5個羅氏沼蝦群體微衛星遺傳多樣性分析
2020-11-05 09:22:26慎佩晶張宇飛李喜蓮徐洋程海華陳雪峰
浙江農業科學 2020年11期
慎佩晶,張宇飛,李喜蓮,徐洋,程海華,陳雪峰
(農業農村部淡水漁業健康養殖重點實驗室浙江省淡水水產遺傳育種重點實驗室 浙江省淡水水產研究所,浙江 湖州 313001)
羅氏沼蝦(Macrobrachiumrosenbergii),又稱大頭蝦、馬來西亞大蝦,隸屬于節肢動物門,軟甲綱,十足目,長臂蝦科,沼蝦屬,原產于東南亞,是目前世界上重要的淡水養殖蝦類之一,也是世界上最大的淡水蝦。它在水生態系統中分布較廣,在淡水區域(如江河、湖泊)和半咸水區域(如河口)均有分布,對水環境中的理化指標變化反應較敏感[1]。它具有生長速度快、食性較廣、生長周期短和抗病能力強等特點,自1976年該蝦引入我國以來,已在浙江、江蘇、上海、廣東、廣西、湖南、湖北等十多個省市自治區養殖推廣。
目前,用于蝦類遺傳多樣性研究的分子標記方法較多。限制性片段長度多態性(RFLP)[2]、隨機擴增多態性DNA(RAPD)[3]、簡單重復序列標記(SSR)[4]、ISSR[5]、EST-SSR[6]、相關序列多態性(SRAP)[7]、擴增片段長度多態性(AFLP)[8]等多種標記技術已在蝦類多樣性研究中被采用。其中SSR由2~6個核苷酸為重復單元組成的DNA序列,在真核生物的基因組中分布廣泛。這一序列具有共顯性、多態性相對豐富、基因組覆蓋高等優點。該技術具有易于操作、周期短、等位基因多樣性高和穩定性高等特點[9],使得SSR標記成為基因組研究中的一種主要的分子標記,被廣泛用于遺傳圖譜構建[10]、品種鑒定[11]、輔助選擇育種[12]、親緣關系鑒定[13]、遺傳多樣性分析[14]和系統發育學[15]等研究中。