基于mtDNA COX3 基因?qū)ξ鞑靥厣笈H后w遺傳結(jié)構(gòu)的分析
2023-09-25 03:43:00者玉琦武志娟王吉坤鐘金城柴志欣信金偉2
草業(yè)學(xué)報 2023年9期
關(guān)鍵詞:研究
者玉琦,武志娟,王吉坤,鐘金城,柴志欣,信金偉2
(1. 青藏高原動物遺傳資源保護與利用四川省重點實驗室,四川 成都 610225;2. 省部共建青稞和牦牛種質(zhì)資源與遺傳改良國家重點實驗室,西藏 拉薩 850000;3. 四川省青藏高原草食家畜工程技術(shù)中心,四川 成都 610225)
牦牛是青藏高原的特有畜種資源,在我國主要分布于青海、西藏、四川、甘肅、云南、新疆等海拔3500~5000 m的高寒草原草地。西藏自治區(qū)作為我國第二大牦牛養(yǎng)殖和主產(chǎn)區(qū),因其特殊的地理特征及生態(tài)環(huán)境形成了斯布牦牛、江達牦牛、嘉黎牦牛、類烏齊牦牛、桑日牦牛、工布江達牦牛等地方品種或群體,牦牛遺傳資源十分豐富。西藏自治區(qū)也成為目前我國地方牦牛品種及遺傳資源發(fā)掘潛力最大的區(qū)域,其中西藏高山牦牛、斯布牦牛、帕里牦牛、娘亞牦牛、類烏齊牦牛及查吾拉牦牛已列入國家畜禽品種遺傳資源名錄,江達牦牛也于2021 年提交申報國家優(yōu)良牦牛遺傳資源。牦牛是經(jīng)長期自然選擇和人工選擇形成的能適應(yīng)高寒、缺氧、強紫外線、牧草期短等極端環(huán)境條件的特色牛種,在遺傳上是一個極為寶貴的基因庫,因此應(yīng)該在正確分類以及認(rèn)識的基礎(chǔ)上對其合理利用,利用特色優(yōu)勢資源帶動畜產(chǎn)經(jīng)濟發(fā)展[1]。
線粒體DNA(mitochondrial DNA, mtDNA)作為高等動物唯一的核外遺傳物質(zhì),因其具有母系遺傳、多態(tài)性豐富、結(jié)構(gòu)簡單等特點被認(rèn)為是研究物種起源、演化和分類最好的分子遺傳標(biāo)記[2]。動物mtDNA 包含細(xì)胞色素C 氧化酶(cytochrome C oxidase,COX)3 個亞基基因(COX1,COX2,COX3)、Cytb、ATP6、ATP8等13 個蛋白基因,它們都是線粒體內(nèi)膜呼吸鏈的重要組成因子。細(xì)胞色素C 氧化酶是由細(xì)胞核基因組和線粒體基因組分別編碼的亞基共同組成的復(fù)雜復(fù)合物[3-4],COX3是線粒體呼吸鏈電子傳遞的終末復(fù)合物,屬亞鐵血紅素,銅氧化酶的超家族,是線粒體氧化能力的關(guān)鍵調(diào)節(jié)因子。已有研究表明,包括COX3在內(nèi)的細(xì)胞色素C 氧化酶相對保守,常用來研究物種分子系統(tǒng)演化及分類,被認(rèn)為是對屬內(nèi)不同種、種內(nèi)不同亞種或不同地理型之間物種鑒定較為有效的分子標(biāo)記[5-6]。