長江口浮游動物群落結構時空變化特征研究
2023-09-14 11:46:02楊陽陽楊杰青王云龍
海洋漁業 2023年4期
楊陽陽,楊杰青,王云龍,晁 敏
(1.上海海洋大學海洋生態與環境學院,上海 201306;2.中國水產科學研究院東海水產研究所,農業農村部東海漁業資源開發利用重點實驗室,上海 200090)
長江口是中國主要的河口之一,受長江沖淡水、沿岸流系、臺灣暖流等多重因素的影響[1-2],水文環境復雜多變,且受人類活動影響顯著[3],是典型的河口生態區[4-5]。長江口擁有優越的自然條件,其豐富的營養物質以及較高的初級生產力為浮游動物的生長提供了優良條件。浮游動物在海洋生態系統中有著至關重要的作用,它是生態系統完整性的生物指標[6],其種類組成、生物量和次級生產力的變化都會對生態系統的結構和功能產生影響[7],并可對水環境的變化產生響應[8]。
有關長江口浮游動物群落結構變化的研究已有不少報道[9-15],這些研究分析了浮游動物群落結構的變化規律,表明鹽度和溫度[10,12-14]是影響浮游動物群落結構的主要因子,高倩等[13]、鄭金秀等[14]都研究表明,溫度是影響群落結構季節變化的主要因素,鹽度是影響群落結構區域變化的決定因素;王曉東和李超倫[15]研究表明,浮游動物不同聚類組群的地理分布與海域的鹽度梯度分布相對應;徐兆禮[12]研究表明,群落結構受到不同性質水團以及長江沖淡水的交互影響。上述研究大多針對某個季節或某幾個航次,在較長時間序列上的群落結構研究較少。國外對河口浮游動物的研究也很多[16-21],如BURDIS和HIRSCH[16]使用聚類、排序等方法研究密西西比河浮游動物群落結構的時空變化特征,通過ANOSIM程序分析群落結構間是否存在差異性,采用SIMPER程序分析導致群落差異的貢獻物種;STEFANIDIS和PAPASTERGIADOU[17]采用Kruskal-Wallis分析方法檢驗希臘湖泊中浮游動物豐度的差異性。本研究基于2006—2008年、2016—2018年兩個時間段每年春季和夏季的調查資料,探討浮游動物群落結構的時空變化,確定較長時間序列基礎上的浮游動物生態地理劃分特點,明確浮游動物的季節及年間變化,以期為更好地了解長江口及其鄰近海域的生態變化提供科學參考。