SSR技術也在羅氏沼蝦上得到廣泛應用。王傳聰等[16]對羅氏沼蝦肝胰腺組織轉錄組測序共檢測出15 356個SSR位點。蔣飛等[17]應用SSR標記對羅氏沼蝦5個選擇系子二代共630尾蝦的遺傳多樣性和遺傳分化進行了研究。戴習林等[18]利用SSR對羅氏沼蝦3個養殖群體進行遺傳多樣性分析,并探討了最適宜的樣本量及標記量。
本研究選用5個群體127份羅氏沼蝦樣品,對羅氏沼蝦種質進行SSR分子標記遺傳多樣性分析,可以有效挖掘特異的羅氏沼蝦種質材料,促進種質材料的有效利用,這對改良現有品種,提高羅氏沼蝦產量和品質,具有極其重要的意義,也為分子標記輔助育種提供理論依據。
1 材料與方法
1.1 樣品采集
試驗材料為羅氏沼蝦南太湖2號,從緬甸與孟加拉引種的緬甸群體和孟加拉群體,2018年新引進的以色列超雌蝦和泰國蝦群體,取自浙江省淡水水產研究所羅氏沼蝦長興保種基地。其中,南太湖2號為連續第10代選育的核心群,緬甸群體為從緬甸引進的野生種經馴化養殖的第5代,孟加拉群體為從孟加拉引進野生種經馴化養殖的第7代。
分別從以色列群體(I)、緬甸群體(R)、孟加拉群體(B)、核心群體(C)、泰國群體(T)中隨機選取8、30、28、30、31個羅氏沼蝦個體,采集其腹部肌肉置于無水乙醇中固定,-20 ℃保存備用。
1.2 基因組DNA提取
羅氏沼蝦肌肉組織基因組DNA提取采用天根生物公司植物基因組提取試劑盒(DP320)[17],具體步驟按說明書進行。采用NanoDrop2000C分光光度計檢測DNA質量和濃度,-20 ℃保存備用。
1.3 微衛星引物設計與合成
9對微衛星引物均為本實驗室自行開發的SSR引物(表1),由南京金斯瑞生物科技有限公司合成,并在正向引物5′端添加FAM熒光標記。