目前,研究者基于線粒體COX3基因已對四川黑熊[7]、瓢雞[8]、長爪沙鼠[9]等物種開展了種系發(fā)育研究,獲得了比較有參考價值的分類結(jié)果,但利用COX3基因?qū)﹃笈_z傳多樣性及分子進化方面的研究還鮮有報道。本研究選取西藏7 個特色牦牛群體共140 頭個體,測定COX3基因蛋白質(zhì)編碼區(qū)(coding sequence,CDS)序列,分析其遺傳多樣性,探討不同群體間的系統(tǒng)進化關(guān)系,旨在為西藏牦牛遺傳資源保護及利用提供參考。
1 材料與方法
2022 年5 月樣品采集于西藏自治區(qū),相關(guān)試驗于2022 年7-9 月在西南民族大學(xué)青藏高原動物遺傳資源保護與利用四川省重點實驗室完成。
1.1 試驗材料
從西藏帕里牦牛、嘉黎牦牛、類烏齊牦牛、工布江達牦牛、斯布牦牛、桑日牦牛和江達牦牛中分別選取成年健康牦牛20 頭,共140 頭(表1),用耳樣采集器采集長寬各1 cm 耳組織,75%乙醇保存帶回實驗室,于-80 ℃保存?zhèn)溆谩?/p>

表1 樣本信息Table 1 Sample information
1.2 基因組DNA 提取及檢測
剪綠豆大小耳組織樣品去除毛發(fā)后,利用動物組織基因組DNA 提取試劑盒(TianGen 生物技術(shù)公司)提取其基因組DNA,用1%瓊脂糖凝膠電泳、超微量核酸蛋白測定儀(NanoDrop One,美國)分別檢測DNA 的純度和濃度,檢測符合要求后置于-20 ℃保存。
1.3 PCR 擴增及測序
根據(jù)GenBank 中公布的牦牛COX3基因CDS 核苷酸序列(NC_006380.3),利用Primer Premier 5.0 設(shè)計特異性引物,引物序列為:F:5′-TAATCGGAGGGGCTACACTT-3′,R:5′-TCGTATGGACTTGTCTTCTCA-3′。PCR 反應(yīng)體系為25 μL,分別加入上、下游引物各1 μL(10 pmol·L-1),DNA 模板1 μL,2×Long Taq DNA 預(yù)混酶12.5 μL,滅菌ddH2O 9.5 μL。PCR 擴增體系:94 ℃預(yù)變性4 min,94 ℃變性45 s,54 ℃退火35 s,72 ℃延伸1 min,共32 個循環(huán);72 ℃延伸10 min,4 ℃保存。將PCR 擴增產(chǎn)物經(jīng)1%瓊脂糖凝膠電泳分離,用膠回收試劑盒(OMEGA)回收純化,純化后的PCR 產(chǎn)物(每個體3 個重復(fù))送北京擎科生物科技有限公司成都分公司測序。
1.4 數(shù)據(jù)統(tǒng)計處理
利用DNAMAN 軟件對所測序列進行比對、校正獲得西藏牦牛mtDNACOX3基因CDS 序列;采用鼠小弟(https://cloud.keyandaydayup.com/)統(tǒng)計基因序列長度及堿基組成;利用DNASP 5.1 軟件進行多態(tài)位點、單倍型多樣性和核苷酸多樣性分析;根據(jù)西藏牦牛COX3基因CDS 單倍型序列的變異位點,利用MEGA 7.0 軟件計算群體間的Kimura 雙參數(shù)遺傳距離,構(gòu)建群體間聚類關(guān)系,并繪制單倍型網(wǎng)絡(luò)關(guān)系圖,采用鄰接法(neighbor joining,NJ)構(gòu)建牦牛系統(tǒng)發(fā)育樹。
