玉米穗部性狀QTL定位與候選基因分析
2023-09-24 07:39:44劉鵬飛王安貴祝云芳陳澤輝郭向陽
種子 2023年7期
聶 蕾, 何 玥, 郭 爽, 王 棟, 涂 亮,劉鵬飛, 王安貴, 祝云芳, 吳 迅,3, 陳澤輝, 郭向陽
(1.貴州大學農學院, 貴陽 550025; 2.貴州省農業科學院旱糧研究所, 貴陽 550006;3.農業農村部喀斯特山區作物基因資源與種質創新重點實驗室, 貴陽 550006)
糧食安全已經成為世界各國普遍關注的問題,玉米是主要糧食作物之一,提高玉米產量對保障糧食安全起著至關重要的作用[1]。穗部性狀作為玉米產量的重要構成因子,包括穗長、穗粗、穗行數、穗重等,具有較產量性狀高的配合力和受環境影響程度小的優勢,在深度解析玉米產量性狀變異的遺傳基礎方面發揮著重要作用[2]。探討穗部性狀變異的遺傳基礎,對指導玉米育種、提高玉米產量具有十分重要的意義。例如,穗部性狀能夠預測玉米的產量,以便更好地利用種植期間的資源,最終達到高產的目的[3]。近年來,有關玉米穗部性狀QTL定位的研究取得了顯著進展,王輝等[4]以鄭58和HD568為親本構建的220份重組自交系群體為供試材料,在3個不同種植密度下共檢測到42個QTL,其中包括8個穗長、14個穗粗、14個穗行數和6個行粒數,單個QTL可解釋4.20%~14.07%的表型變異。宋曉恒等[5]通過豫82和豫87-1雜交構建的重組近交系(RIL),對玉米穗長、穗粗和籽粒深度進行QTL定位分析,共檢測到7個QTL,分布在1號、2號、4號、5號、8號染色體上。Shi等[6]以雜交種Xianyu335開發的240個優良品系所組成DH群體為構圖群體,基于964個SNP構建遺傳圖譜,在3種不同的生長條件下共鑒定出4個穗粗QTL,分別位于1號、3號、4號染色體上,可解釋表型變異率在4.7%~28.3%之間。白娜等[7]采用地方品種四路糯經作為供體親本,穗行數可達18~22行的農531作為輪回親本,通過完備區間作圖法(Inclusive composite interval mapping,ICIM)對穗行數QTL進行定位解析,在第5染色體136.3~140.0 Mb之間,定位到穗行數主效位點qKRN5.04,解釋了21.76%表型變異。趙強等[8]以T32和齊319為親本構建的雙親分離群體為材料,利用高密度SNP標記對5個玉米穗部相關性狀進行QTL定位,檢測到16個QTL,篩選出4個控制穗部的候選基因。Yan等[9]利用266份F2∶3家系為材料及150個SSR和26個RFLP分子標記進行QTL定位,共測得29個和單穗產量,百粒重、穗行數與行粒數相關QTL,單個位點可以解釋3.70%~16.80%的表型變異,其中11個位點在多環境中被檢測到。譚巍巍等[10]以黃早四為共同親本,分別以掖478和齊319雜交構建兩套F2∶3群體為試驗材料,在3個不同環境下,兩個群體分別檢測到33個和46個穗部相關性狀QTL。