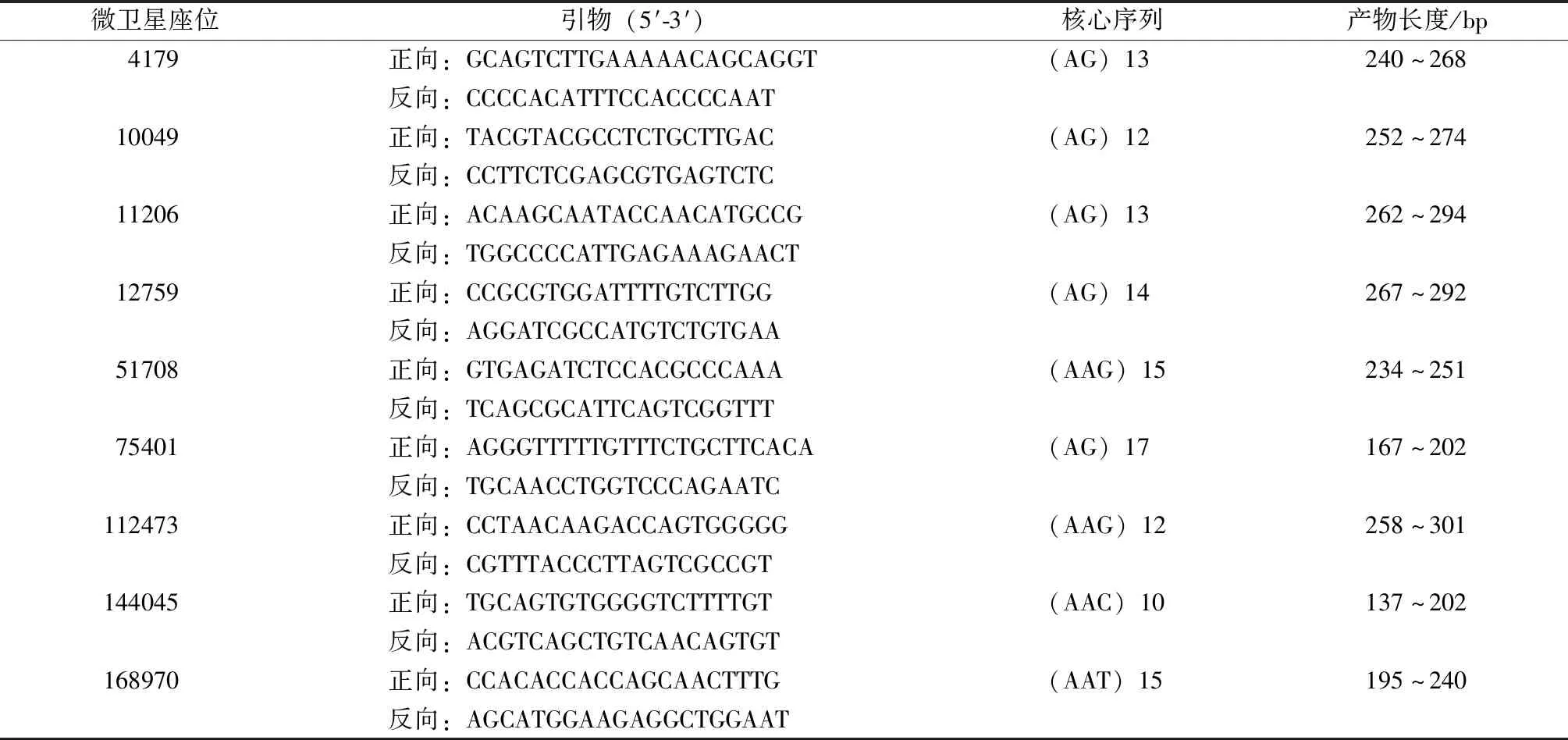
表1 微衛星引物的信息
1.4 PCR擴增
以這9對引物分別對127個樣品DNA進行PCR擴增,反應體系為20 μL:10×buffer 2 μL,dNTPs(2.5 mmol·L-1)0.5 μL,熒光引物(10 μmol·L-1)各1 μL,DNA模板約50 ng,Taq酶(5 U·μL-1)0.5 μL,加ddH2O至20 μL。PCR反應程序為:94 ℃預變性5 min;94 ℃變性30 s,合適的退火溫度下退火30 s,72 ℃延伸30 s(35個循環);72 ℃延伸7 min。各對引物的最適退火溫度通過梯度PCR試驗確定。PCR產物純化后,在乙醇已經揮發完全的板中加入內標LIZ500和HiDi,振蕩充分混勻,瞬時離心以除去管壁樣品。放入PCR儀,95 ℃ 4 min變性。SSR-PCR擴增產物放到3730XL中進行毛細管電泳。
1.5 數據處理與分析
使用Gene Mapper V3.0軟件對基因掃描文件進行分析,記錄每個位點的相對分子質量大小,得到相應的位點的片段大小和信號值。根據ROX-500標準,自動校準尺寸和峰值。采用Popgen1.32軟件計算微衛星座位的有效等位基因數(Ne)、等位基因數(Na)、表觀雜合度(Ho)、Shannon’s信息指數(I)、期望雜合度(He)、基因流(Nm)、多態信息含量(polymorphism information content,PIC)、F-統計量(Fis、Fit、Fst)、群體遺傳距離等相關參數,并構建UPGMA聚類圖。
2 結果與分析
2.1 9個微衛星座位遺傳多態性
