饑餓脅迫對擬穴青蟹大眼幼體的比較轉(zhuǎn)錄組分析
2023-09-14 11:46:04胡偉勝馬凌波張鳳英劉志強馬春艷
海洋漁業(yè) 2023年4期
胡偉勝,王 偉,馬凌波,張鳳英,劉志強,趙 明,馬春艷
(1.中國水產(chǎn)科學(xué)研究院東海水產(chǎn)研究所遺傳育種與生物技術(shù)實驗室,上海 200090;2.上海海洋大學(xué)水產(chǎn)與生命學(xué)院,水產(chǎn)科學(xué)國家級實驗教學(xué)示范中心,上海 201306)
自然界中,食物匱乏是甲殼類時常面臨的環(huán)境脅迫之一。甲殼類面對短期或長期的營養(yǎng)限制時經(jīng)常通過調(diào)整代謝、行為和消耗內(nèi)部儲存的氨基酸、脂質(zhì)和碳水化合物以維持基本的生理功能并實現(xiàn)有限的生長和繁殖[1-7]。饑餓脅迫下,顆粒新厚蟹(Neohelicegranulata)肌肉組織的氨基酸代謝和肝胰腺的甘油異生發(fā)生明顯改變[1-2]。近期的研究表明,饑餓脅迫激活了中華絨螯蟹(Eriocheirsinensis)肝胰腺的脂質(zhì)代謝和糖原代謝[3]。饑餓壓力下,艾氏濱蟹(Carcinus aestuarii)免疫功能顯著改變[4],凡納濱對蝦(Litopenaeusvannamei)免疫酶的活性顯著升高[5]。饑餓壓力下,甲殼類行為也會發(fā)生適應(yīng)性改變。例如,在饑餓壓力下,三疣梭子蟹(Portunustrituberculatus)和日本蟳(Charybdis japonica)的捕食率呈先升后降的趨勢[6],凡納濱對蝦的游泳速度明顯放緩[7]。因甲殼動物的腸道微生物群落與宿主的消化和免疫功能緊密相關(guān)[8-9],在饑餓狀態(tài)下的凡納濱對蝦腸道微生物的多樣性和群落結(jié)構(gòu)與健康蝦雖無統(tǒng)計學(xué)差異,但是饑餓蝦的腸道中機會病原體豐度更高,導(dǎo)致蝦生病幾率大大增加[5]。
擬穴青蟹(Scyllaparamamosain)俗稱青蟹,隸屬于節(jié)肢動物門(Arthropoda),甲殼綱(Crustacea),十足目(Decapoda),梭子蟹科(Portunidae),青蟹屬。屬廣溫性和廣鹽性海洋經(jīng)濟甲殼動物,是我國東南沿海地區(qū)重要的海水養(yǎng)殖蟹類。迄今為止,關(guān)于梭子蟹科的饑餓脅迫研究主要側(cè)重于表觀指標的呈現(xiàn)。例如,饑餓后復(fù)投喂降低了鋸緣青蟹[10]和三疣梭子蟹[11]Ⅰ期溞狀幼體的生長指標和存活率。