), 393?405.
Robertson, C. E., Hermann, K. L., Mynick, A., Kravitz, D. J., & Kanwisher, N. (2016). Neural representations integrate the current field of view with the remembered 360 panorama in scene-selective cortex.(18), 2463?2468.
Sauer, J.-F., Folschweiller, S., & Bartos, M. (2022). Topographically organized representation of space and context in the medial prefrontal cortex.(6), e2117300119. https://doi.org/10.1073/pnas.2117300119
Schafer, M., & Schiller, D. (2018). Navigating social space.(2), 476?489.
Schinazi, V. R., & Epstein, R. A. (2010). Neural correlates of real-world route learning.(2), 725?735.
Schlesiger, M. I., Boublil, B. L., Hales, J. B., Leutgeb, J. K., & Leutgeb, S. (2018). Hippocampal global remapping can occur without input from the medial entorhinal cortex.(12), 3152?3159.
Schuck, N. W., Cai, M. B., Wilson, R. C., & Niv, Y. (2016). Human orbitofrontal cortex represents a cognitive map of state space.(6), 1402?1412.
Shallice, T., & Burgess, P. W. (1991). Deficits in strategy application following frontal lobe damage in man.(Pt 2), 727?741.
Sloan, H. L., Good, M., & Dunnett, S. B. (2006). Double dissociation between hippocampal and prefrontal lesions on an operant delayed matching task and a water maze reference memory task.(1), 116?126.
Son, J.-Y., Bhandari, A., & FeldmanHall, O. (2021). Cognitive maps of social features enable flexible inference in social networks.(39), e2021699118. https://doi.org/10. 1073/pnas.2021699118
Stachenfeld, K. L., Botvinick, M. M., & Gershman, S. J. (2017). The hippocampus as a predictive map.(11), 1643?1653.
Stensola, H., Stensola, T., Solstad, T., Fr?land, K., Moser, M.-B., & Moser, E. I. (2012). The entorhinal grid map is discretized.(7427), 72?78.
Stevens, A., & Coupe, P. (1978). Distortions in judged spatial relations.(4), 422?437.
Sung, J. Y., Harris, O. K., Hensley, N. M., Chemero, A. P., & Morehouse, N. I. (2021). Beyond cognitive templates: Re-examining template metaphors used for animal recognition and navigation.(3), 825?841.
Taube, J. S., Muller, R. U., & Ranck, J. B. (1990). Head- direction cells recorded from the postsubiculum in freely moving rats. I. Description and quantitative analysis.(2), 420?435.
Thorndyke, P. W. (1981). Distance estimation from cognitive maps.(4), 526?550.
Tversky, B. (1981). Distortions in memory for maps.(3), 407?433.
Tolman, E. C. (1948). Cognitive maps in rats and men.(4), 189–208. https://doi.org/10. 1037/h0061626
Tversky, B. (1993). Cognitive maps, cognitive collages, and spatial mental models. In: Frank, A. U., Campari, I. (Eds.),(Vol. 716, pp. 14?24). Springer, Berlin, Heidelberg.
Wang, R. F., & Brockmole, J. R. (2003). Human navigation in nested environments.(3), 398?404.
Warren, W. H. (2019). Non-euclidean navigation.(Suppl_1), jeb187971. https:// doi.org/10.1242/jeb.187971
Warren, W. H., Rothman, D. B., Schnapp, B. H., & Ericson, J. D. (2017). Wormholes in virtual space: From cognitive maps to cognitive graphs., 152?163.
Wernle, T., Waaga, T., M?rreaunet, M., Treves, A., Moser, M.-B., & Moser, E. I. (2018). Integration of grid maps in merged environments.(1), 92?101.
Whittington, J. C., Muller, T. H., Mark, S., Chen, G., Barry, C., Burgess, N., & Behrens, T. E. (2020). The Tolman - Eichenbaum machine: Unifying space and relational memory through generalization in the hippocampal formation.(5), 1249?1263.
Widdowson, C., & Wang, R. F. (2022). Human navigation in curved spaces., 104923. https://doi.org/ 10.1016/j.cognition.2021.104923
Widloski, J., & Foster, D. J. (2022). Flexible rerouting of hippocampal replay sequences around changing barriers in the absence of global place field remapping.(9), 1547?1558.
Wu, X., & Foster, D. J. (2014). Hippocampal replay captures the unique topological structure of a novel environment.(19), 6459?6469.
Yousif, S. R. (2022). Redundancy and reducibility in the formats of spatial representations.(6), 1778?1793.
Zhang, B., & Naya, Y. (2020). Medial prefrontal cortex represents the object-based cognitive map when remembering an egocentric target location.(10), 5356?5371.
Zhang, L., Chen, P., Schafer, M., Zheng, S., Chen, L., Wang, S., ... Huang, R. (2022). A specific brain network for a social map in the human brain.(1), 1773. https://doi.org/10.1038/s41598-022-05601-4
Zhao, M. (2018). Human spatial representation: What we cannot learn from the studies of rodent navigation.(5), 2453?2465.
Zong, W., Zhou, J., Gardner, M. P., Costa, K. M., Zhang, Z., & Schoenbaum, G. (2023). Schema cell formation in orbitofrontal cortex is suppressed by hippocampal output.2023-05. https://doi.org/10.1101/2023.05.03.539307
The cognitive map and its intrinsic mechanisms
WU Wenya, WANG Liang
(CAS Key Laboratory of Mental Health, Institute of Psychology, Chinese Academy of Sciences, Beijing 100101, China)(Department of Psychology, University of Chinese Academy of Sciences, Beijing 100049, China)
Spatial navigation is vital for the survival and reproduction of humans and other animals living in complex environments. Effective spatial representations, also known as cognitive maps, are the basis of efficient spatial navigation. Prototypical properties of cognitive maps include selectivity, flexibility, and hierarchy. Several brain regions such as the hippocampus, the scene-selective areas and the prefrontal cortex are involved in the construction of cognitive maps. There are two theoretical disputes in the representational formats of cognitive maps, namely Euclidean map and topological graph, neither of which can fully account for the navigational behavior. Therefore, several hybrid theories have been put forward in order to reconcile this controversy such as the labeled graph hypothesis, the reference frame network theory, etc. Future researchers are suggested to focus on dynamic changes of hierarchical organization during the process of constructing cognitive maps, expansion of spatial dimensions and categories, and limitations in the concept of cognitive maps.
cognitive map, Euclidean map, topological graph, hierarchy, spatial representation
2023-04-19
* 國家自然科學基金(32020103009), 科技創新2030 (2022ZD0205000), 中國科學院心理研究所自主部署項目(E2CX4215CX)
王亮, E-mail: lwang@psych.ac.cn
B842, B845
猜你喜歡
作文周刊·小學二年級版(2022年20期)2022-05-05 01:33:06
中老年保健(2021年12期)2021-08-24 03:30:40
中國傳媒大學學報(自然科學版)(2021年1期)2021-06-09 08:43:00
中國生殖健康(2020年6期)2020-02-01 06:28:50
創新作文(小學版)(2019年10期)2019-09-25 08:12:28
中國生殖健康(2019年11期)2019-01-07 01:28:02
小學生學習指導(低年級)(2017年5期)2017-05-04 04:14:38
中國科技博覽(2016年2期)2016-04-25 20:32:39
小學生導刊(2016年34期)2016-04-11 00:49:44
作文與考試·小學高年級版(2015年17期)2015-05-30 10:48:04