這些研究為深入揭示玉米產量性狀變異的遺傳基礎提供了豐富的科學支撐,但比較發現,不同研究者因為所用群體類型不同、大小差異以及標記等的不同,其研究結果也不盡相同。因此,本研究以兩個熱帶玉米骨干自交系(T32和QR273)為親本構建的F2、F2∶3家系為試驗材料,結合高通量的GBS測序技術和多環境下的穗部性狀評價結果,鑒定控制玉米穗部性狀相關遺傳區段,揭示其遺傳效應并開發功能性標記。在此基礎上,利用生物信息學分析方法分析控制穗部性狀變異的候選基因,為初步解析玉米穗部性狀變異的遺傳基礎提供更豐富的理論支撐,也為后續的精細定位、候選基因圖位克隆和分子標記輔助育種提供科學依據。
1 材料與方法
1.1 試驗材料
本研究使用的試驗材料選擇熱帶種質的玉米骨干玉米自交系QR273和T32,由貴州省農業科學院旱糧研究所提供。其中,QR273表現出配合力高、籽粒寬短,T32則表現為穗粗大,籽粒長厚,以這兩個自交系為親本構建的150份F2、F2∶3家系為作圖群體,后代的穗部性狀表現出顯著差異。
1.2 試驗方法
1.2.1田間試驗及性狀考察
2021年在海南種植150份F2分離后代,5葉期單株取樣提取基因組DNA,用于基因型鑒定,同時自交獲得F2∶3家系。2022年在貴州省貴陽市貴州省農業科學研究院試驗基地、甘肅省張掖市農業科學研究院試驗基地各栽植150個F2∶3家系及2個親本。試驗按隨機區組設計,重復2次,行長4 m,行距0.64 m,每行18株,田間管理按照當地玉米種植和管理方法執行。
玉米成熟期,每行連續采收所有果穗,自然風干至恒重,進行室內考種,每個材料隨機選取5株進行穗長、穗粗、穗行數、行粒數、禿尖長等穗部性狀的調查,參考石云素等[11]制定的《玉米種質資源描述規范與數據標準》。
1.2.2表型數據分析
應用 Microsoft Excel2010 軟件整理表型數據,采用SAS9.0軟件進行統計分析、方差分析、相關性分析及廣義遺傳力[12],遺傳力公式如下:
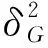
1.2.3基因型鑒定
DNA質量檢測和GBS測序委托北京康普生生物有限公司完成,具體操作依據吳迅等[13]描述的方法執行。即玉米F2植株5葉時期時,取各單株的幼嫩葉,采用改良CTAB法提取基因組DNA[14];利用簡化基因組測序技術(GBS)對每個單株的基因型進行鑒定。
1.2.4QTL分析
利用IciMapping4.1軟件中的完備區間作圖法對5個穗部性狀進行QTL檢測,隨機抽樣1 000例,QTL定位中閾值定為3。QTL命名格式為 q+性狀英文縮寫字母+染色體+序號。參考Stuber等[15]方法,對基因作用模式進行了估計,|d/a|=|顯性效應值/加性效應值|、A(加性方式):|d/a|=0.00-0.20;PD(部分顯性):|d/a|=0.21-0.80;D(顯性方式):|d/a|=0.81-1.20;OD(超顯性方式):|d/a|>1.20。