對于饑餓脅迫分子機理的研究目前相對較少見,特別是幼體階段的個體。本文對正常投喂和饑餓狀態(tài)下的擬穴青蟹大眼幼體作比較轉(zhuǎn)錄組分析,以探究饑餓脅迫對擬穴青蟹大眼幼體代謝過程的影響,以期為擬穴青蟹苗種的健康培育提供理論參考,并豐富擬穴青蟹的分子生物學(xué)基礎(chǔ)數(shù)據(jù)。
1 材料與方法
1.1 樣本采集
實驗用蟹采自中國水產(chǎn)科學(xué)研究院東海水產(chǎn)研究所浙江寧海研究中心,正式實驗前將即將蛻殼的Ⅴ期溞狀幼體從育苗池轉(zhuǎn)移至塑料圓柱形桶(底面半徑25 cm,高80 cm)中,待其順利蛻殼后撈取大眼幼體開展后續(xù)實驗。本實驗選用的大眼幼體來源于課題組培育的同一親本“東方1號”。實驗期間用1 L的玻璃燒杯養(yǎng)殖大眼幼體,保持溫度27.5~30.0℃、鹽度24~26、pH 8.0~8.4,餌料為孵化的豐年蟲無節(jié)幼體。每天上午8∶00采用滴管滴入的方式投喂餌料。共設(shè)2個處理組,分別為對照組(正常投喂3 d組,F(xiàn)3S0)和饑餓組(饑餓3 d組,S3F0),每組3個重復(fù),每個重復(fù)8只大眼幼體。處理3 d后采用滴管吸入的方式采集大眼幼體,并將其置于RNA保存液中4℃過夜,-80℃保存。
1.2 RNA提取
8只大眼幼體作為一個樣品,用TRIzol法提取樣品RNA,利用Nanodrop 2000超微量分光光度計(Thermo,美國)檢測RNA濃度和純度,用瓊脂糖凝膠電泳檢測RNA完整性,用Agilent 2100生物分析儀(Agilent,美國)測定RNA完整值(RNA integrity number,RIN)。OD260/280介于1.9~2.1。
1.3 文庫構(gòu)建
利用帶有Oligo(dT)的磁珠與mRNA上的poly-A結(jié)合純化mRNA,采用離子打斷的方式,將mRNA分成200~300 bp的片段。以mRNA作為模板,反轉(zhuǎn)錄合成cDNA第一鏈,再以第一鏈cDNA為模板合成雙鏈cDNA。雙鏈cDNA經(jīng)末端修復(fù)、加接頭、連接、純化、PCR擴增和cDNA回收構(gòu)建cDNA文庫。使用Agilent 2100質(zhì)檢文庫,利用Pico green檢測文庫濃度,用qRT-PCR定量檢測有效文庫。文庫基于Illumina HiSeq測序平臺測序(Illumina HiSeq 3000)。
1.4 轉(zhuǎn)錄組De novo組裝