2 結(jié)果與分析
2.1 西藏牦牛mtDNA COX3 基因的PCR 擴增
PCR 擴增產(chǎn)物經(jīng)凝膠成像系統(tǒng)檢測可見條帶單一,清晰明亮,大小為1200 bp 左右,且無雜帶及拖帶(圖1),符合后續(xù)試驗需要。
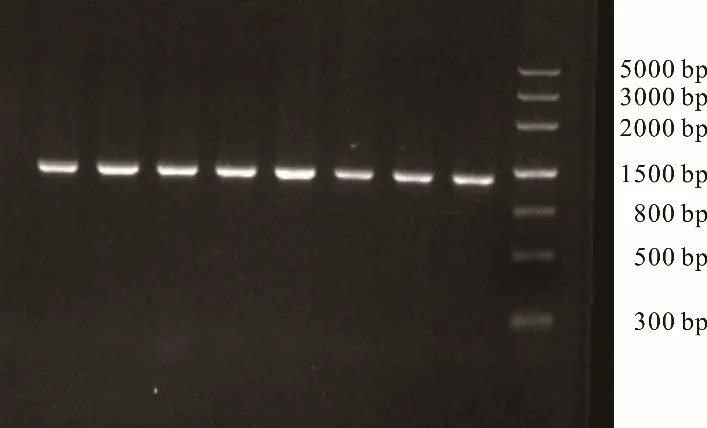
圖1 江達牦牛COX3 基因PCR 產(chǎn)物電泳Fig. 1 Electrophoresis of PCR product of COX3 gene of Jiangda yak
2.2 西藏牦牛mtDNA COX3 基因的遺傳多樣性
2.2.1 核苷酸及氨基酸組成 經(jīng)測序獲得7 個西藏牦牛群體共140 頭個體的COX3基因CDS 序列,其長度均為781 bp,T、C、A、G 4種核苷酸的平均比例分別為29.2%(28.9%~29.6%)、29.4%(28.9%~29.6%)、26.2%(26.0%~26.5%)、15.2%(15.0%~15.2%),其中A+T 含 量55.4%,G+C 含 量44.6%,A+T 堿 基含量占比較大。經(jīng)DNASP 5.1 軟件統(tǒng)計分析顯示,所有牦牛群體序列共有55 個多態(tài)位點,其中單一多態(tài)位點5個(位于144、217、757、765、769 位堿基處),約占多態(tài)位點總數(shù)的9.09%,簡約信息位點50 個(位于27、36、51、64、66、69、91、94、97、102、108、119、120、121、123、127、150、177、181、186、198、252、255、273、300、309、327、330、366、375、417、451、453、462、471、510、519、540、549、555、574、576、582、594、621、633、657、667、681、744 位堿基處),約占多態(tài)位點總數(shù)的90.91%。同義突變45 個,序列變異中包含堿基替換11 個,無堿基缺失、插入(表2)。
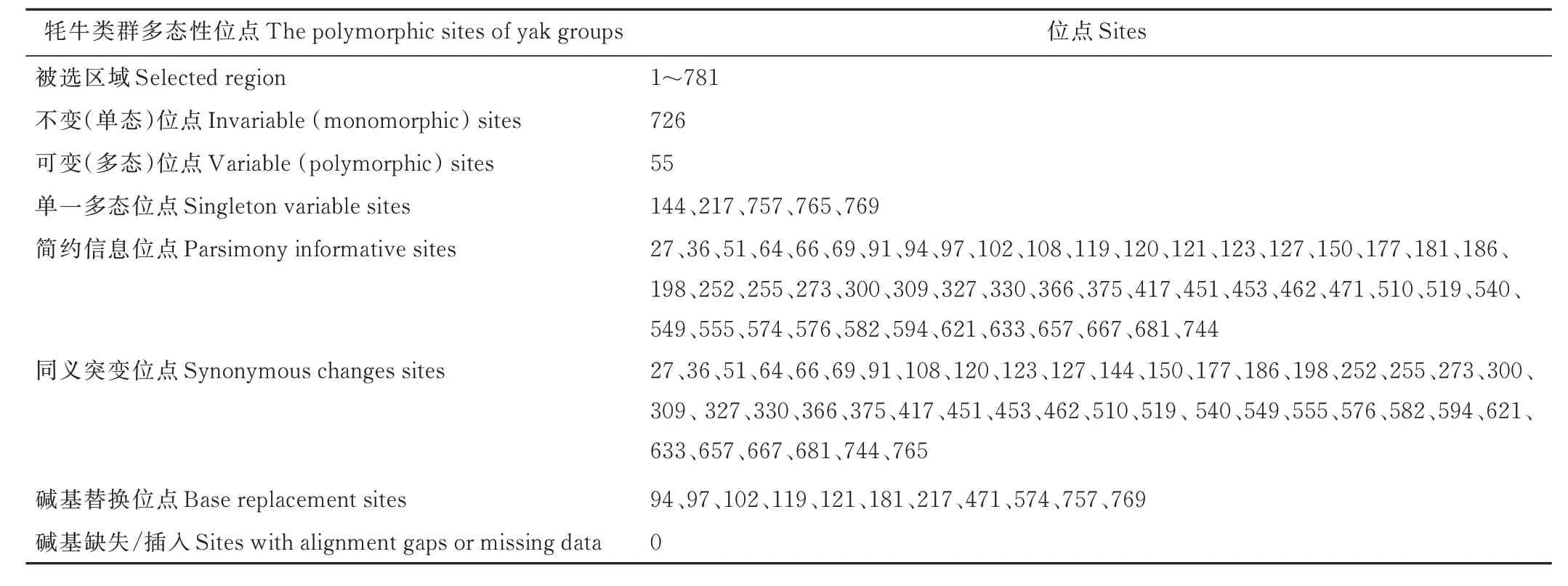
表2 西藏牦牛遺傳多樣性Table 2 Genetic diversity of Tibetan yak
經(jīng)分析得到7 個西藏牦牛群體COX3基因不同氨基酸的平均含量(表3),可以看出牦牛富含多種氨基酸,其中纈氨酸與亮氨酸平均含量較高,色氨酸平均含量最低(0.40%)。酸性氨基酸、堿性氨基酸的含量分別為4.84%、9.68%;親水性氨基酸、疏水性氨基酸的含量分別為35.08%、59.35%。
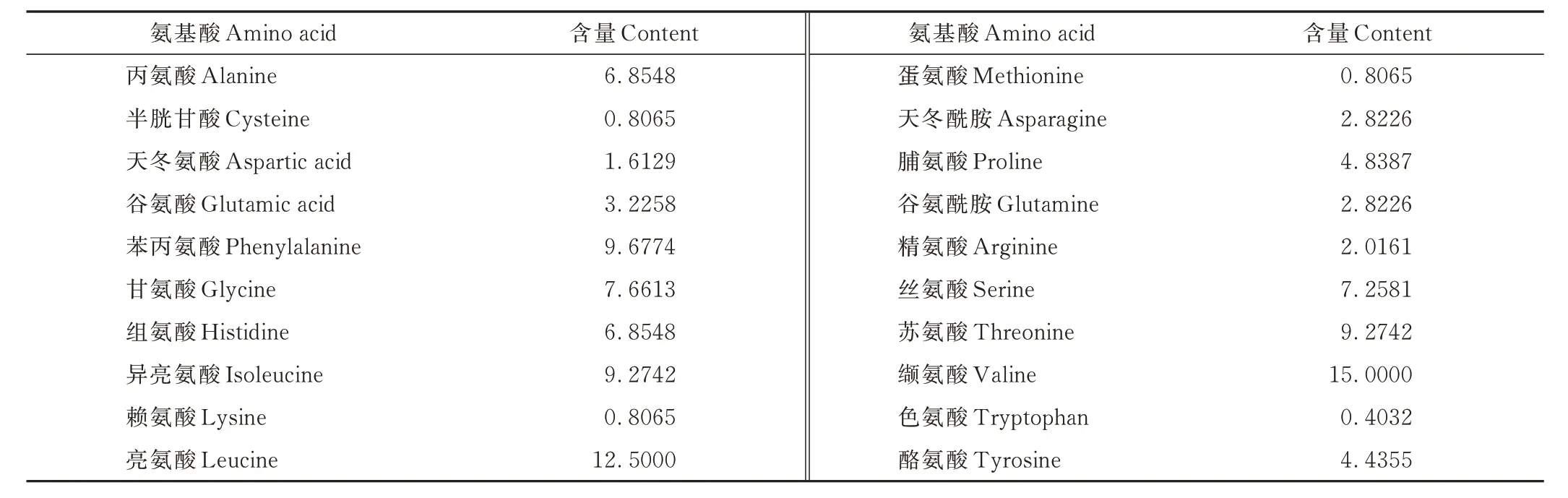