1 材料與方法
1.1 資料來源及采樣方法
數據來自農業農村部東海漁業生態環境監測中心2006—2008年和2016—2018年每年春季(5月)和夏季(8月)在長江口海域的調查資料。相比2006—2008年,2016—2018年站位設置有部分調整,因此只采用共有的11個站位資料,其中1~4站位位于口門區及最大渾濁帶區域,5~11站位位于杭州灣(圖1)。
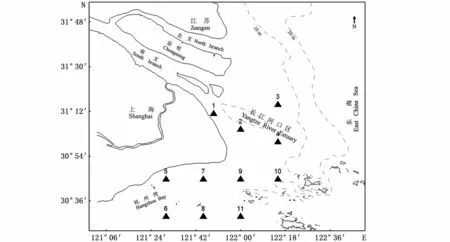
圖1 長江口調查海域及站位分布Fig.1 Investigation sea area and sam pling stations in the Yangtze Estuary
浮游動物標本采集使用淺水I型浮游生物網自底至表垂直拖拽,采集及標本處理均按照《海洋調查規范》(GB/T 12763.6—2007)[18]進行,經5%福爾馬林溶液固定、保存,實驗室內進行分類、鑒定和統計。本文浮游動物數據未統計水母類和海樽類,浮游動物豐度單位為:個·m-3。
1.2 統計分析
浮游動物優勢種判別采用優勢度公式:
式中,ni為第i種浮游動物豐度;N為浮游動物總豐度;fi為第i種浮游動物在各站位中的出現頻率。優勢度Y≥0.02的種為本文優勢種。
以2006—2008年春、夏季和2016—2018年春、夏季各站每種浮游動物豐度均值數據為基礎,用SPSS 26進行Kruskal-Wallis檢驗,分析種類組成和豐度差異[17];用PRIMER 5.0進行群落聚類、排序(nMDS分析)、ANOSIM和SIMPER分析,其中聚類和nMDS分析用于研究具有時空異質性的站位組[24],ANOSIM程序確認異質性[25],SIMPER程序確定對分組有主要貢獻的種類[26]。用Surfer 14繪制站位圖及浮游動物豐度空間分布圖。
2 結果與分析
2.1 長江口環境概況