Illumina測序得到原始序列(raw reads)。對原始序列進行數(shù)據(jù)過濾,將原始序列中的接頭序列和低質(zhì)量序列去除,獲得高質(zhì)量序列(clean reads)。使用Trinity軟件對高質(zhì)量序列進行拼接得到轉(zhuǎn)錄本和unigene。為獲得全面的基因功能信息,將拼接得到的unigene與NR、GO、KEGG、Pfam、eggNOG和Swissprot等數(shù)據(jù)庫進行比對注釋。
1.5 差異基因表達
采用DESeq軟件分析基因表達差異,將篩選條件設(shè)定為:|log2FoldChange|>1,顯著性P<0.05。使用topGO對差異基因進行GO富集分析。利用KOBAS數(shù)據(jù)庫(KOBAS網(wǎng)址:http://kobas.cbi.pku.edu.cn/)對差異基因進行KEGG富集分析和KO分析。當矯正后的P值<0.05時,認為該功能顯著富集。
1.6 實時熒光定量PCR(qRT-PCR)驗證
為了驗證測序結(jié)果的準確度,實驗隨機挑選了Met、MIH、E75、α-amylase和Trypsin基因進行qRT-PCR驗證。利用NCBI Primer-Blast軟件設(shè)計熒光定量引物,內(nèi)參選擇18S rRNA(表1)。引物送至上海杰李生物技術(shù)有限公司合成。使用ABI QuantStudioTM6 Flex System 儀和TaKaRa SYBR Premix ExTaqⅡ試劑分析F3S0和S3F0差異基因的表達水平。反應(yīng)體系采用TaKaRa SYBR Premix ExTaqⅡ說明書中20μL體系標準,PCR反應(yīng)程序為:95℃30 s;95℃5 s,60℃34 s,40個循環(huán);95℃15 s,60℃1 min,95℃15 s。Met、MIH、E75、α-amylase和Trypsin基因的相對表達分析采用2-△△Ct法。數(shù)據(jù)處理使用SPSS26.0 軟件進行單因素方差分析(one-way ANOVA)和Duncan檢驗,顯著性水平設(shè)置為P<0.05。
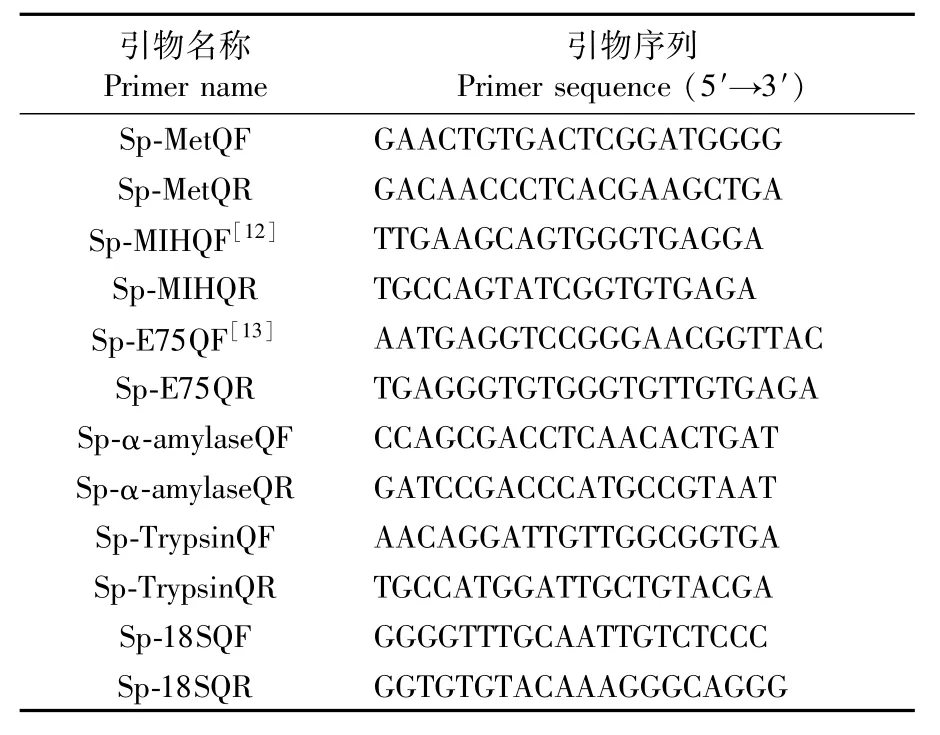
表1 qRT-PCR引物序列Tab.1 Primers for quantitative RT-PCR
2 結(jié)果與分析
2.1 轉(zhuǎn)錄組測序和De novo組裝