表3 西藏牦牛COX3 基因的氨基酸組成Table 3 Amino acid composition of COX3 gene of Tibetan yak (%)
2.2.2 核苷酸多樣性及單倍型組成 利用DNASP 軟件統(tǒng)計分析牦牛群體的核苷酸多樣性(表4)和單倍型多樣性(表5)。西藏牦牛COX3基因CDS 區(qū)共有11 種單倍型(表6),平均單倍型多樣性(average haplotype diversity,Hd)、平均核苷酸多樣性(average nucleotide diversity,Pi)分別為0.665 和0.00480,牦牛群體單倍型多樣性變化范圍為0.100~0.858;其中類烏齊牦牛最高,為0.858,嘉黎牦牛最低,為0.100;核苷酸多樣性為0.00013~0.01802;其中類烏齊牦牛最高,為0.01802,嘉黎牦牛最低,為0.00013。綜合核苷酸多樣性、單倍型多樣性以及單倍型數(shù)量來看,西藏牦牛群體遺傳多樣性豐富,且類烏齊牦牛的遺傳多樣性最高,而嘉黎牦牛的遺傳多樣性最低(表4 和表5)。西藏牦牛群體的Tajima’s D 值檢測結(jié)果顯示,工布江達牦牛和桑日牦牛值為正數(shù),其余牦牛群體均為負(fù)值,此外除帕里牦牛外,其余牦牛群體差異均不顯著(P>0.10)(表4)。
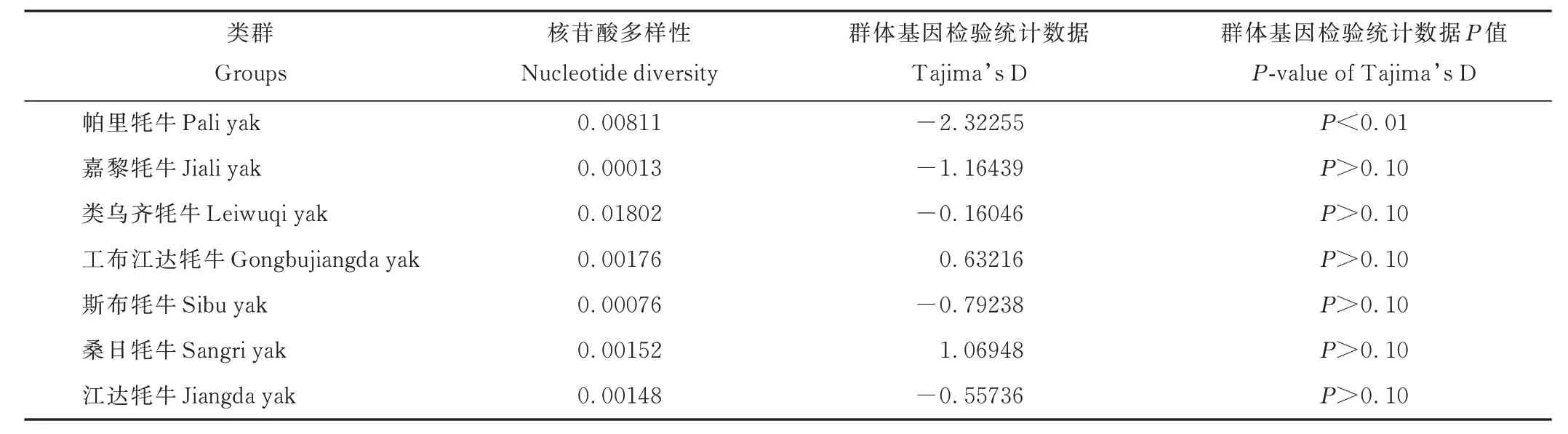
表4 西藏牦牛COX3 基因CDS 區(qū)核苷酸多樣性Table 4 Nucleotide diversity in CDS region of COX3 of Tibetan yak

表5 西藏牦牛群體的單倍型多樣性Table 5 Haplotype diversity of Tibetan yak groups
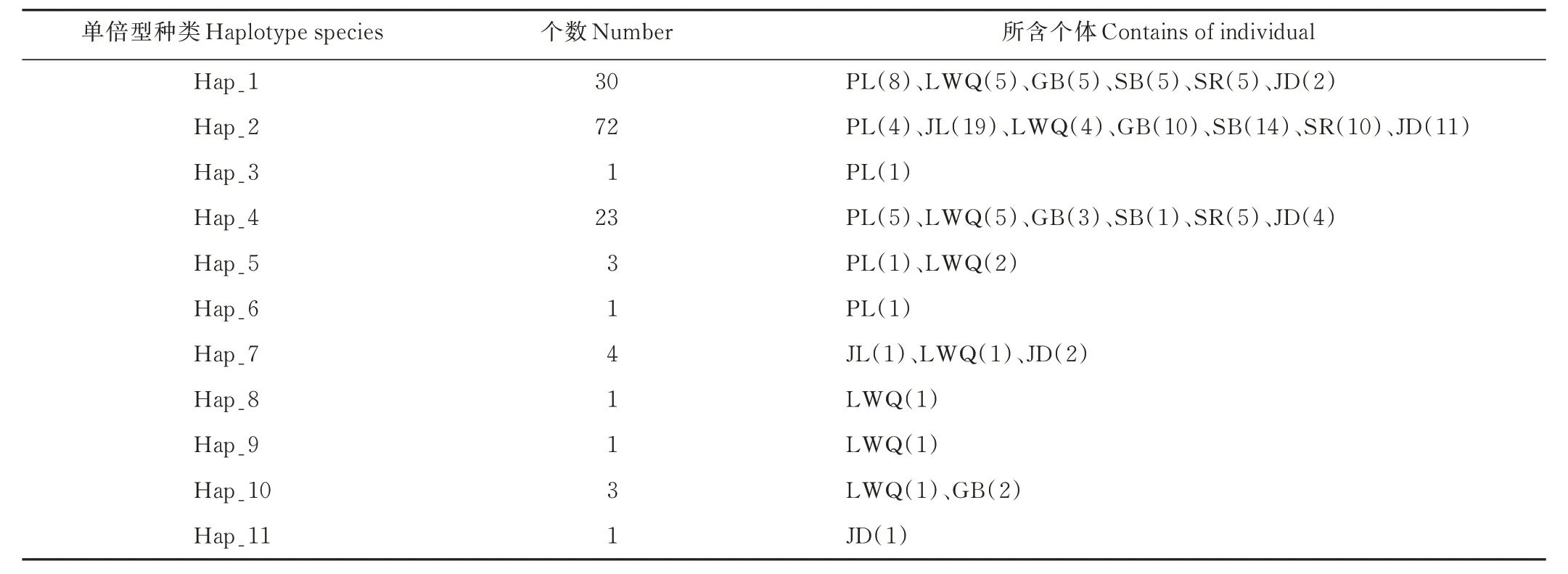
表6 西藏牦牛群體的單倍型數(shù)量分布Table 6 Haplotype distribution of Tibetan yak groups
統(tǒng)計分析各群體單倍型數(shù)量及分布情況(表6),Hap_2,Hap_1 及Hap_4 數(shù)量較多,分別為72、30 和23 個,幾乎分布于本研究的所有牦牛群體,其余單倍型占比較小,僅分布于少數(shù)幾個牦牛群體。類烏齊牦牛單倍型最多,共享8 種單倍型,其次是帕里牦牛,與其他地方品種共享6 種單倍型。
2.2.3 西藏牦牛群體間的遺傳距離及聚類分析 根據(jù)西藏牦牛mtDNACOX3基因CDS 區(qū)堿基序列,利用MEGA 7.0 基于Kimura-2-parameter 計算7 個牦牛群體間的遺傳距離。群體間遺傳距離的變異范圍為0.001~0.014,類烏齊牦牛和帕里牦牛間遺傳距離最大(表7),但本研究7 個西藏牦牛群體間的遺傳距離總體上均較小,表明它們之間的親緣關(guān)系較近,且存在基因交流。
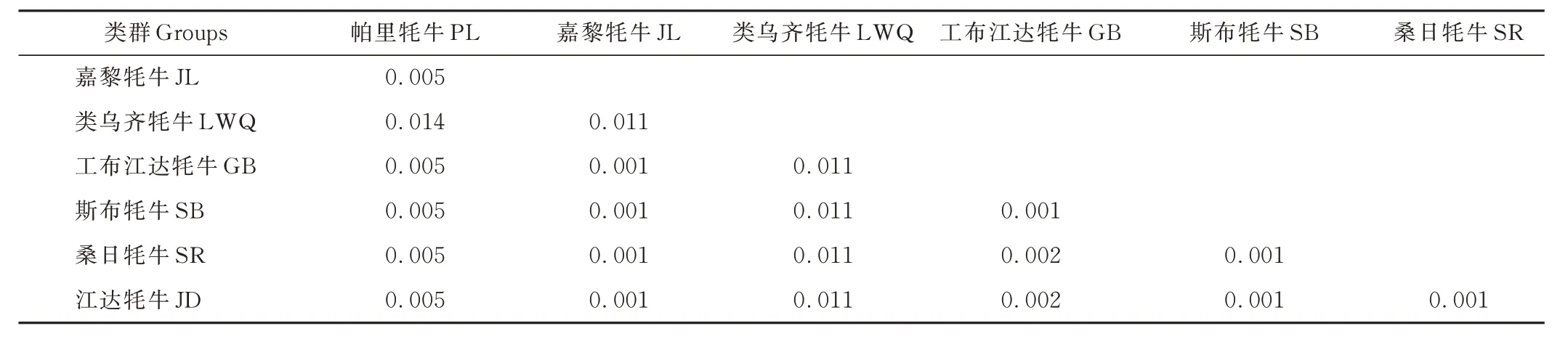
