高寒草甸植物群落功能多樣性對不同生長期干旱的響應機制
2023-09-11 07:26:04劉慧慧胡國錚干珠扎布冀國旭王子欣高清竹王慶鎖旦久羅布
生態學報 2023年16期
劉慧慧, 胡國錚, 干珠扎布, 冀國旭, 王子欣, 高清竹, 王慶鎖,嚴 俊, 旦久羅布
1 中國農業科學院農業環境與可持續發展研究所, 北京 100081 2 農業農村部農業農村生態環境重點實驗室,北京 100081 3 國家農業環境數據中心,北京 100081 4 西藏那曲市草原站,那曲 852000
在干旱脅迫下植物形態結構、生理功能及生物化學過程受到抑制作用,如葉面積和葉厚度、葉片氣孔導度、葉片凈光合速率[1]受限。在植物個體水平上表達為植物功能性狀的差異[2],植物通過采取不同的資源分配策略,以達到最高的生存能力[3],進而對生態系統關鍵過程產生不同影響。植物物種間差異體現在物種對不同生境的適應,即植物的功能性狀[4]。與關鍵生態過程相關的性狀被稱為植物生活史的核心,包括植物個體大小、種子大小和葉組織結構(葉面積、干物質含量等),并對資源利用、植物再生和擴散密切相關并對干擾有顯著響應[5—6]。功能多樣性作為生物多樣性的一個重要方面,是指可以影響生態系統功能的物種性狀值和分布范圍[7]。基于植物功能性狀和群落功能多樣性的研究,能更好地探知不同生境中植物面臨的限制和機遇[8],預測植物群落組成[9—10],衡量功能多樣性對生態系統功能影響的作用[6,8,11—12]。越來越多的研究表明,生態系統的功能如群落生產力、物質循環、能量流動、對干擾或入侵的恢復力[13—14]更依賴于群落功能多樣性[2,15—18]。全面深入地了解植物功能多樣性與生態系統過程或功能之間的關系,有助于更好地應對全球氣候變化和人類活動干擾下生物多樣性喪失的生態學后果。
被稱為“第三極”的青藏高原作為地球上最高的生物地理單元,具有獨特的水熱條件和復雜特殊的地理環境,氣候敏感并且生態環境脆弱[19—21],極易受到氣候變化和人類活動的干擾。在全球變化背景下,降水格局發生季節性變化,青藏高原出現極端干旱事件的頻率增加、強度加劇[22—23]。在草地生態系統中,干旱通常導致功能多樣性降低[24],不同植物的適應干旱的策略不同,優勢種多表現出忍耐干旱的策略,而稀有種多表現出躲避干旱的策略[24]。然而降水季節性的改變對植物功能多樣性影響的研究較少,青藏高原高寒草甸植物功能多樣性對不同生長季干旱的響應機制尚且不明。
通過模擬不同時期干旱事件,觀測藏北高寒草甸群落優勢種的蓋度、選取并測定植物功能性狀,基于功能性狀和物種蓋度計算群落功能多樣性指數,并分析群落功能性狀和功能多樣性對不同時期干旱的響應情況,以闡明不同生長時期干旱條件下高寒草甸植物群落功能多樣性對干旱脅迫的響應機制。為預測未來季節性干旱事件對青藏高原高寒草甸植物群落功能性狀、群落特征和功能多樣性的影響提供科學依據。
1 材料與方法
1.1 研究區概況
研究區地處念青唐古拉山以北的高寒草甸,位于西藏那曲市“國家農業環境那曲野外科學試驗站”內,地理坐標為92°02′E,31°44′N(圖1),平均海拔4500 m以上。根據國家氣象科學數據中心[25],試驗區近十年平均溫0.3℃,生長季(5—9月)月平均溫度高于0℃。年均降水量為450—500 mm,90%以上集中在生長季,平均土壤體積含水量約為22.86%,土壤質地為高山草甸土。研究區草地類型為寒冷濕潤凍原、高山草甸類[26],主要以高山嵩草(Kobresiapygmaea(C. B. Clarke) C. B. Clarke)為建群種,常見種有藏豆(Hedysarumtibeticum(Bentham) B. H. Choi &H. Ohashi)、釘柱委陵菜(PotentillasaundersianaRoyle)、多裂委陵菜(PotentillamultifidaL.)、二裂委陵菜(PotentillabifurcaL.)、弱小火絨草(Leontopodiumpusillum(Beauv.) Hand.-Mazz.)等,植物返青期多在5月,植物枯黃期多在8—9月。