1.2.5候選基因預測
基于兩個環境下玉米穗部相關性狀的QTL定位結果,結合玉米B73參考基因組中的物理位置(Ref Gen_v4(https://www.maizeGDB.org/),綜合分析不同QTL的遺傳背景和效應,鑒定控制目標性狀的基因位點所在染色體區段,與公共數據庫和已發表的文獻QTL定位信息進行比較,尋找“一致性”QTL,并對該區段內的基因進行分析,預測候選基因。
2 結果與分析
2.1 穗部性狀表型分析
在兩個環境條件下,根據雙親(T32和QR273)與150個F2∶3家系5個穗部性狀表型的平均值,可以看出雙親在不同環境中的穗部性狀表現存在顯著差異。親本QR273在兩種環境中的穗長、行粒數的表現顯著大于親本T32,在貴陽單一環境下5個性狀表現均高于T32;而對張掖環境中的穗粗、穗行數和禿尖長而言,親本T32的表現優于QR273。F2∶3家系的穗長、穗行數和行粒數的均值介于雙親之間,為中親優勢;穗粗在兩個環境中的表現都較高,其均值大于雙親為超親優勢。5個穗部性狀偏度和峰度的絕對值均小于1,符合正態分布,可實現QTL定位分析。從遺傳力的研究結果中可看出,該群體遺傳力在不同環境中存在差異,如貴陽環境下5個穗部相關性狀的遺傳力高于張掖環境下,在貴陽穗粗遺傳力是61.87%,而在張掖遺傳力為11.11%(表1)。
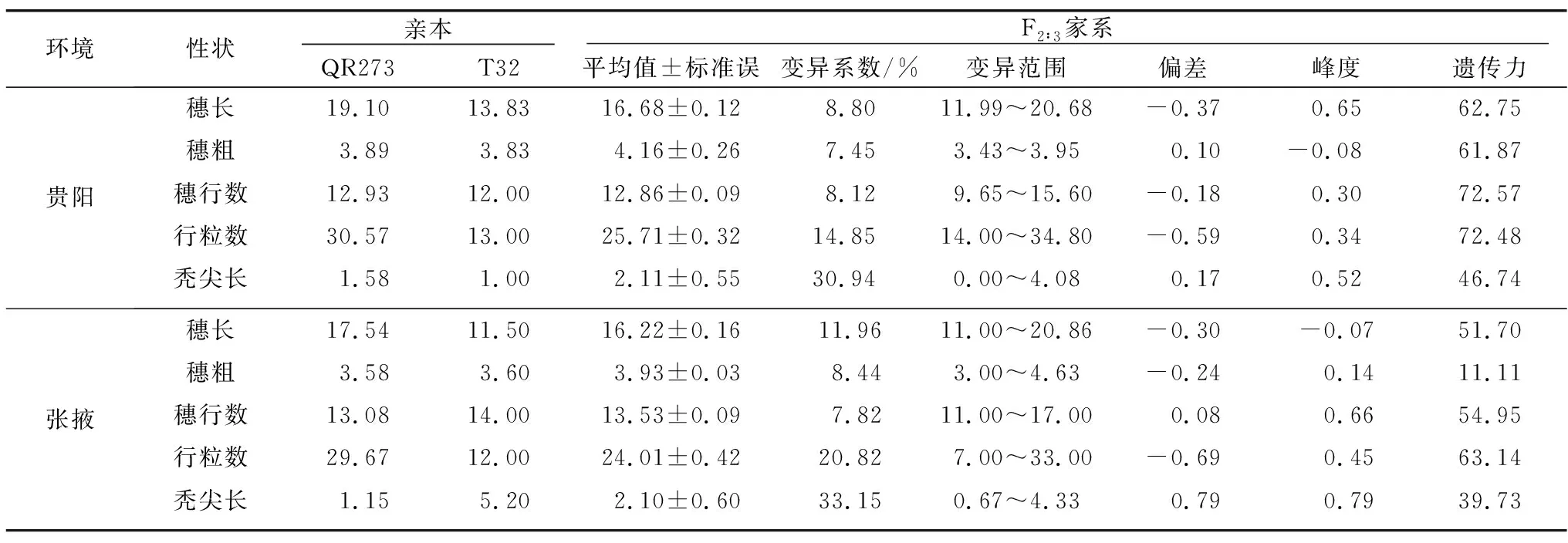
表1 親本和 F2∶3群體的穗部性狀表現Table 1 Performance of ear traits in parents and F2∶3 population
2.2 穗部性狀相關性分析
表2為穗部各個性狀之間相關分析。結果表明,在不同環境條件中,穗長和穗粗、行粒數、禿尖長都表現出顯著的正相關,穗長和穗行數在兩個環境中均表現為顯著負相關。穗粗除禿尖長外,與其他穗部性狀在兩個環境中呈顯著正相關。行粒數與貴陽環境中禿尖長之間存在極顯著的負相關,與張掖呈負相關。
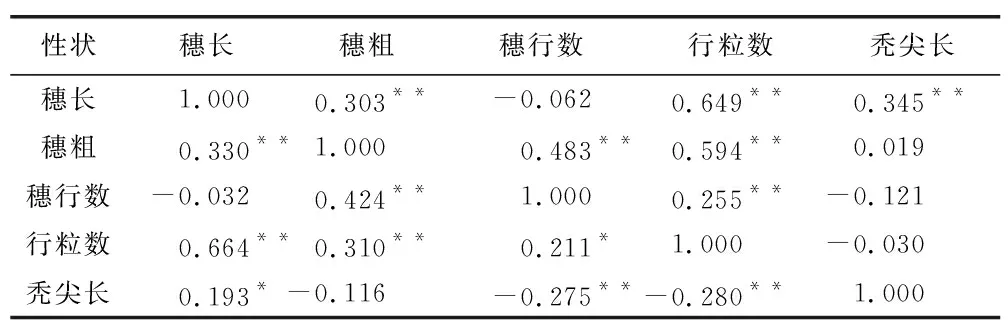
表2 穗部之間的相關性分析Table 2 Correlation analysis between ears
2.3 高密度物理圖譜構建
按照缺失率小于0.05,具有確定物理圖距為標準對獲得的10萬個SNPs進行過濾,最后獲得68 994個高質量SNPs,均勻地分布在10條染色體上(圖1)。各染色體上的標記數量大小依次表現為Ch4、Ch2、Ch1、Ch3、Ch5、Ch8、Ch9、Ch7、Ch6、Ch10,其數量分別為8 901個、8 824個、8 725個、7 623個、7 174個、6 407個、5 782個、5 592個、5 432個、4 534個。

注:每一行代表一個SNP標記,不同的顏色顯示不同的標記密度。淺色表示標記的密度較低,深色表示標記的密度較高。圖1 物理圖譜Fig.1 Physical map