采用篩選出的9對引物對127份羅氏沼蝦種質材料的遺傳多樣性進行分析,9對引物均具有多態性,擴增出的條帶在137~301 bp(如圖1為引物11206在羅氏沼蝦1、16、28號樣品中的擴增結果),共檢測到70個等位位點,每個引物檢測到5~10個,平均為7.777 8個。其中,引物4179檢測到的等位位點最多,有10個;引物168970檢測的等位位點次之,有9個。
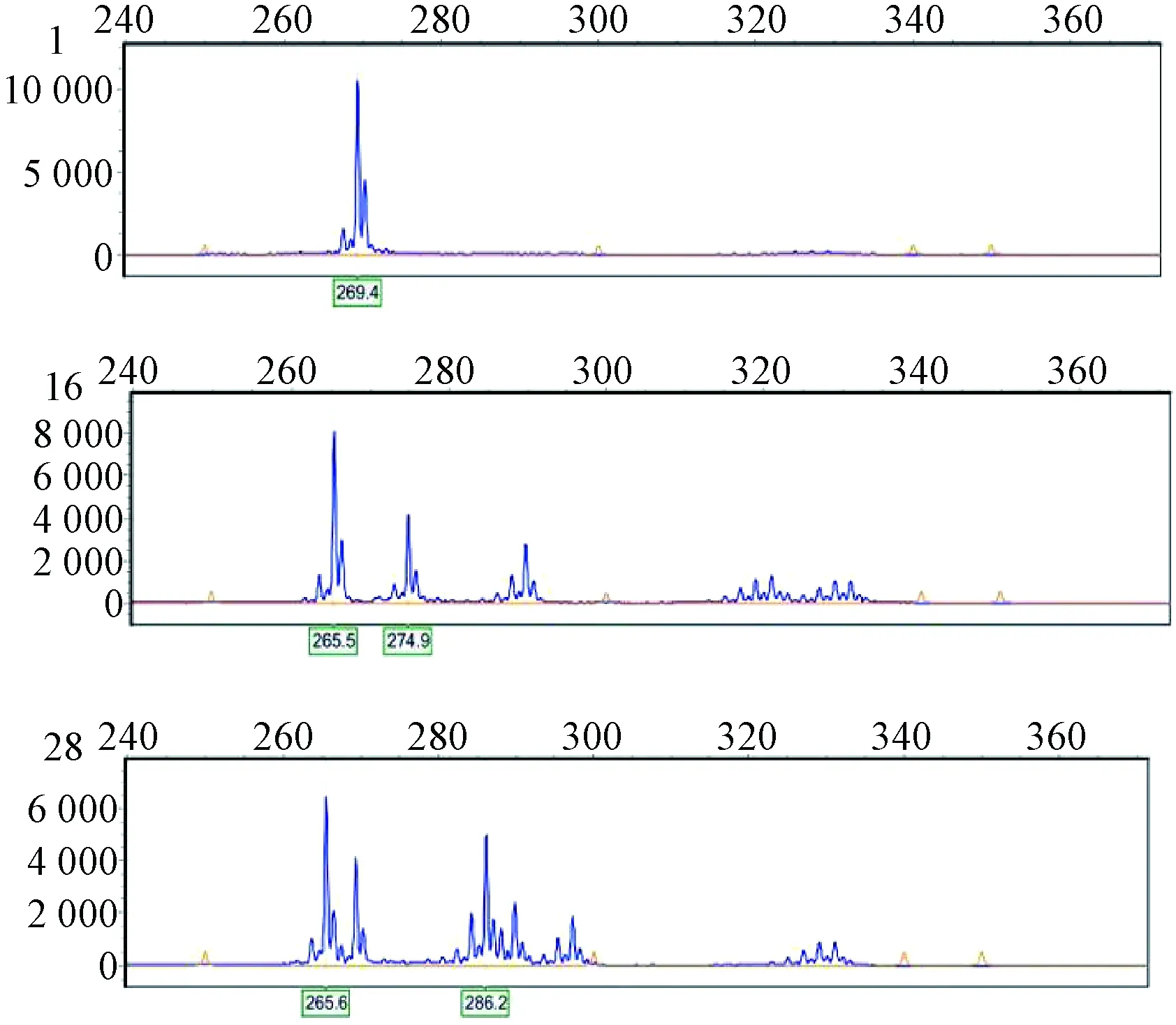
圖1 引物11206在羅氏沼蝦1、16、28號樣品中的基因分型
各座位的表觀雜合度在0.149 6~0.637 8,群體均值為(0.325 6±0.173 6);期望雜合度在0.362 2~0.850 4,群體均值為(0.674 4±0.173 6);9對引物的多態信息量在0.607 4~0.844 9,平均值為0.828 0,其中引物168970的多態信息量最高,為0.844 9,引物10049的多態信息量最低,僅為0.607 4。9對引物的PIC均大于0.5,說明這些標記能提供較高可信度的信息(表2)。

表2 羅氏沼蝦5個地理群體9個微衛星位點的遺傳多樣性
2.2 群體內遺傳多態性
9對SSR引物均擴增出清晰的條帶,羅氏沼蝦種質資源的等位基因(Na)平均為2.777 8~6.666 7。以色列群體最少,泰國群體最多,5個種質資源總體等位基因數平均為5.044 5。有效等位基因(Ne)平均為2.078 8~3.930 5;以色列群體最少,泰國群體最多,5個種質來源總體平均為3.131 4。5個群體PIC值平均在0.453 7~0.548 3,以色列群體最低,泰國群體最高。PIC為衡量基因變異程度高低的多態性信息量指標[19],PIC>0.50時,該引物為高度多態性信息引物;0.25≤PIC≤0.50時,該引物為中度多態性信息引物;當PIC<0.25時,該引物為低度多態性信息引物。核心群體及泰國群體具有較高的遺傳多樣性;以色列群體、緬甸群體和孟加拉群體具有中度的遺傳多樣性(表3)。