對正常投喂和饑餓3 d的擬穴青蟹大眼幼體進行測序,分別獲得162 469 532和139 222 024條原始序列。原始數(shù)據(jù)過濾(去除接頭序列和低質(zhì)量序列)后分別獲得 150 103 944 和128 924 890條高質(zhì)量序列(表2)。使用Trinity軟件對高質(zhì)量序列做De novo組裝,獲得158 201個轉(zhuǎn)錄本。將轉(zhuǎn)錄本去冗余后獲得79 581個unigene,最大長度20 651 bp,平均長度1 065.44 bp(表3)。后續(xù)分析全部以unigene序列作為參考序列。
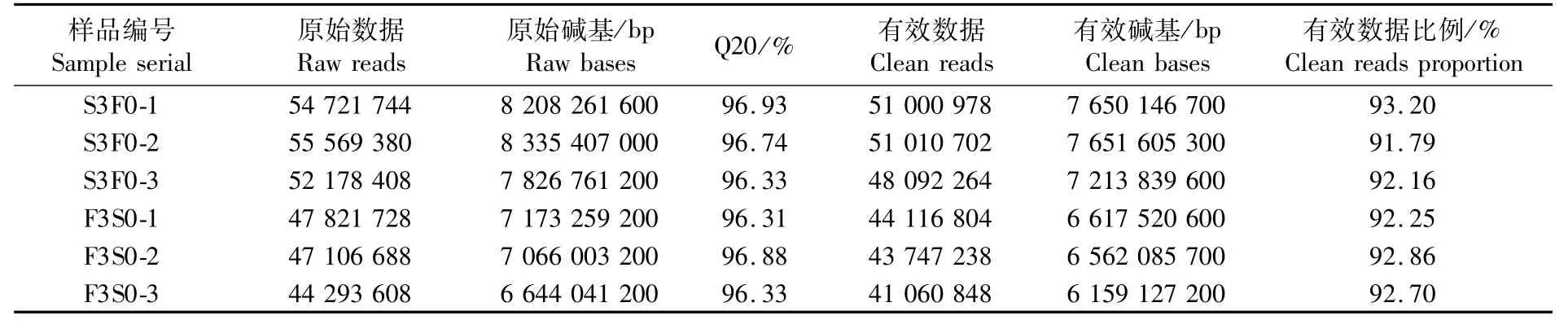
表2 轉(zhuǎn)錄組測序數(shù)據(jù)統(tǒng)計Tab.2 Statistic summary of sequencing data

表3 De novo組裝結(jié)果統(tǒng)計Tab.3 Statistic summary of De novo assembly data
2.2 功能注釋
Nr、GO、KEGG、Pfam、EggNOG和Swissprot數(shù)據(jù)庫的注釋信息統(tǒng)計結(jié)果見表4。注釋到GO數(shù)據(jù)庫的unigene共7 294個(表4)。GO功能注釋分為三大類:生物過程(biological process)、細胞組分(cellular component)和分子功能(molecular function)。生物過程中,代謝過程(GO:0008152 metabolic process)、細胞過程(GO:0009987 cellular process)和單一生物過程(GO:0044699 single-organism process)是主要的子類;細胞組分中,細胞(GO:0005623 cell)、細胞部分(GO:0044464 cell part)、細胞器(GO:004322 6 organelle)和膜(GO:0016020 membrane)是主要的子類;分子功能中,催化活性(GO:0003824 catalytic activity)和結(jié)合(GO:0005488 binding)是主要的子類(圖1)。

圖1 GO分類功能注釋Fig.1 Functional classification annotation of GO database
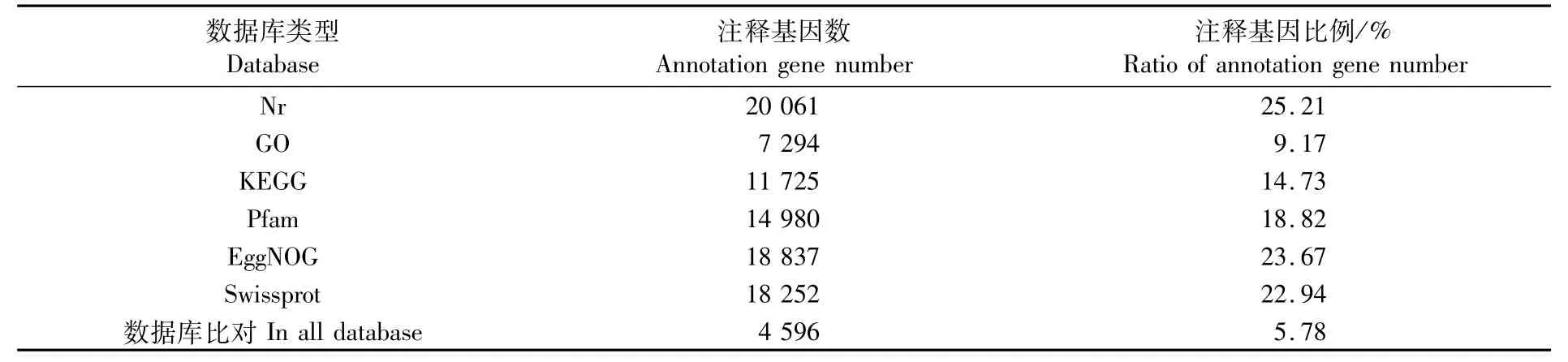
表4 基因注釋數(shù)量統(tǒng)計Tab.4 Statistic summ ary of annotation gene number