圖1 研究區域圖Fig.1 Experimental plot●為試驗站地理位置,試驗樣地(右上),試驗樣地多年月平均降水量和氣溫情況(右下)。
1.2 試驗設計
本試驗自2016年起,通過設置截雨棚裝置模擬高寒草甸在不同生長階段的干旱事件[27],截雨棚為透光率90%的PC板,地面覆蓋范圍2 m×2 m的可移動裝置(圖1)。設置生長季前期(5—6月)干旱(Early growing season drought,ED)、生長季中期(7—8月)干旱(Middle growing season drought,MD)、非生長季(9月—次年4月)干旱(Non-growing season drought,ND)和對照未截雨(Control,CK)4個處理,4次重復,共16個子小區。每個處理小區邊界土壤中垂直埋入地下深度為30 cm的隔水板,并在處理小區間設置2 m緩沖區。每個處理小區分成4個0.5 m×0.5 m的子小區,其一作為觀測區,其三為破壞性采樣區,并在小區中埋設有EM50自動記錄器(Decagon公司,美國)監測土壤溫濕度,干旱處理小區的土壤體積含水量在截雨期間顯著降低[28]。
1.3 功能性狀的測定
于2021年8月中旬生長季高峰期進行群落調查和樣品采樣,根據“質量比假說”選取優勢種作為功能性狀的觀測對象[29—30],共選取高山嵩草、藏豆、釘柱委陵菜、多裂委陵菜、二裂委陵菜及弱小火絨草6個優勢物種,各小區中優勢種的分蓋度總和平均占小區群落總蓋度86.47%,最高達到95.11%。在各試驗小區的采樣子小區中根據“五點取樣法”選取5—10株個體,沿葉柄基部剪下最新生長且完全展葉的10—20個葉片,由于高山嵩草葉片狹小,采樣時選取20個葉片,其余5種植物采取10個葉片。取樣后即刻裝入密封袋中,保持葉片含水量。為研究不同時期干旱下高寒草甸植物的功能性狀的差異,參考LEDA Traitbase數據庫(www.leda-traitbase.org.)[2,8]對植物功能性狀的描述和選定標準,測定了8項葉片功能性狀:葉面積(LA:mm2)、比葉面積(葉面積與葉干重之比,SLA:mm2/mg)、葉干物質含量(葉干重與葉鮮重之比,LDMC: mg/mg)、葉碳含量(LCC:mg/mg)、葉氮含量(LNC:mg/mg)、葉磷含量(LPC:mg/mg)、葉碳氮含量比(C∶N:mg/mg)以及葉氮磷含量比(N∶P:mg/mg)。稱取葉片鮮重,置于60℃烘箱中烘干48 h至恒重,稱得干重,測量精確到0.01 g。葉面積通過對葉片掃描后,采用ImageJ(https://imagej.nih.gov/ij)軟件測量。LCC和LNC采用有機元素分析儀(Flash EA1112,Thermo Scientific,USA)測定,LPC采用釩鉬黃吸光光度法測定。
以優勢種的相對蓋度為權重,計算了每個功能性狀的群落加權平均值(CWM),即LACWM、SLACWM、LDMCCWM、LCCCWM、LNCCWM、LPCCWM、C∶NCWM和N∶PCWM,以表征群落的功能性狀。計算公式[31]:
式中,Pi為第i種蓋度占總蓋度的比例,n為物種數目,Xi是物種的特征值。
1.4 群落功能多樣性
群落功能多樣性由功能均勻度、功能分異度和功能分散度等指數描述,每個指數描述群落特征分布的不同方面[32],且對干旱的敏感程度及對生態過程影響存在差異。采用R-4.0.5軟件的FD程序包中dbFD函數[33]計算了功能均勻度指數(FEve)、功能分異度(FDiv)、功能分散度(FDis)和Rao二次熵(RaoQ)指數[34—35],各指標計算公式(表1)。數據均通過Bartlett方差齊性檢驗符合方差齊性,同時計算各功能多樣性指數的變異度。