2006—2008年長江口平均徑流量為7 624.6億m3,2016—2018年平均徑流量為9 285.3億m3,2016—2018年流量明顯增加。鹽度春季高、夏季低,2016—2018年低于2006—2008年,且空間分布格局呈近岸低、外海高的變化趨勢。水溫春季低、夏季高,口門區及沿岸站位高于外海站位,夏季水溫的空間分布格局與長江沖淡水的擴散方向相一致。
2.2 種類組成
12次航次共鑒定出浮游動物109種,分別隸屬于5門14大類,包括枝角類、橈足類、端足類、磷蝦類、十足類、等足類、糠蝦類、漣蟲類、介形類、翼足類、異足類、多毛類、毛顎類和被囊類,橈足類的種類數占絕對優勢(表1)。其中,2006—2008年春、夏季6個航次共采集浮游動物13大類群73種,2016—2018年春、夏季6個航次共采集浮游動物10大類群63種。春季浮游動物種類數70種,夏季83種,2016—2018年春、夏季物種數相比2006—2008年均有所減少(圖2)。Kruskal-Wallis檢驗結果表明,同一調查階段不同季節間(春季和夏季)、同一季節不同調查階段間(2006—2008年和2016—2018年)的類群物種數變化均不顯著(P>0.05),但同一季節不同調查階段間的站位物種數有顯著變化(P<0.05)。
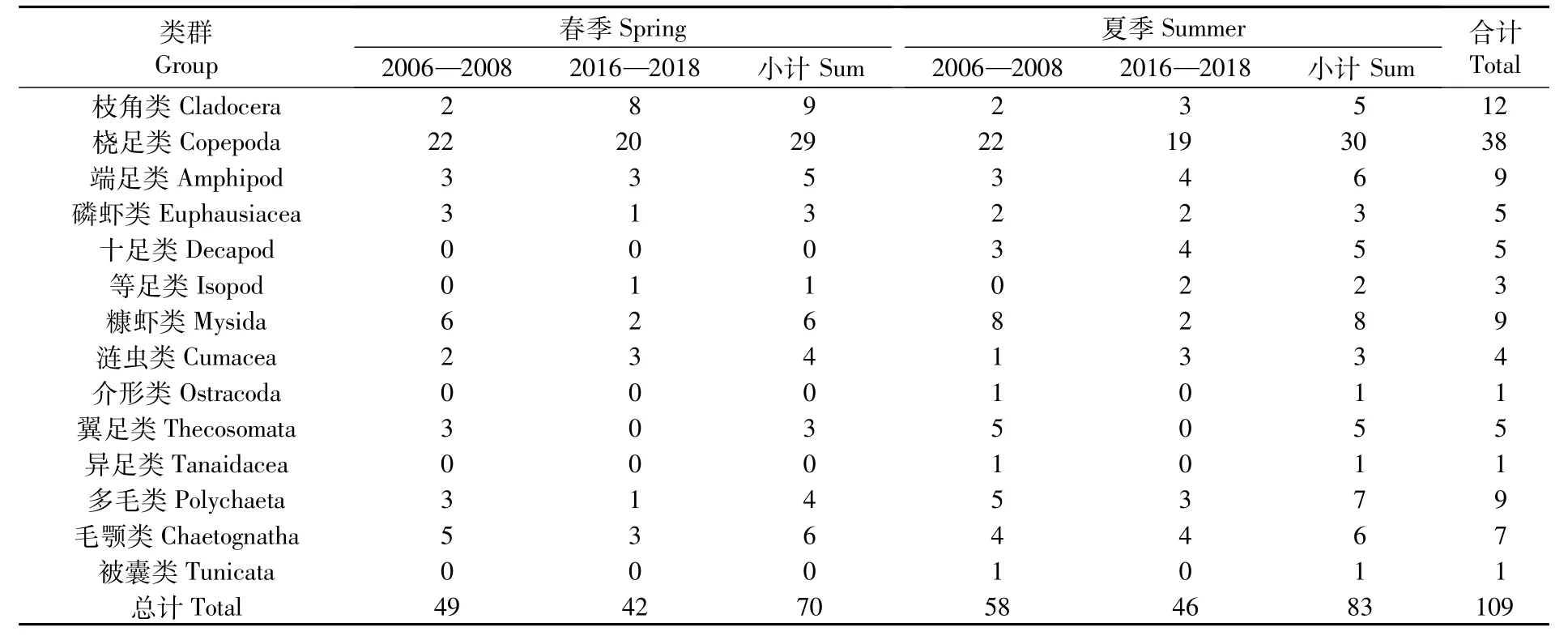
表1 長江河口浮游動物不同類群物種數Tab.1 Species of different groups of zooplankton in the Yangtze Estuary
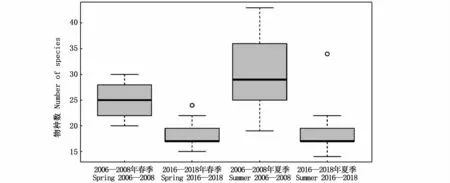
圖2 長江河口各調查站物種數Fig.2 Number of species at each survey station in the Yangtze Estuary
2.3 浮游動物優勢種
優勢度Y≥0.02的優勢種(表2)共計13種,分屬3個類群,包括橈足類10種,糠蝦類2種,毛顎類1種。
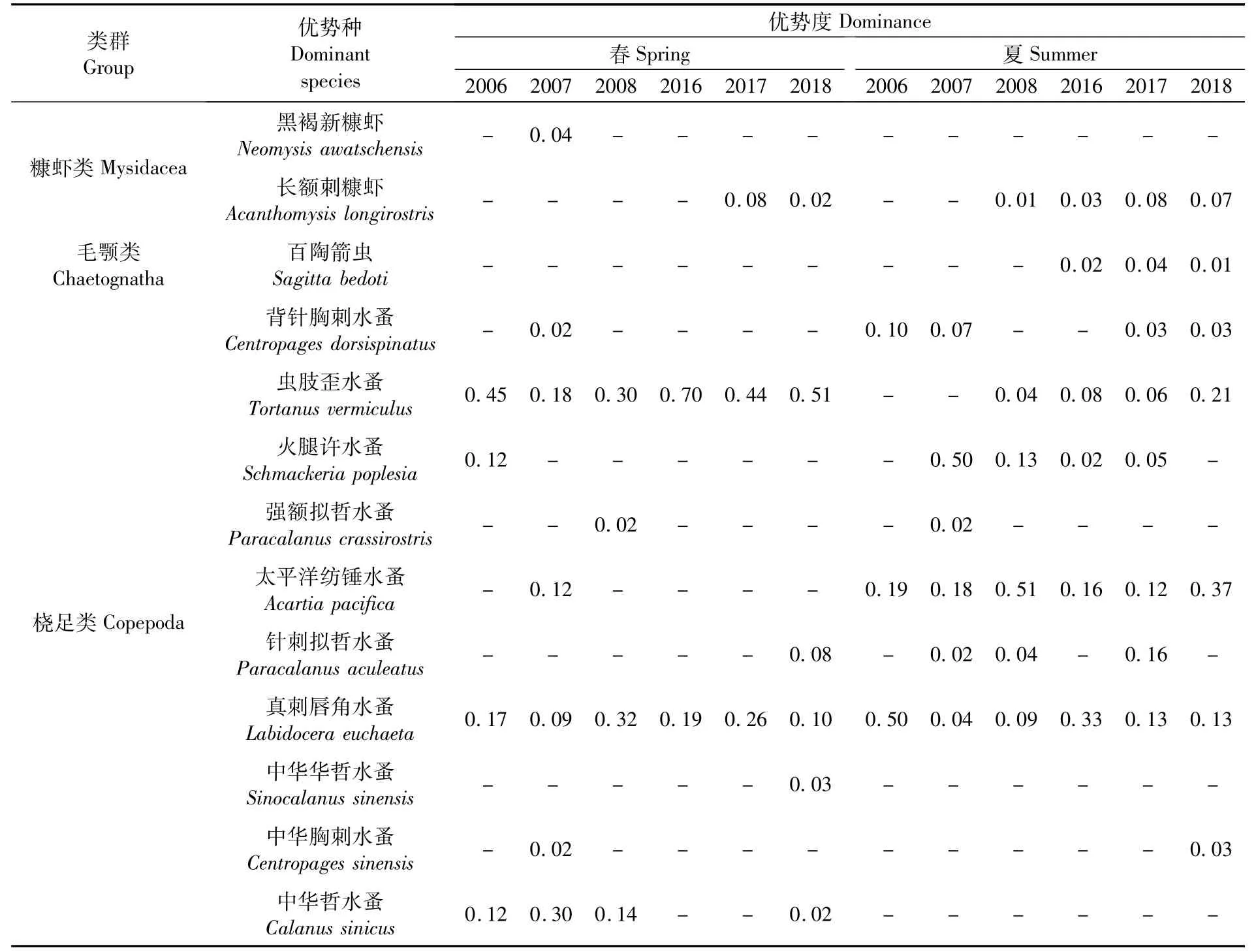
表2 各航次長江口浮游動物優勢種Tab.2 Dom inant species of zoop lankton in the Yangtze Estuary in each voyage