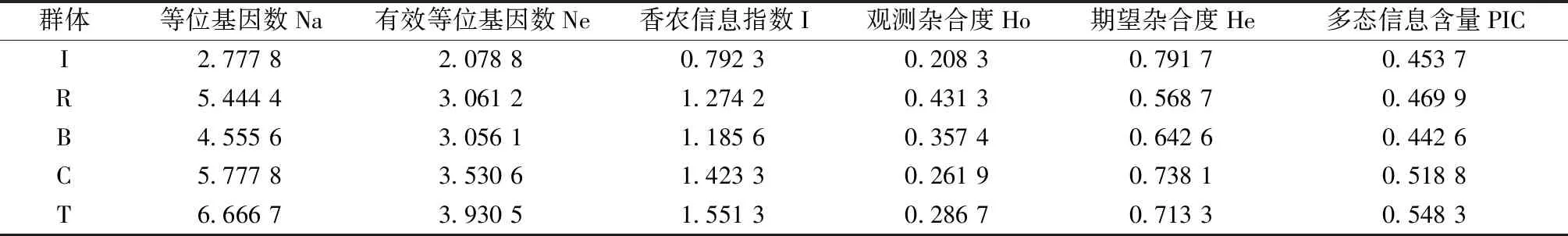
表3 羅氏沼蝦5個群體的遺傳多樣性
2.3 微衛星座位遺傳分化
對羅氏沼蝦5個群體的Fit、Fis、Nm統計顯示,Fis在微衛星位點12759、112473和168970上表現為正值,分別為0.162 9、0.093 7和0.411 9,其余位點均為負值,總體平均為-0.079 3。Fit在位點4179、10049、11206和51708為負值,分別是-0.136 8、-0.221 0、-0.007 0和-0.312 8,平均值為0.105 8。各位點的基因流(Nm)在0.661 5~4.406 0,其中≥1的位點有7個(4179、10049、11206、51708、75401、14405和168970),占所有檢測位點的77.78%,均值為1.207 7(表4)。
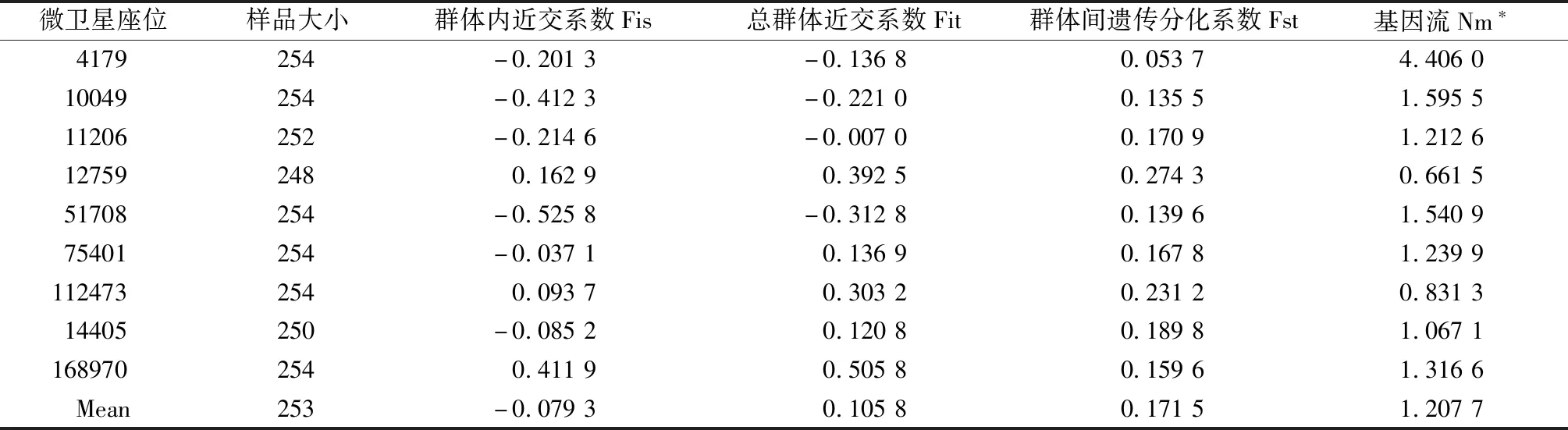
表4 微衛星座位的F值和基因流
2.4 群體間遺傳距離
5個群體間遺傳距離范圍在0.227 6~1.001 2,其中遺傳距離最大的是孟加拉群體與以色列群體之間的距離為1.001 2;遺傳距離最小的為孟加拉群體與核心群體之間的距離為0.227 6;平均遺傳距離為0.619 4(表5),這與前人的研究結果[20]相比,本研究的遺傳多樣性更為豐富。這可能與本研究采集的群體樣品來源地跨度大、分布廣有關。

表5 Nei氏遺傳距離和遺傳相似性
2.5 群體遺傳結構