表7 西藏牦牛群體間Kimura 雙參數(shù)遺傳距離Table 7 Kimura two-parameter genetic distance among Tibetan yak groups
根據(jù)雙參數(shù)遺傳距離,對7 個西藏牦牛群體進行聚類分析,構(gòu)建系統(tǒng)發(fā)育樹(圖2)。結(jié)果表明,斯布牦牛與桑日牦牛、工布江達牦牛、帕里牦牛、嘉黎牦牛及江達牦牛聚為一類,類烏齊牦牛單獨聚為一類。
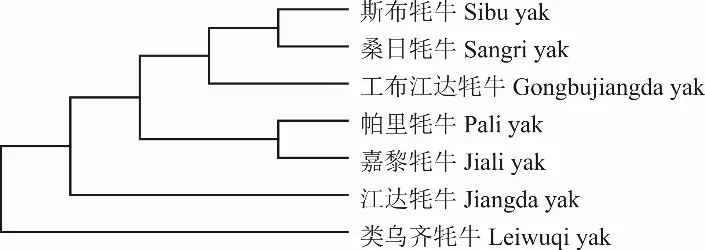
圖2 基于Kimura 雙參數(shù)遺傳距離的西藏牦牛類群間NJ 聚類關(guān)系Fig. 2 NJ tree based on Kimura 2-parameter distance among groups of Tibetan yak
2.2.4 系統(tǒng)發(fā)育樹及單倍型網(wǎng)絡(luò)關(guān)系 根據(jù)單倍型序列的變異位點,運用MEGA 7.0 軟件繪制西藏牦牛COX3基因CDS 區(qū)單倍型網(wǎng)絡(luò)關(guān)系圖及系統(tǒng)發(fā)育樹。11 種單倍型可分為2 個聚類簇,說明本研究的7個牦牛群體有兩個母系起源(圖3)。聚類簇Ⅰ包含9種單倍型,占全部單倍型的81.8%,涵蓋本研究中大部分牦牛群體,聚類簇Ⅱ包括2 種單倍型,占單倍型總數(shù)的18.2%。對單倍型個體構(gòu)建系統(tǒng)發(fā)育樹(圖4),發(fā) 現(xiàn)Hap_1、Hap_2、Hap_3、Hap_6、Hap_7、Hap_8、Hap_11 聚為一類后才與Hap_4 和Hap_10 聚為一大類,Hap_5 和Hap_9 聚為一類,其進化關(guān)系與單倍型聚類及分布狀態(tài)一致。
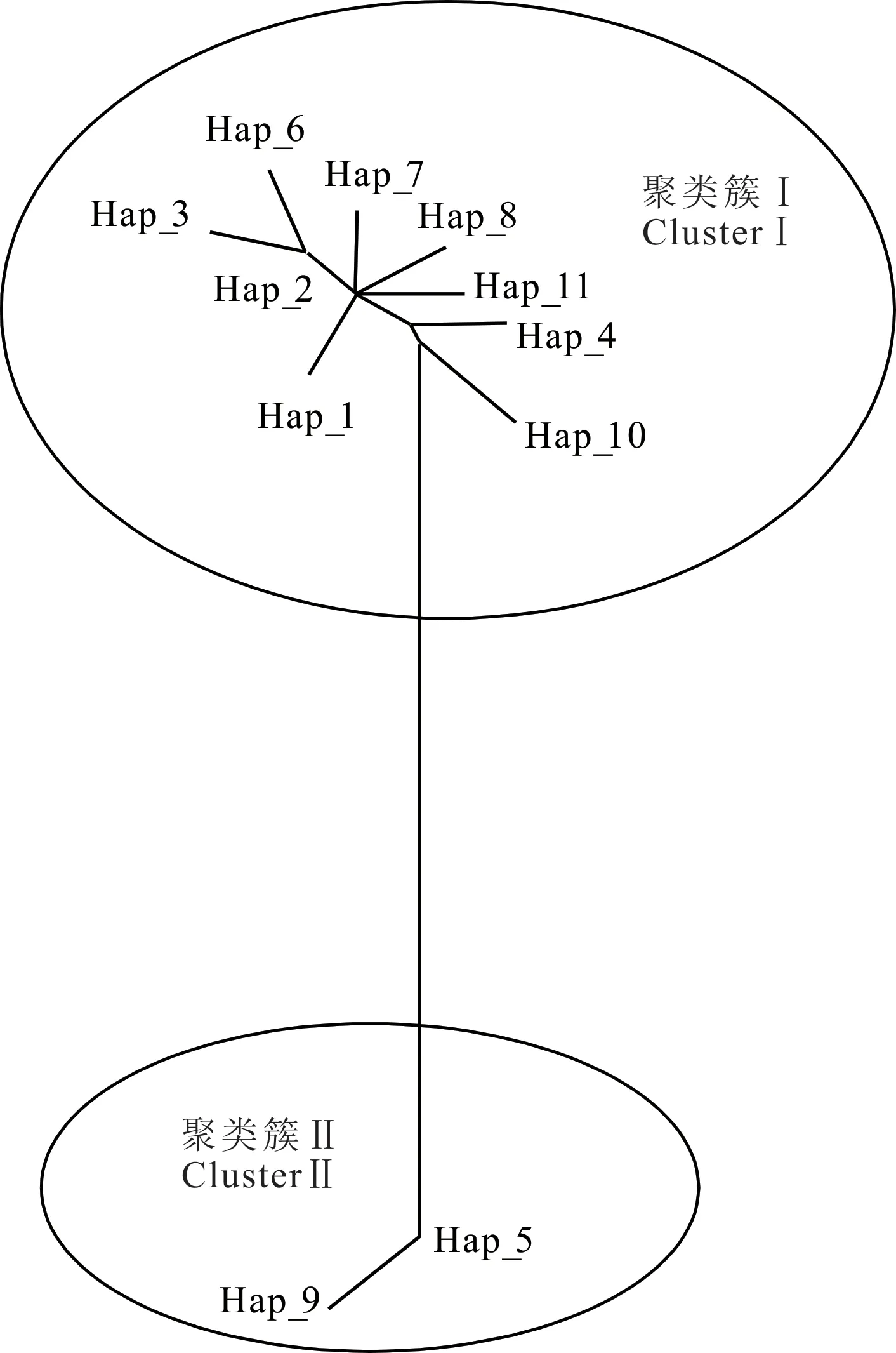
圖3 西藏牦牛mtDNA COX3 基因單倍型網(wǎng)絡(luò)關(guān)系Fig. 3 Network based on mtDNA COX3gene haplotype of Tibetan yak
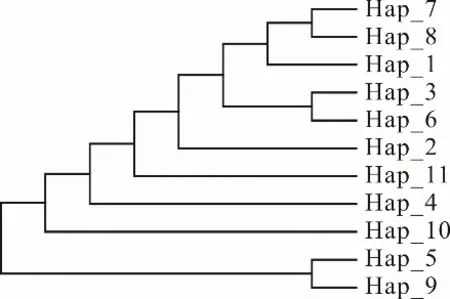
圖4 西藏牦牛11 種單倍型的系統(tǒng)發(fā)育樹Fig. 4 Phylogenetic trees based on 11 haplotypes of Tibetan yak
2.2.5 不同牛種間單倍型網(wǎng)絡(luò)進化關(guān)系 將從GenBank 下載的6 條其他牛種(家牦牛、野牦牛、印度野牛、白肢野牛、普通牛、原牛)的COX3基因序列作為對照,利用MEGA 7.0 軟件構(gòu)建了13 個群體COX3基因以及11 個單倍型間的系統(tǒng)發(fā)育樹。江達牦牛首先與家牦牛聚為一類,接著與本研究的其他牦牛群體聚為一大類,然后又與野牦牛聚為一大類,最后才與印度野牛、白肢野牛聚為一類(圖5)。本研究中的西藏牦牛基本上可劃分到家牦牛、原牛和普通牛三大單倍型群體中,其中8 個單倍型屬于家牦牛支系,2 個屬于原牛和普通牛支系,表明家牦牛、原牛和普通牛是西藏牦牛的混合母系起源,但受家牦牛影響較大,與NJ系統(tǒng)發(fā)育樹聚類結(jié)果一致(圖6)。