在12次調查中,真刺唇角水蚤(Labidocera euchaeta)均為優勢種,蟲肢歪水蚤(Tortanus vermiculus)有10次為優勢種,太平洋紡錘水蚤(Acartiapacifica)有7次為優勢種。
春季主要優勢種群比較穩定,蟲肢歪水蚤、真刺唇角水蚤為春季主要優勢種,兩者在6個航次中均為優勢種,其中蟲肢歪水蚤有4次為第一優勢種、2次為第二優勢種,真刺唇角水蚤有1次為第一優勢種、4次為第二優勢種。夏季主要優勢種群亦比較穩定,主要優勢種為太平洋紡錘水蚤、真刺唇角水蚤和火腿許水蚤(Schmackeria poplesia)。太平洋紡錘水蚤和真刺唇角水蚤在6個航次中均為優勢種,其中,太平洋紡錘水蚤2次為第一優勢種、3次為第二優勢種,真刺唇角水蚤有2次為第一優勢種;火腿許水蚤有4次為優勢種,其中1次為第一優勢種。
2.4 浮游動物豐度時空分布特征
2006—2008年春季航次豐度均值為333.87個·m-3,夏季為562.75個·m-3;2016—2018年春季航次豐度均值為119.30個·m-3,夏季為208.40個·m-3。浮游動物豐度呈夏季高于春季及2016—2018年春、夏季豐度均低于2006—2008年同期的特點。
春季,兩個調查階段中浮游動物豐度均呈現沿岸略高、外海略低的特點(圖3-A,C),豐度高值區均分布在北導堤外側(3號站)及杭州灣灣口(7號站)附近海域。Kruskal-Wallis檢驗結果顯示,不同調查階段間的豐度具有顯著性差異(P<0.05);但在每一調查階段中,相鄰3個航次間的浮游動物豐度無顯著性差異(P>0.05)。
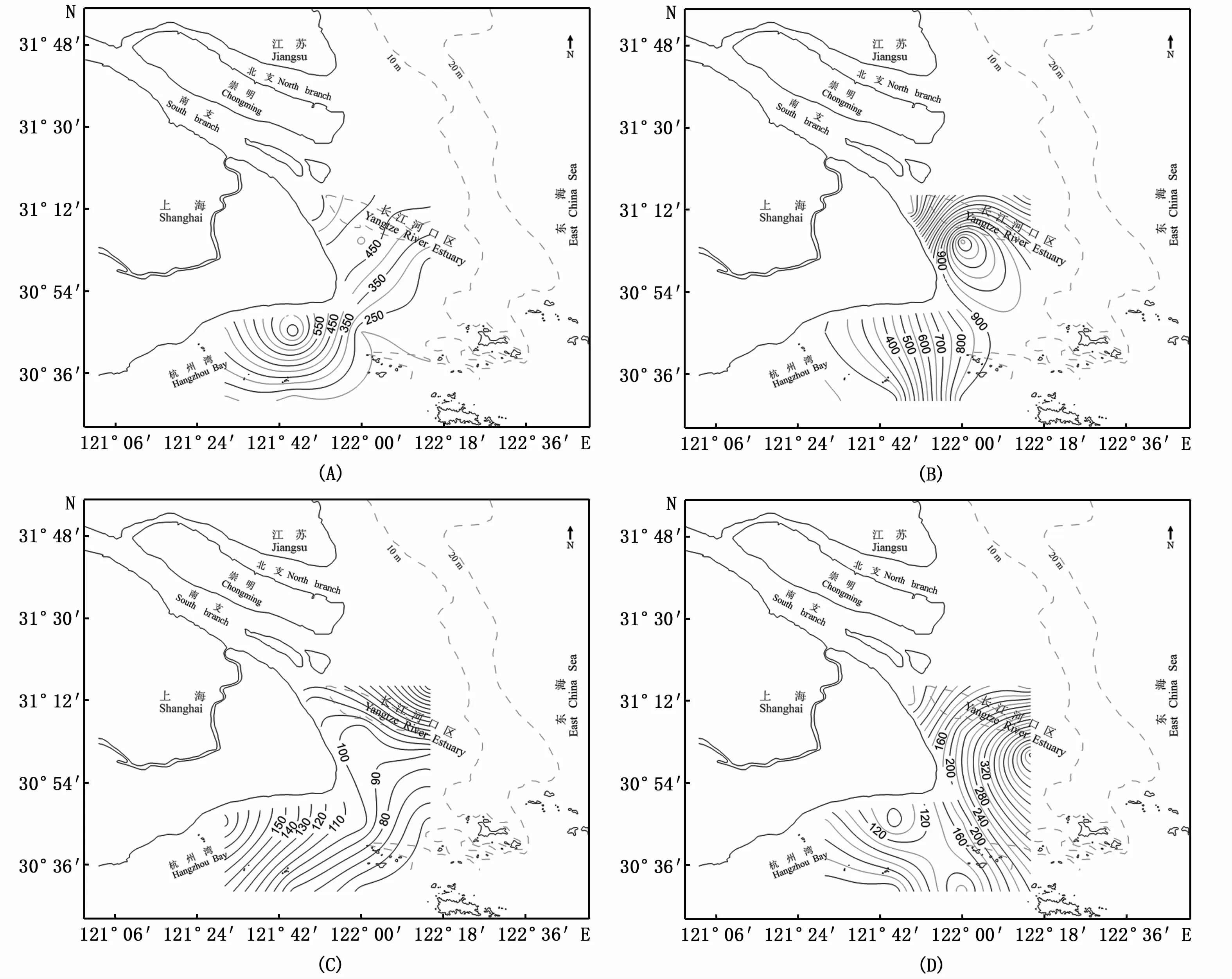
圖3 長江口浮游動物豐度的分布(單位:個·m-3)Fig.3 Abundance distribution of zooplankton in the Yangtze Estuary
夏季,與春季相比,浮游動物豐度高值區向外海移動(圖3-B,D),高值區多分布于崎嶇列島-雞骨礁連線鄰近海域。Kruskal-Wallis檢驗結果顯示,不同調查階段間的豐度具有顯著性差異(P<0.05),2016—2018年相鄰3個航次間的浮游動物豐度間有顯著性差異(P<0.05),但2006—2008年相鄰3個航次間的浮游動物豐度間無顯著性差異(P>0.05)。