采用Popgen1.32軟件,對5個群體的微衛星遺傳多樣性進行分析,以遺傳相似系數為基礎進行UPGMA聚類,繪制的遺傳聚類樹狀圖能直觀表現個體間親緣。由圖2可知,孟加拉群體與核心群體首先聚為一支,再與以色列群體與緬甸群體聚成的一支聚合,聚類最遠的為泰國群體。
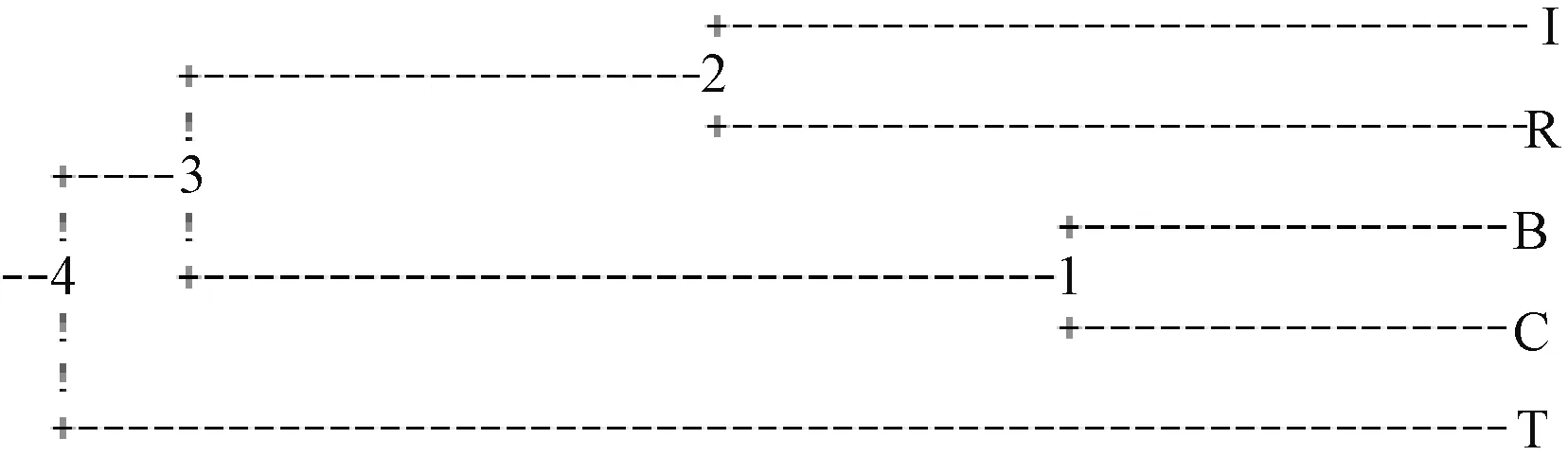
圖2 羅氏沼蝦5個群體的UPGMA聚類
3 討論
種質資源是育種的基礎,育種材料的遺傳多樣性能提高育種水平,作為種質評價和利用的基礎。遺傳多樣性是物種長期生存和保持進化的基礎[21],也可為基因資源的發掘提供必要的信息[22]。群體遺傳結構的研究對物種的保護、利用和改良具有重要的理論和實踐意義。
我國自1976年引進羅氏沼蝦,由于長期近親間和在有限的小群體交配繁殖,使這些羅氏沼蝦種群基因流失,出現退化現象,抗病性與生長能力都在降低[23]。因此,對羅氏沼蝦的群體遺傳結構和遺傳多樣性的研究成為確定優先保護種群、有效的取樣策略和近交衰退種群的恢復策略的制定都有極為重要的意義[24-25]。周勁松[26]對羅氏沼蝦緬甸引進種和浙江本地種及其雜交種進行SRAP分析,并利用RAPD技術分析了羅氏沼蝦單對交配親本及F1代的遺傳關系及分離規律。李明云等[27]利用RAPD技術,對浙江省羅氏沼蝦養殖群體和緬甸群體的遺傳差異進行了分析,發現緬甸自然群體與浙江養殖群體遺傳距離為0.184 5。甘西等[23]采用RAPD技術,對馬來西亞引進后代的養殖群體(簡稱NY)和新加坡F1代與養殖群體交配繁殖的后代(簡稱NX)的遺傳多樣性進行了研究,Nx群體內的遺傳變異度明顯高于Ny群體。
本研究采用9對SSR引物對羅氏沼蝦5個群體的127個樣品進行遺傳多樣性及遺傳結構分析,共得到70個等位位點,平均多態信息含量(PIC)為0.828 0;群體結構分析結果顯示,5個種群遺傳變異豐富,可以為新品種的創制提供幫助。