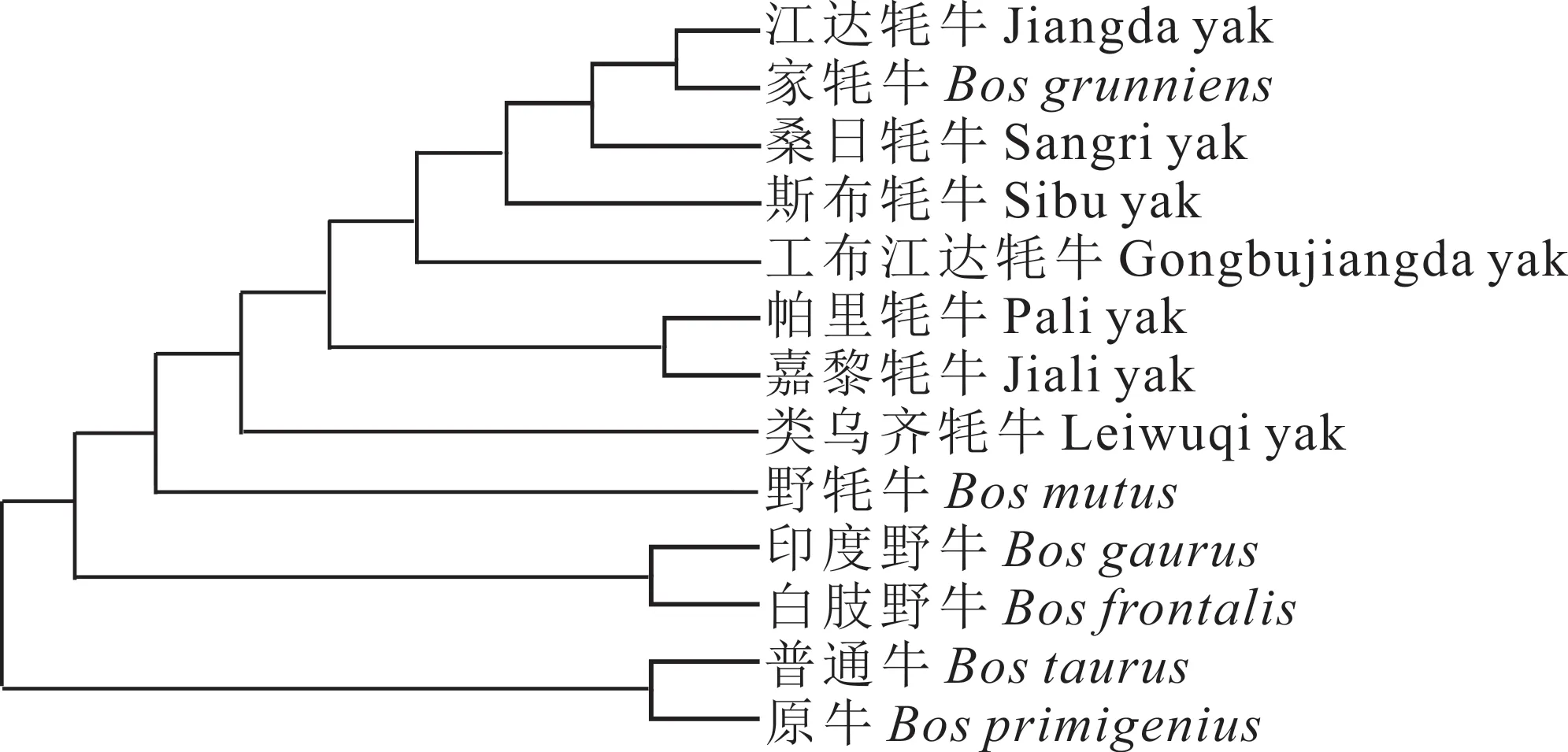
圖5 不同牛種COX3 基因的NJ 系統(tǒng)發(fā)育樹Fig. 5 Phylogenetic trees of COX3 gene based on NJ analysis in different cattle breeds
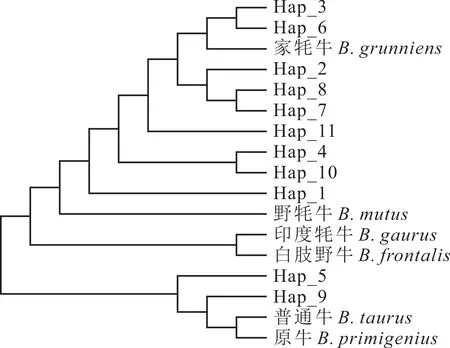
圖6 不同牛種COX3 基因單倍型的NJ 系統(tǒng)發(fā)育樹Fig. 6 Phylogenetic trees of COX3haplotype based on NJ analysis in different cattle breeds
3 討論
3.1 西藏牦牛mtDNA COX3 基因CDS 區(qū)序列遺傳多樣性分析
本研究測定了140 條西藏牦牛mtDNACOX3基因CDS 區(qū)序列,長度均為781 bp,序列中A+T 含量較G+C 含量高,說明西藏牦牛mtDNACOX3富含堿基AT,具有一定的堿基偏好性,符合大多數(shù)動物mtDNA AT 堿基含量偏高的普遍特征,屬于該基因序列中的優(yōu)勢堿基組成,這與杜玉杰等[7]對四川黑熊COXⅢ基因(A+T 含量56.4%),周武等[10]對豫西黑豬COXⅠ基因(A+T 含量59.3%)及謝雯琴等[11]對蛇源裂頭蚴COX3基因(A+T 含量65.13%~66.37%)A、T 堿基偏好的研究結(jié)果一致,這一結(jié)果與一般脊椎動物mtDNA 中 AT 含量在50%~63%堿基組成相吻合[12]。同時,相較于其他3 種堿基,G 堿基的含量最少(15.2%),表現(xiàn)出明顯的非G 堿基偏好性,這種非G 偏愛(anti-ghias)的特性是后生動物線粒體基因組L-鏈堿基組成所獨有的特點[13]。本研究西藏牦牛COX3基因富含多種氨基酸,其氨基酸組成與蛋白質(zhì)的質(zhì)量密切相關(guān),并受多種因素影響。動物機體中氨基酸平衡程度會直接影響動物的采食量,且動物體內(nèi)氨基酸供給處于平衡會降低動物尿氮損失從而提高氨基酸的利用效率,氨基酸不平衡會降低動物的生長率[14]。纈氨酸與亮氨酸是影響牦牛肉質(zhì)及牦牛乳組成特性的主要氨基酸[15-16],且纈氨酸與肉品香味有關(guān)[17],本研究中纈氨酸與亮氨酸含量較高,從側(cè)面反映出西藏牦牛肉質(zhì)風(fēng)味鮮美且乳成分營養(yǎng)價值較高。