注釋到KEGG數(shù)據(jù)庫的unigene共11 725個(表4),level1分為5類,即代謝(metabolism)、遺傳信息處理(genetic information processing)、環(huán)境信息處理(environmental information processing)、細胞過程(cellular process)和有機系統(tǒng)(organismal system),其注釋的unigene數(shù)量分別為4 055、2 272、1 823、2 112和3 602個(圖2)。
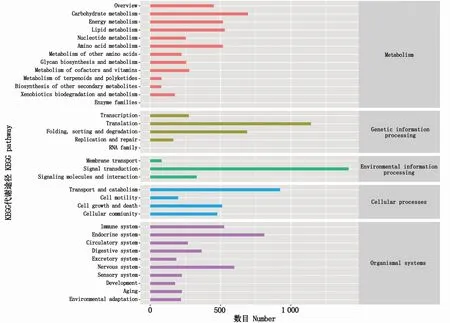
圖2 KEGG分類注釋Fig.2 Functional classification annotation of KEGG database
2.3 表達差異分析和富集分析
轉(zhuǎn)錄組測序共獲得5 621個差異表達基因,饑餓組與對照組相比,上調(diào)2 641個,下調(diào)2 980個。GO富集分析顯示,共254條GO Term得到顯著性富集(矯正后的P值小于0.05)。本文僅列出富集最顯著的前20個GO Term(圖3),其中前5條GO Term 分別是催化活性(catalytic activity)、碳水化合物代謝過程(carbohydrate metabolic process)、氧化還原過程(oxidationreduction process)、 氧 化 還 原 酶 活 性(oxidoreductase activity)和DNA復(fù)制起始(DNA replication initiation)。
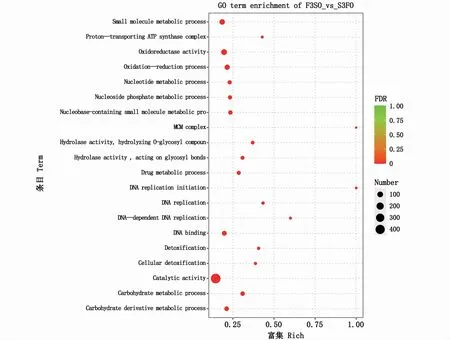
圖3 差異表達基因富集最顯著的前20條GO TermFig.3 Top 20 significantly enriched GO term s based on differentially expressed genes
KEGG 富集分析顯示,共 60 條 KEGG pathway得到顯著性富集(矯正后的P值小于0.05)。本文僅列出富集最顯著的前20條KEGG pathway(圖4),其中前5條KEGG pathway分別是DNA復(fù)制(DNA replication)、纈氨酸、亮氨酸和異亮氨酸降解(valine,leucine and isoleucine degradation)、細胞周期(cell cycle)、淀粉和蔗糖代謝(starch and sucrosemetabolism)、堿基切除修復(fù)(base excision repair)。值得注意的是,DNA復(fù)制通路富集的21個差異表達基因全部下調(diào)(圖5)。此外,KEGG顯著富集的通路中氨基酸代謝、脂質(zhì)代謝和碳水化合物代謝的占比相對較大(表5)。