2.5 浮游動物群落聚類及nMDS分析
分別對2006—2008年春、夏季和2016—2018年春、夏季浮游動物豐度數據進行聚類和nMDS分析,結果見圖4和圖5。nMDS分析的stress值均低于0.2,說明聚類結果有一定的可信度[27-28]。

圖4 長江口不同季節浮游動物群落聚類Fig.4 Clustering of zooplankton community in different seasons in the Yangtze Estuary

圖5 長江口不同季節浮游動物群聚排序Fig.5 O rdination p lots of zoop lankton community in different seasons in the Yangtze Estuary
2006—2008年春、夏季浮游動物都可劃分為3個類群:類群I僅包含3號站,為河口近海水域站位,主要受近海水團影響,屬近海群落;類群II為河口水域站位,其中春季為河口口門區1號站位所形成的浮游動物群落,夏季則為河口口門區1號和九段沙附近2號站位形成的浮游動物群落,該區為最大渾濁帶的核心區域,屬長江口口門水域群落;類群III(共有站位有4、5、6、7、8、9、10、11)主要為崎嶇列島-雞骨礁連線附近站位及杭州灣口各站,主要受枯季長江徑流與口外近海水團的交錯影響,屬咸淡水交錯水域群落。
2016—2018年春、夏季浮游動物可劃分為相同的2個類群:類群IV(站位有4、5、6、7、8、9、10、11)主要為崎嶇列島-雞骨礁連線附近站位及杭州灣口各站,主要受夏季長江口沖淡水與外海水團的交錯影響,屬咸淡水交錯水域群落;類群V(站位有1、2、3)主要為河口最大渾濁帶水域,屬夏季長江沖淡水控制區群落。
2.6 不同時空浮游動物群落相似性分析
聚類和nMDS分析結果表明,類群I、II和V在地理位置上接近,均為近河口浮游動物群落,可概括為同一組群,命名為近河口組群。類群III和IV位于河口口門南側的杭州灣、崎嶇列島-雞骨礁連線水域,受咸淡水控制,可概括為同一組群,命名為河口南側組群。