此外,通過對西藏牦牛COX3序列的比較分析,共發(fā)現(xiàn)11 種單倍型,55 個多態(tài)位點,群體間多態(tài)位點較豐富,變異較高的原因可能與牦牛所處環(huán)境有關(guān),紫外線可能是誘發(fā)突變的主要原因,亦或與部分牦牛群體受到的選擇壓力較小,積累的突變較多有關(guān)。通常認(rèn)為群體中的單倍型多樣性(Hd)、核苷酸多樣性(Pi)是衡量一個群體線粒體DNA 變異程度的兩個關(guān)鍵指標(biāo),Hd和Pi值越大,表明群體的遺傳多態(tài)性越豐富[18],本研究中西藏7 個牦牛群體的Hd和Pi分別為0.665 和0.00480,略低于姬秋梅等[1]對西藏牦牛Cytb基因的研究結(jié)果(Hd=0.884),但高于趙上娟等[2]對西藏牦牛CO Ⅲ基因(Pi=0.0018)以及李雙等[19]對溫嶺高峰牛線粒體DNA 全基因組(Hd=0.778,Pi=0.0017)的研究結(jié)果,可能與所研究的牦牛群體數(shù)量及參照基因不同有關(guān),但類烏齊牦牛的核苷酸多樣性和單倍型多樣性均高于其他牦牛群體,遺傳多樣性最豐富。Tajima’s D 值檢測結(jié)果顯示,工布江達牦牛與桑日牦牛值為正數(shù),說明其序列進化方式為平衡選擇,存在部分單倍型分化及群體收縮現(xiàn)象;其余牦牛群體均為負(fù)值,表明這些牦牛群體在進化過程中存在負(fù)向選擇或群體擴張現(xiàn)象。嘉黎牦牛、類烏齊牦牛、斯布牦牛以及江達牦牛群體的Tajima’s D 值均為負(fù)值,且中性檢驗結(jié)果不顯著,表明以上牦牛群體在進化過程中受到自然或者人工選擇后,群體發(fā)生了擴張現(xiàn)象,同時線粒體DNA序列在進化上遵循中性模型。以上結(jié)果均表明,西藏牦牛COX3基因序列的核苷酸替代率高,遺傳多樣性較為豐富。
3.2 西藏牦牛群體的系統(tǒng)進化關(guān)系分析
本研究根據(jù)Kimura 雙參數(shù)遺傳距離對西藏牦牛群體進行聚類分析,根據(jù)NJ 聚類關(guān)系,可將西藏7 個牦牛群體分為兩大類,即類烏齊牦牛單獨聚為一類,其余牦牛群體則聚為一類。相關(guān)研究表明,西藏牦牛群體的劃分與地理位置并不呈現(xiàn)明顯的相關(guān)性,不能單一地通過區(qū)域分布劃分,可能與牦牛的“單一起源”有關(guān)[20]。類烏齊牦牛在本研究7 個牦牛群體中遺傳多樣性最豐富,其次是帕里牦牛,此外,類烏齊牦牛也是本研究中單倍型最多的群體,結(jié)合西藏發(fā)展史和考古學(xué)等資料[21-22],類烏齊牦牛作為昌都市具有代表性的牦牛品種,隨著古羌人和古鮮卑人的融合,遷移以及現(xiàn)代西藏各民族的遷移,類烏齊牦牛也逐步走向青藏高原各地。本研究兩大分類與Chai等[23]利用全基因組測序?qū)θ珖?2 個家牦牛群體的進化和分化研究的分類結(jié)果一致,帕里牦牛、嘉黎牦牛、工布江達牦牛、斯布牦牛、桑日牦牛、江達牦牛聚為一類,主要分布于西藏的核心區(qū)域,包括西藏的中部、西部和南部;類烏齊牦牛聚為一類,主要分布于青藏高原的邊緣地帶。