2.4 穗部性狀QTL分析
由表3可知,兩個環境下共檢測到46個穗部相關性狀QTL,分布在10條染色體上,單個QTL可解釋表型變異范圍為1.89%~17.18%,其中,解釋表型變異大于10%的有6個,分別為qEL1-3、qED6-1、qED6-2、qERN3-1、qERN7和qEBT8-1。檢測到與穗長相關的QTL共有10個,分布在1號、2號、3號、4號、6號染色體上,單個QTL可解釋的表型變異3.35%~11.80%。在張掖環境下,位于1號染色體上的qEL1-3解釋的表型變異最大,其貢獻率為11.80%;穗粗在兩環境下共檢測到4個QTL,分布于1號、5號、6號染色體上,單個QTL可解釋的表型變異范圍為5.92%~11.03%;檢測到與穗行數有關的QTL共19個,分布在除了第1號和第9號染色體外的其他8條染色體上,單個QTL可解釋的表型變異范圍為1.89%~17.18%;檢測到與行粒數相關QTL共12個,分布在1號、2號、3號、5號、7號、9號、10號染色體上,單個可解釋表型變異范圍為3.50%~8.71%;檢測到與禿尖長相關QTL共5個,主要分布1號、2號和8號染色體上,單個QTL可解釋的表型變異范圍為2.16%~10.36%,qEBT8-1解釋的表型變異最大,為10.36%。就基因作用方式而言,5個穗部相關性狀的QTL表現不盡一致。其中4個QTL基因作用方式表現為加性效應;10個呈部分顯性;4個呈顯性;28個為超顯性。本研究多數與穗部性狀有關的QTL為部分顯性及超顯性,與Xiang等[16]、蘭進好等[17]的研究結果一致。在46個QTL中,增效等位基因來源于QR273的有17個,來自T32的有29個。

表3 5個穗部相關性狀QTL定位結果Table 3 QTL mapping results of 5 ear-related traits
2.5 一致性QTL
Tuberosa等[18]研究表明,對同一性狀QTL進行不同環境中的檢測,若其標記區間一致、增效等位基因起源一致則視為同一QTL。本研究通過對兩環境條件下的初步定位結果比較發現,qEL1-1和qEL1-2都在1號染色體13.07~13.09 Mb區間內被檢測到,增效等位基因來源于QR273;qERN5-1和qERN5-2都在5號染色體194.67~195.42 Mb區間內被檢測到,增效等位基因來源于QR273,這兩個在不同環境下均被檢測到的“一致性QTL”受環境影響較小,能夠穩定表達對于玉米產量的形成具有重要意義(表4)。
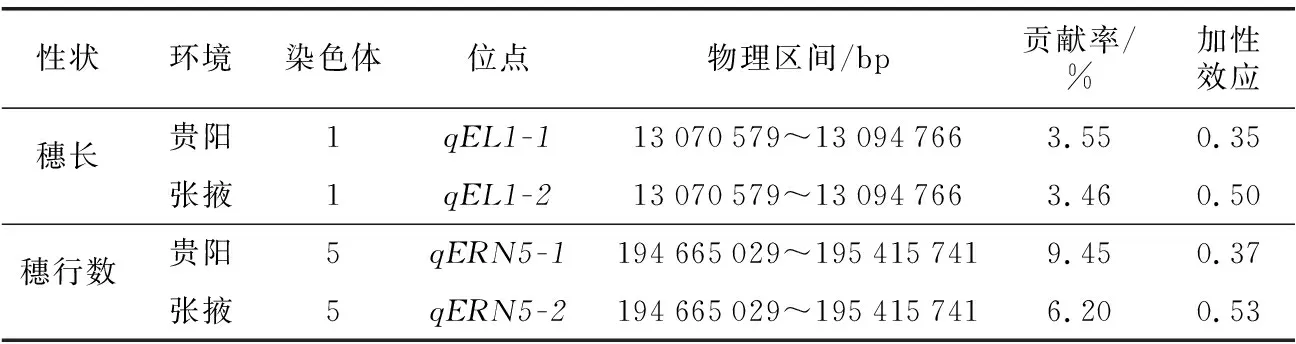
表4 一致性QTLTable 4 Consistency QTL
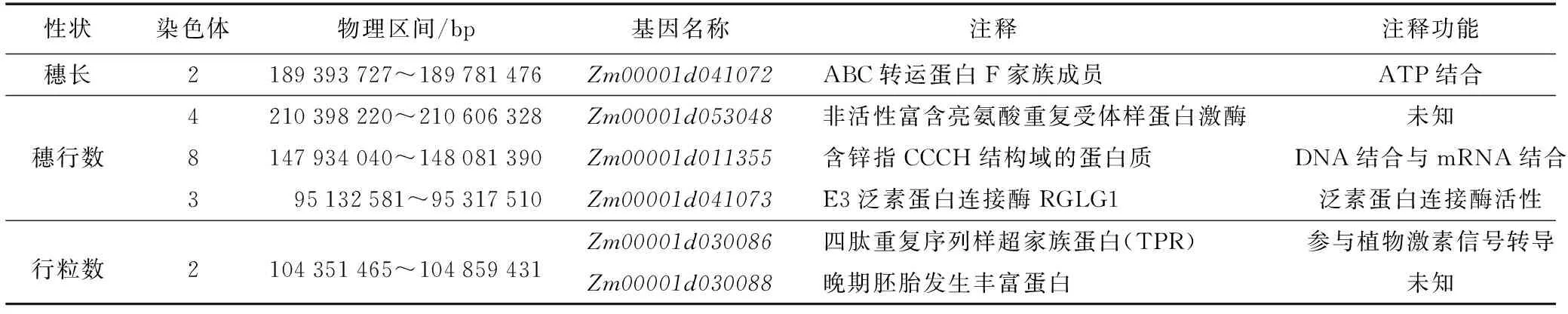
表5 基因位置及功能注釋Table 5 The gene location and functional annotation