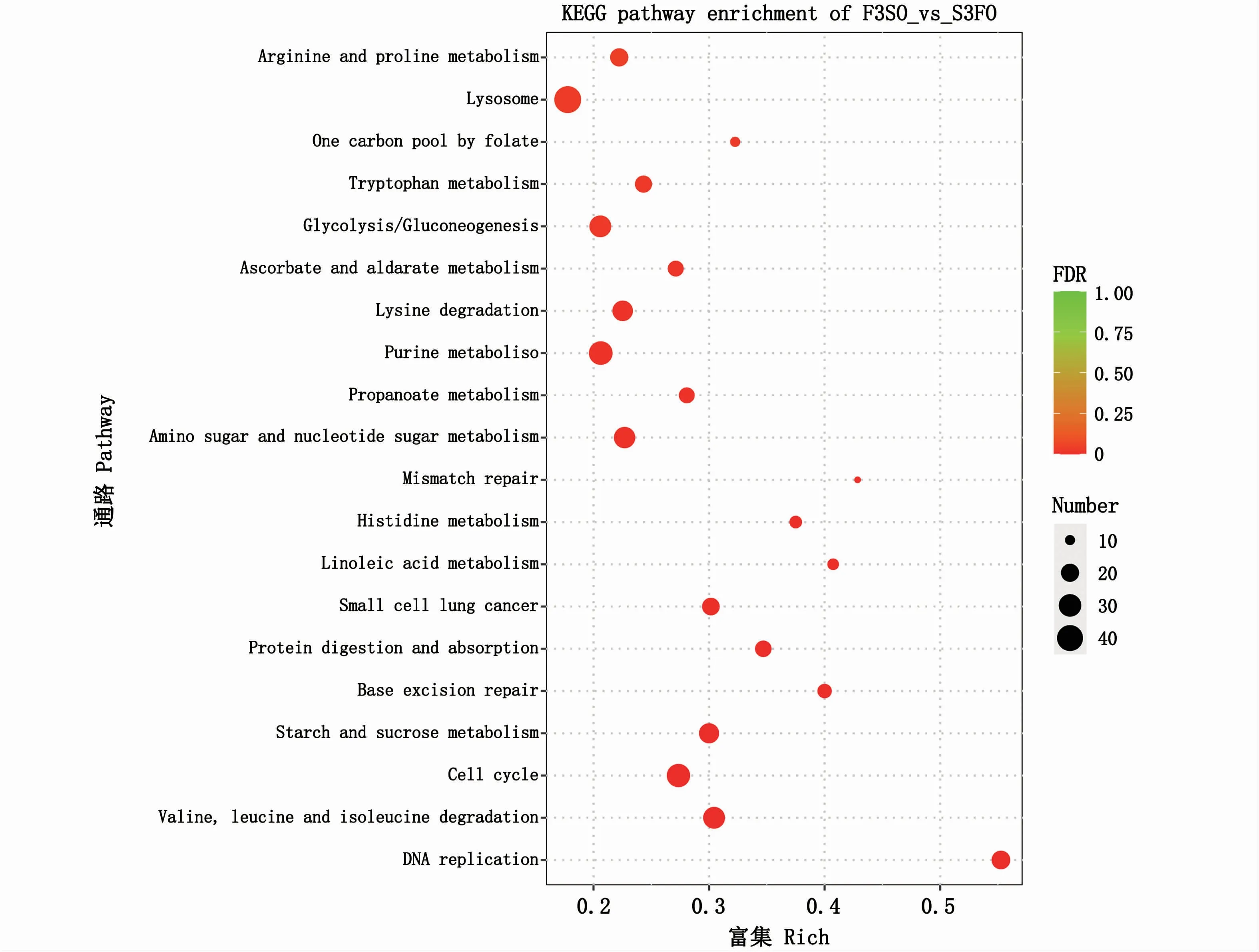
圖4 差異表達基因富集最顯著的前20條KEGG通路Fig.4 Top 20 significantly enriched KEGG pathways based on differentially expressed genes
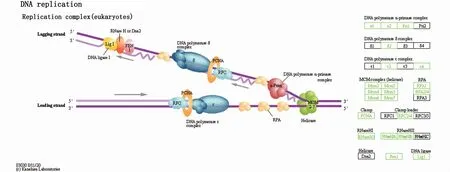
圖5 差異表達基因在DNA復(fù)制通路中的分布Fig.5 Distribution of differentially expressed genes in DNA replication pathway
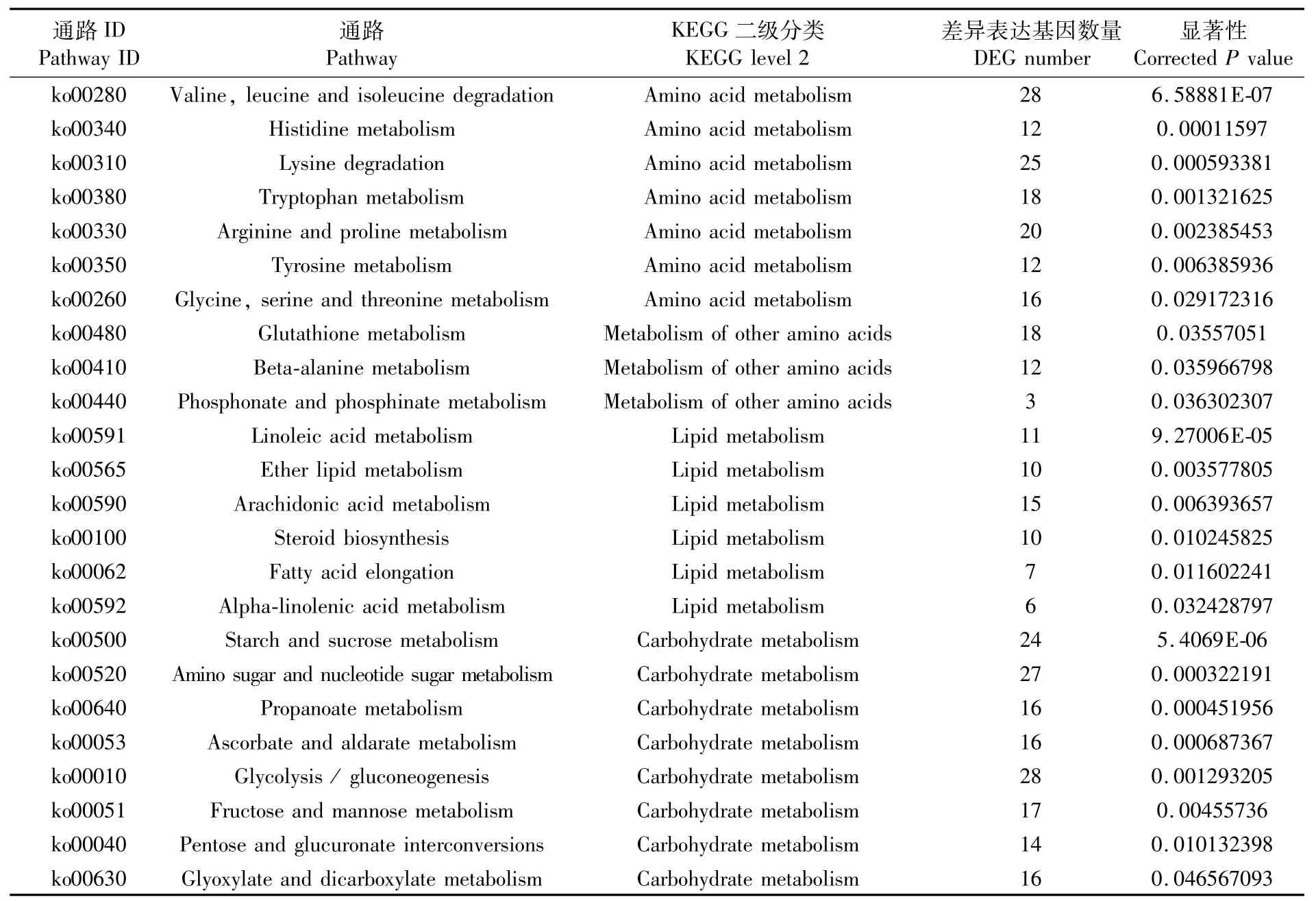
表5 與氨基酸代謝、脂質(zhì)代謝和碳水化合物代謝相關(guān)的顯著富集的KEGG通路Tab.5 Significantly enriched KEGG pathways related to am ino acid metabolism,lipid metabolism and carbohyd ratemetabolism
2.4 饑餓脅迫下消化系統(tǒng)、嗅覺和晝夜節(jié)律相關(guān)基因的篩選
基于KEGG通路結(jié)合以往的報道,篩選了部分與消化系統(tǒng)、嗅覺和晝夜節(jié)律相關(guān)的顯著差異表達基因(表6)。
2.5 差異表達基因qRT-PCR驗證
選取上調(diào)基因Met、MIH、E75和下調(diào)基因αamylase和Trypsin進行qRT-PCR驗證(圖6)。結(jié)果顯示,驗證結(jié)果與測序結(jié)果是一致的,表明轉(zhuǎn)錄組的測序結(jié)果和生物信息學(xué)分析是可靠的。
3 討論
擬穴青蟹作為一種重要的海水養(yǎng)殖蟹,大眼幼體期的營養(yǎng)狀態(tài)對其后期生長發(fā)育是十分重要的[17]。但目前關(guān)于饑餓是如何影響擬穴青蟹生長發(fā)育的分子研究未見報道,因而通過轉(zhuǎn)錄組測序挖掘相關(guān)的代謝通路是十分必要的。本研究測序獲得79 581個unigene,平均長度1 065.44 bp,以Nr數(shù)據(jù)庫注釋的unigene數(shù)最多(20 061個),但也僅占總數(shù)的25.21%,可見大部分unigene未得到有效注釋。