ANOSIM相似性分析檢驗結果如表3所示,同一調查階段同一季節的不同聚類組群間均具有顯著性差異(P<0.05)。春季,蟲肢歪水蚤、真刺唇角水蚤等是導致浮游動物群落聚群組內相似和組間分異的關鍵物種。夏季,太平洋紡錘水蚤、火腿許水蚤和真刺唇角水蚤是導致浮游動物群落聚群組內相似和組間分異的關鍵物種。
ANOSIM相似性分析結果如表4和表5所示,同一組群在同一季節不同調查階段間均具有顯著性差異(P<0.05),在同一調查階段不同季節間亦具有顯著性差異(P<0.05)。春季,導致不同調查階段間浮游動物群落分異的關鍵物種有蟲肢歪水蚤、真刺唇角水蚤和中華哲水蚤,但其豐度在2016—2018年均顯著降低;夏季,導致不同調查階段間浮游動物群落分異的關鍵物種有太平洋紡錘水蚤、火腿許水蚤和真刺唇角水蚤,其豐度在2016—2018年亦顯著降低。季節差異的相似性分析結果表明,在兩個調查階段中,蟲肢歪水蚤、太平洋紡錘水蚤及真刺唇角水蚤的豐度變化均是導致浮游動物豐度季節差異的最主要原因。
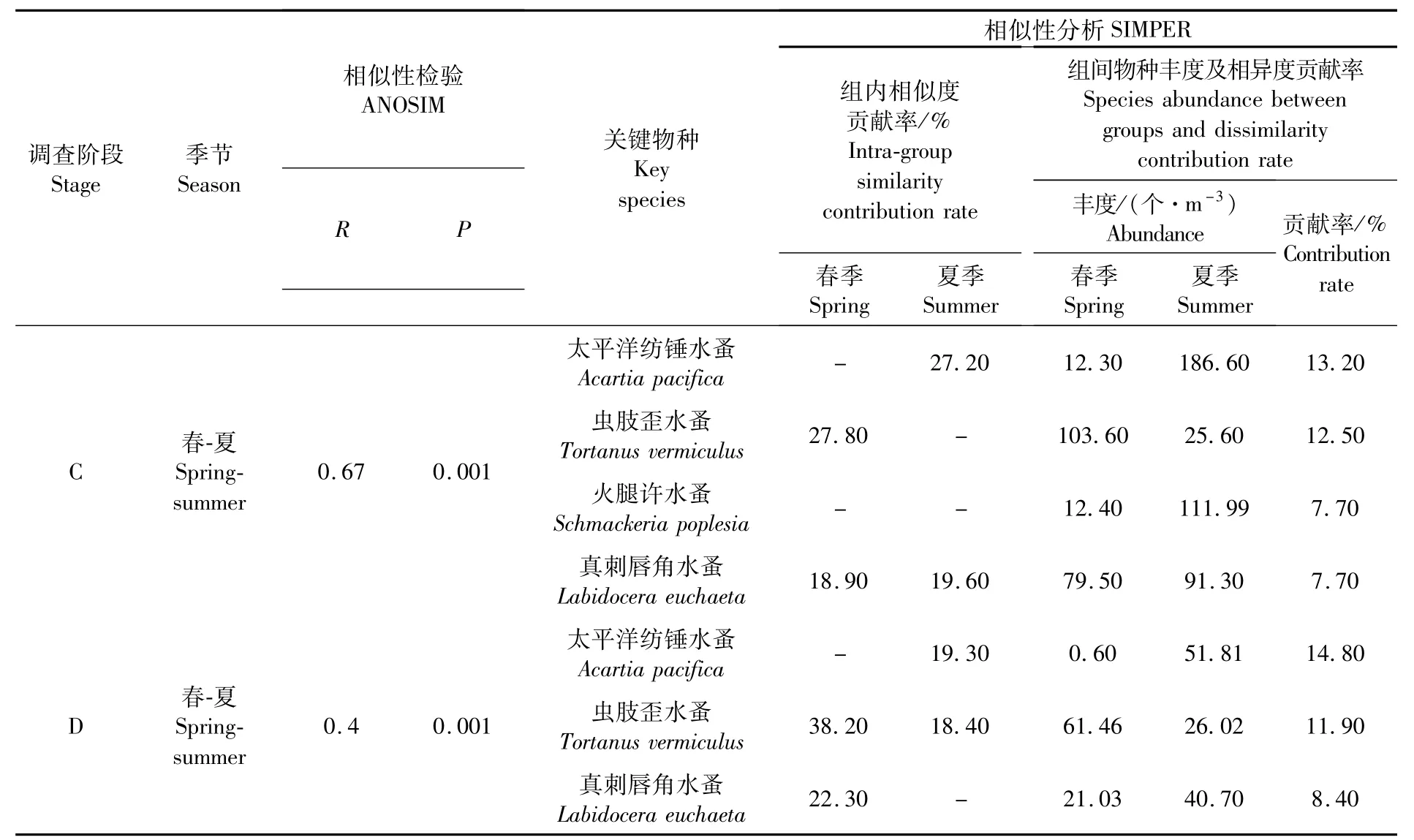
表5 不同季節相似性分析檢驗和相似性分析Tab.5 Sim ilarity analysis test and sim ilarity analysis in different seasons
3 討論
3.1 浮游動物種類組成以及優勢種的時空變化
分析比較不同調查階段浮游動物的種類組成,可以發現與2006—2008年相比,2016—2018年浮游動物種類數呈減少趨勢,這與楊穎等[29]在長江口的數據趨勢相似,橈足類種類數比2006—2008年減少。浮游動物種類的變動與河口水沙條件的變化有很大關系[30],從大通站徑流數據看,2016—2018年3年平均徑流量比2006—2008年增加了21.8%,徑流量的增加可能導致了強卷螺 (Agadina syimpsoni)、長軸螺 (Peraclis reticulata)等暖水性浮游翼足類在調查區的消失。