2.6 候選基因分析
結合兩環境下的定位結果與已公開QTL數據庫對比發現,本研究檢測到46個QTL中有8個QTL與前人定位結果重疊,以篩選出的8個一致性QTL為目標,基于Gramene 網站數據庫(http://www.gramene.org)對目標候選基因進行分析,結合前人的研究報道,初步預測得到Zm00001d041072、Zm00001d053048、Zm00001d011355、Zm00001d041073、Zm00001d030086、Zm00001d030088等6個與穗部相關的候選基因。其中,Zm00001d041072是從控制穗長QTL區間發現,它編碼的ABC轉運蛋白在植物生長發育中發揮作用,因為植物生長發育需要多種信號分子來響應和溝通,這些信號分子跨膜轉運時需要ABC轉運蛋白[19];Zm00001d053048位于穗行數QTL區間內,其編碼的富含亮氨酸重復序列的類受體激酶為植物最大跨膜類受體激酶亞族,對植物發育及抗病反應具有廣泛的調控作用[20]。這些受體蛋白激酶是脅迫表達相關基因中的一部分,參與植物體對逆境的應答及防御相關過程;Zm00001d011355編碼的含鋅指CCCH結構域的蛋白質,CCCH型鋅指蛋白對植物生長發育及生物與非生物脅迫響應具有重要的調控作用。RNA能被該蛋白識別并結合[21];Zm00001d041073編碼的E3泛素蛋白連接酶RGLG1,2012年Cheng等[22]發現RGLG1、RGLG2對擬南芥對干旱脅迫的反應具有負調控作用,二者通過泛素化修飾ERF53,促進了它被26S蛋白酶體的降解,削弱植物對干旱的抗性等[22]。RGLG1,RGLG2也涉及生長素信號途徑、生長素信號轉導障礙,生長發育缺陷,對生長素轉運蛋白如PIN的含量有影響,對生長素的分配也有一定的影響[23]。Zm00001d030086是從控制行粒數的QTL區間內找到的,它所編碼的四肽重復序列樣超家族蛋白(TPR)是一類廣泛存在于植物體內的轉錄共抑制因子,參與多種植物激素信號轉導途徑,生物脅迫和非生物脅迫相關基因的轉錄抑制調控。擬南芥 TPR 除了參與激素信號轉導外,還參與調控植物的花期等生命過程[24];Zm00001d030088編碼的晚期胚胎發生豐富蛋白,該蛋白是一類較常見的當植物遭受干旱脅迫時產生的具有保護功能的蛋白[25]。有研究結果表明,LEA蛋白基因的表達能夠提高植物的耐旱性和抗寒性[26]。
3 討 論
玉米穗部性狀受到復雜基因網絡的調控,容易受到環境的影響。通過與前人研究結果分析比較,共發現8個控制玉米穗部相關性狀的一致性QTL。