以往的研究表明,饑餓狀態(tài)下鋸緣青蟹、三疣梭子蟹和遠海梭子蟹(Portunuspelagicus)Ⅰ期溞狀幼體的生化組成發(fā)生了改變,主要體現(xiàn)為蛋白質(zhì)、脂質(zhì)的減少[14-16]。本研究中,多數(shù)氨基酸代謝和脂質(zhì)代謝相關(guān)通路同樣得到顯著性富集。不同的是,鋸緣青蟹[14]和三疣梭子蟹[15]Ⅰ期溞狀幼體在饑餓狀態(tài)下碳水化合物的含量基本保持不變,而本研究中有8條碳水化合物代謝相關(guān)通路得到顯著性富集。此外,鋸緣青蟹Ⅰ期溞狀幼體在饑餓狀態(tài)下蛋白酶、淀粉酶和酯酶的活性明顯下降[18]。本研究中擬穴青蟹大眼幼體的胰蛋白酶、淀粉酶和三酰甘油酯酶基因的表達水平是顯著下降的。可見,由于饑餓原因,青蟹大眼幼體下調(diào)了自身的消化酶分泌水平。
DNA復(fù)制是細胞最基本的生命活動之一,是生物體生存和繁殖的基礎(chǔ)。KEGG富集分析顯示,DNA復(fù)制通路富集是最顯著的,且富集的21個差異表達基因全部下調(diào)。此外,堿基切除修復(fù)、錯配修復(fù)、非同源末端連接和細胞周期途徑同樣顯著富集。以上這些結(jié)果表明,饑餓干擾了擬穴青蟹大眼幼體的DNA復(fù)制和修復(fù),進而影響了細胞周期。此外,細胞凋亡代謝通路是顯著富集的。以往的研究有過相似的結(jié)果,例如饑餓脅迫誘導(dǎo)了中華絨螯蟹肝胰腺的細胞凋亡[3]。由此可知,饑餓一定程度干擾了擬穴青蟹大眼幼體細胞的分裂和凋亡。
睡眠和覓食是高度保守、相互關(guān)聯(lián)的行為,對機體生存至關(guān)重要。已有研究發(fā)現(xiàn)饑餓抑制了果蠅(Drosophilamelanogaster)的睡眠,而饑餓后再次攝食果蠅的睡眠時間增加[19]。但對于饑餓調(diào)控睡眠的機理目前依然知之甚少。過去的研究揭示,果蠅大腦中的絲氨酸代謝調(diào)控饑餓誘導(dǎo)的睡眠抑制[20]。而最近的研究顯示,果蠅胰島素樣肽Dilp2是饑餓引起果蠅睡眠深度增加的關(guān)鍵神經(jīng)肽,但其不參與睡眠時間的調(diào)控[21]。根據(jù)先前的報道,果蠅Dilp或者其受體DInR的突變都將抑制睡眠,而饑餓降低了Dilp2的表達水平[22]。