在兩個調查階段均觀測到物種數量具有季節差異,夏季物種數多于春季,主要是由于夏季處于豐水期,長江沖淡水以及外海暖流的勢力較春季均有所增強,不但為整個調查水域引入了更多的外海暖水種,而且不同水團間的劇烈交匯,形成了更為復雜的水域環境,可容納更多種的生物類型生存,因此浮游動物類群變化范圍被進一步擴大[10,31]。
在本研究鑒定的優勢種中,真刺唇角水蚤、蟲肢歪水蚤和太平洋紡錘水蚤廣泛分布于調查海域各站,其中蟲肢歪水蚤豐度高值通常出現在春季,真刺唇角水蚤在春夏季均有較高的豐度,太平洋紡錘水蚤在夏季有較高豐度,這些物種均為典型河口、沿岸低鹽物種[23],可適應長江口劇烈變化的溫鹽環境。值得注意的是中華哲水蚤,其屬近海廣溫廣鹽種,在2006—2008年春季航次的近河口海域大量出現(表2,表3),該海域受長江徑流和海洋潮汐的影響,該種豐度與溫鹽呈顯著正相關關系[10];當2016—2018年長江徑流量相對增加時,該海域受低鹽水控制,中華哲水蚤不再成為優勢物種。
上述分析結果表明,真刺唇角水蚤、蟲肢歪水蚤和太平洋紡錘水蚤等河口沿岸低鹽種為長江河口區主要優勢物種,其豐度會受到水溫、鹽度等環境因子的綜合影響,某些物種如中華哲水蚤的分布會受到長江徑流和海洋潮汐相互作用強度以及適溫適鹽性的影響,當長江入海流量減少時,近海廣鹽物種有可能進入河口區成為優勢物種。
3.2 浮游動物豐度的時空變化特征
春季,蟲肢歪水蚤和真刺唇角水蚤等優勢種在長江口沿岸海域大量聚集,導致其在該區域的豐度值偏高,使浮游動物豐度均值呈現沿岸高外海略低的特點,這與之前的研究結果相似[31-32]。夏季,受長江沖淡水勢力增大影響,豐度高值區相比春季向外海移動,集中在咸淡水交錯水域。夏季浮游動物群落構成受到高溫高鹽的臺灣暖流與長江沖淡水的影響,沿岸流帶來的太平洋紡錘水蚤、火腿許水蚤等適應河口低鹽生境的優勢種大量繁殖。
浮游動物豐度均值呈現出夏季高春季低的現象,主要是由于夏季水溫較高,咸淡水交錯區營養物質豐富,浮游植物數量充足,有利于浮游動物的生長繁殖,導致夏季相比春季豐度增加[33]。