Zhou等[27]利用四交群體在1號染色體184.92~209.86 Mb(B73Ref-Gen_v4)區間內定位到1個穗長QTL,本研究在1號染色體189.39~189.78 Mb和188.85~189.35 Mb區間內定位到2個控制穗長的QTL,分別為qEL2和qEL1-3;譚巍巍等[10]在3號染色體94.27~141.39 Mb和10號染色體60.38~90.58 Mb區間內分別定位到控制穗行數的qKRE2-3-2和qKRE1-10-1,本研究在相鄰位置95.13~95.31 Mb和83.88~85.11 Mb區間內分別定位到控制穗行數的qERN3-1和qERN10;張君[28]在8號染色體137.81~148.01 Mb區間內定位到控制穗行數的qERN8b,本研究在相鄰位置147.93~148.08 Mb區間內定位到控制穗行數的qERN8;王輝等[4]在4號染色體215.09~205.41 Mb區間內定位到一個控制穗行數的QTL為qERN4,本研究在4號染色體207.41~210.61 Mb區間內定位到兩個控制穗行數的QTL,分別為qERN4-1和qERN4-2;王輝等[4]在2號染色體105.71~103.87 Mb區間內定位到一個控制行粒數的QTL為qKNR4,本研究在2號染色體104.35~104.85 Mb區間內定位到控制行粒數的QTL為qKNR2-3。除此之外,本研究在1號染色體上檢測到的QTL較多,與劉琳[29]的研究結果一致,在5號、6號、7號、9號染色體上定位到一些新的穗部性狀QTL。
隨著分子生物學的日益發展,愈來愈多的研究者發現植物激素合成以及一些重要轉錄因子和蛋白激酶對玉米生長發育發揮重要作用[30]。趙強等[8]基于高密度SNP標記對玉米穗部相關性狀的候選基因分析,在穗長、穗行數和禿尖長區間內預測出4個候選基因,與穗行數qERN1位點關聯的候選基因Zm00001d027721,其編碼的高親和力鎳轉運家族蛋白具有運輸功能,本研究與行粒數qKNR2-3位點關聯的候選基因Zm00001d030086也具有參與植物激素信號轉導的功能。Zhou等[27]通過QTL定位與全基因組關聯分析(GWAS)確定了3個控制穗長QTL的候選基因,其編碼的蛋白與植物生長素運輸、細胞增殖和發育調節有關。本研究從控制穗長、穗行數和行粒數的QTL區間內預測了6個可能影響穗部性狀的基因,其編碼的蛋白具有合成激素,運輸和信號傳導的功能,發揮的具體作用在擬南芥等植物中得到了驗證。
4 結 論
本研究以熱帶種質自交系為親本構建雙親分離群體作為試驗材料,通過QTL定位與候選基因分析,共定位到46個穗部性狀QTL。與已發表文獻及公開QTL數據庫進行對比,發現8個與前人研究結果重合,分別控制穗長、穗行數和行粒數的變異,其中1個穗長QTL和1個穗行數QTL在多環境下均被檢測到,且區間完全重疊。研究結果在前人定位結果的基礎上,進一步減小QTL置信區間,為進一步的精細定位提供候選靶標。本研究共預測得到6個候選基因,分別是:Zm00001d041072、Zm00001d053048、Zm00001d011355、Zm00001d041073、Zm00001d030086、Zm00001d030088,其編碼的蛋白具有合成激素等生物學功能,可作為后續基因圖位克隆和分子輔助育種的候選靶標。