本文測序分析結(jié)果顯示,饑餓狀態(tài)下擬穴青蟹晝夜節(jié)律基因per和tim表達水平顯著變化,這表明饑餓干擾了擬穴青蟹的睡眠。與此同時,本文絲氨酸代謝通路顯著富集,部分胰島素樣肽受體相關(guān)基因差異表達,與果蠅中的發(fā)現(xiàn)類似。但對于饑餓是否直接通過上述途徑干擾擬穴青蟹睡眠則需要更多的實驗加以驗證。此外,以往研究中還發(fā)現(xiàn)饑餓調(diào)節(jié)果蠅的嗅覺行為[23],而本文的轉(zhuǎn)錄組測序同樣發(fā)現(xiàn)部分嗅覺相關(guān)基因在饑餓狀態(tài)下是顯著差異表達的,但饑餓與擬穴青蟹嗅覺行為的關(guān)系仍需進一步的驗證。
qRT-PCR結(jié)果表明,測序結(jié)果與驗證結(jié)果是一致的,因而轉(zhuǎn)錄組基于測序結(jié)果進行的生信分析是可信的。本研究通過轉(zhuǎn)錄組分析,獲得了擬穴青蟹大眼幼體的轉(zhuǎn)錄組序列和基因注釋結(jié)果,篩選了與饑餓脅迫相關(guān)的一系列代謝通路,例如氨基酸代謝、脂質(zhì)代謝、碳水化合物代謝和細胞生長和死亡等,分析了饑餓狀態(tài)下部分與消化系統(tǒng)、嗅覺和晝夜節(jié)律相關(guān)的基因表達水平的差異。研究表明,饑餓脅迫干擾了擬穴青蟹大眼幼體的細胞分裂和凋亡,影響了體內(nèi)的氨基酸代謝、脂質(zhì)代謝和碳水化合物代謝。本研究的相關(guān)發(fā)現(xiàn)有助于推進擬穴青蟹饑餓脅迫的分子機制研究。
猜你喜歡
音樂探索(2022年2期)2022-05-30 21:01:37
民用飛機設(shè)計與研究(2020年4期)2021-01-21 09:15:02
小天使·一年級語數(shù)英綜合(2019年8期)2019-08-27 02:23:00
中國特種設(shè)備安全(2018年11期)2019-01-08 02:08:32
電子制作(2018年18期)2018-11-14 01:48:24
小學(xué)科學(xué)(學(xué)生版)(2018年7期)2018-08-13 09:33:04
山東工業(yè)技術(shù)(2016年15期)2016-12-01 05:31:22
鄭州大學(xué)學(xué)報(醫(yī)學(xué)版)(2015年2期)2015-02-27 14:50:46
中國中醫(yī)藥現(xiàn)代遠程教育(2014年11期)2014-08-08 13:23:44
山東女子學(xué)院學(xué)報(2014年6期)2014-03-01 02:24:55