與姬秋梅等[1]基于mtDNAcytb基因及張成福等[24]根據(jù)mtDNA D-loop 區(qū)的分類結(jié)果因牦牛群體數(shù)量不同而存在一定差異。此外,帕里牦牛與其他地方群體共享6 種單倍型,僅次于類烏齊牦牛,表明帕里牦牛與其他地方品種或群體有著廣泛的基因交流,這與帕里牦牛作為西藏傳統(tǒng)的三大優(yōu)勢牦牛群體(嘉黎牦牛、帕里牦牛和斯布牦牛)之一,分布于西藏牧業(yè)發(fā)展的腹心地帶,牦牛引進覆蓋著周邊地區(qū)的實際情況一致。單倍型進化關(guān)系對比可以看出家牦牛、原牛和普通牛是本研究7 個西藏牦牛群體的混合母系起源,但受家牦牛影響較大,用最大似然法構(gòu)建的西藏牦牛系統(tǒng)進化樹與11 種單倍型的系統(tǒng)發(fā)育樹結(jié)果一致。
此外,研究者對mtDNACOX3基因作為物種系統(tǒng)分類依據(jù)的優(yōu)劣評價有所不同,可能與所研究的物種不同有關(guān)[5],也可能是由自然或人工選擇壓力下,不同DNA 序列的進化速率存在一定差異所致[25-26],有待進一步的深入研究。本研究也為mtDNACOX3基因作為系統(tǒng)分類候選標(biāo)記基因提供了一定參考。
3.3 西藏牦牛遺傳資源的保護與利用
牦牛作為“全能型”家畜,可為人們提供肉、乳、皮、毛、役、燃料等畜產(chǎn)品,是青藏高原主要的經(jīng)濟畜種。因此對西藏不同地區(qū)尤其是優(yōu)勢種的數(shù)量、性狀指標(biāo)等進行系統(tǒng)研究分析, 開展特色牦牛群體的種質(zhì)資源評價,適當(dāng)開展優(yōu)良品種選育、雜交優(yōu)勢利用等對牦牛遺傳多樣性的合理開發(fā)和可持續(xù)性利用尤為重要。
4 結(jié)論
本研究西藏7 個牦牛群體mtDNACOX3基因CDS 區(qū)序列遺傳多樣性豐富,其中類烏齊牦牛遺傳多樣性最高;單倍型可聚為2 簇,存在兩個母系起源;7 個牦牛群體可分為2 大類,即類烏齊牦牛(LWQ)為一類,其余牦牛為一類。西藏牦牛群體可劃分到家牦牛、原牛和普通牛三大單倍型群體中,其中8 個單倍型屬于家牦牛支系,2 個屬于原牛和普通牛支系。
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
天津外國語大學(xué)學(xué)報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
遼金歷史與考古(2019年0期)2020-01-06 07:45:20
民用飛機設(shè)計與研究(2019年4期)2019-05-21 07:21:24
電子制作(2018年11期)2018-08-04 03:26:04
汽車工程學(xué)報(2017年2期)2017-07-05 08:13:02
國際商務(wù)財會(2017年8期)2017-06-21 06:14:14
電子制作(2017年23期)2017-02-02 07:17:19