浮游動物豐度在2016—2018年春季和夏季均比2006—2008年同季節顯著下降,從相似性分析結果看,優勢物種豐度的降低是差異的主要原因,推測與長江口近年來的生態環境變化密切相關[34],2016—2018年長江徑流量的增加使得調查區域的鹽度相比2006—2008年時低,限制了真刺唇角水蚤、蟲肢歪水蚤、太平洋紡錘水蚤、中華哲水蚤等在本區域的大量繁殖。由此可見,水團的季節波動、外海高溫高鹽水和長江沖淡水交匯的位置以及潮汐的漲落等因素均有可能影響長江口浮游動物豐度空間分布[35]。
3.3 浮游動物群落結構的時空變化特征
基于較長時間序列的浮游動物豐度數據、群落聚類和nMDS分析,可以發現,不論2006—2008年還是2016—2018年,春、夏季的浮游動物群落空間分布趨勢基本相同,聚類類群位置相對穩定。浮游動物類群I、II和V屬于近河口區域群落,類群III和IV在長江沖淡水和咸水相互作用較強的地區,受咸淡水控制,屬于河口南側組群。本調查海域鹽度在春、夏季均呈現長江口口門和近岸向外海擴散增高的趨勢,因此兩個區域水鹽條件有一定差別,鹽度分布是決定長江口浮游動物群落劃分及其空間分布的主要決定性因子[36]。
浮游動物群落相似性分析結果表明,在長江口及鄰近咸淡水交錯區,春季浮游動物群落可歸納為蟲肢歪水蚤-真刺唇角水蚤群落,夏季可歸納為太平洋紡錘水蚤-真刺唇角水蚤群落,在長江徑流影響力減弱的年份,中華哲水蚤等廣鹽種有可能隨高鹽水進入調查區域成為優勢物種。
猜你喜歡
課堂內外·初中版(科學少年)(2025年1期)2025-02-28 00:00:00
課堂內外·初中版(科學少年)(2025年2期)2025-02-28 00:00:00
我愛學·笑話與口才(2025年3期)2025-02-24 00:00:00
英語世界(2023年10期)2023-11-17 09:18:18
趣味(語文)(2020年3期)2020-07-27 01:42:46
科學大眾(中學)(2019年3期)2019-05-17 10:04:30
汽車觀察(2018年10期)2018-11-06 07:05:26
作文與考試·初中版(2017年12期)2017-04-19 20:26:27
中國衛生(2015年2期)2015-11-12 13:13:54
少兒科學周刊·少年版(2015年1期)2015-07-07 17:15:12