表1 功能多樣性指數各指標及其計算公式和生態學涵義
并根據以下公式計算各功能多樣性指數的變異度:
ΔFDdp=FDdp-FDCK
式中,FD(functional diversity)代表功能多樣性指數,包括FEve,FDiv,FDis和RaoQ指數,dp(drought periods)為干旱時期,包括ED、MD和ND。ΔFD(即ΔFEve、ΔFDiv、ΔFDis和ΔRaoQ)表示不同干旱時期下功能多樣性指數與對照組CK相比的變異程度。
1.5 數據分析
采用R-4.0.5軟件進行統計學分析及繪圖,調用“FD”程序包中的dbFD函數計算功能多樣性指數。數據采用單因素方差分析(ANOVA)和Duncan多重比較分析群落總蓋度、優勢物種相對蓋度、優勢種的功能性狀、群落功能性狀在各處理間的差異,采用變異度分析群落功能多樣性對不同時期干旱處理的響應。并采用Pearson相關系數分析群落功能性狀和功能多樣性的相關性,探討群落功能性狀與功能多樣性對干旱響應間的關系。
2 結果與分析
2.1 優勢種蓋度對不同時期干旱的響應
優勢種的相對蓋度對不同時期干旱的響應存在差異。在ED處理下各優勢種相對蓋度均顯著降低(P<0.05),其中二裂委陵菜相對蓋度最低,為(2.3±1.0)cm,與CK相比降低了66.8%。MD處理下僅多裂委陵菜與二裂委陵菜的相對蓋度顯著降低(P<0.05),分別為(5.7±1.8)cm和(3.8±0.4)cm,比CK降低了 49.0%和44.5%。ND處理對各優勢種的相對蓋度均沒有顯著影響(圖2)。ED和MD處理導致植物群落總蓋度顯著降低(P<0.05)(圖3),分別為(59.2±1.7)cm和(67.0±9.8)cm。ED處理對優勢種相對蓋度的影最大,使其下降了37.6%,MD處理降低了優勢種相對蓋度的29.5%。

圖2 不同時期干旱處理下6個優勢種的相對蓋度Fig.2 Relative coverage of 6 dominant species under drought treatment in different timing不同小寫字母代表組間差異顯著,顯著性水平P<0.05;圖中數據為平均值±標準差;CK對照;ED生長前期干旱;MD生長中期干旱;ND非生長時期干旱
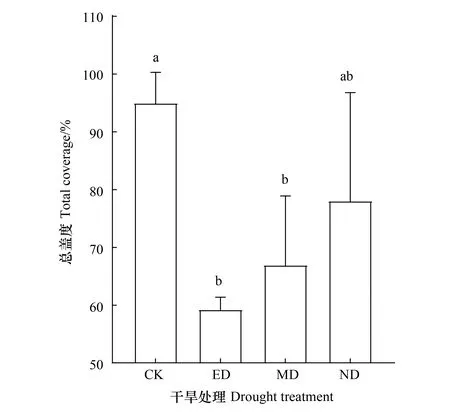
圖3 不同時期干旱處理下群落總蓋度 Fig.3 Total coverage under drought treatment in different timing 不同小寫字母代表組間差異顯著,顯著性水平P<0.05;圖中數據為平均值±標準差
2.2 優勢種功能性狀對不同時期干旱的響應
植物各功能性狀對干旱脅迫產生差異化表達,ED和MD處理對優勢種功能性狀的影響較ND更顯著。ED和MD處理顯著降低了藏豆、多裂委陵菜和弱小火絨草的LA,而釘柱委陵菜的LA僅在ED處理下顯著降低(表2)。ED、MD和ND處理下高山嵩草SLA顯著降低,ED和MD處理下藏豆SLA顯著降低,而弱小火絨草的SLA僅在MD處理下顯著降低(P<0.05)。ED處理下高寒草甸、多裂委陵菜和二裂委陵菜的LDMC顯著升高,而弱小委陵菜在ED和MD處理下均顯著增加;MD處理下藏豆、釘柱委陵菜和多裂委陵菜的LCC顯著增加,而二裂委陵菜在ED和MD處理下顯著升高(P<0.05)。藏豆和二裂委陵菜的LNC在ED、MD和ND處理下均顯著降低,高山嵩草、多裂委陵菜在ED和MD處理下顯著降低(P<0.05)。在MD處理下,藏豆、釘柱委陵菜、二裂委陵菜和弱小火絨草的LPC均顯著降低,而高山嵩草在ED和MD處理下均顯著降低(P<0.05)。對于優勢種的C∶N,在ED和ND處理下高山嵩草顯著降低,ED和MD處理下多裂委陵菜顯著升高,而二裂委陵菜在ED、MD和ND 處理下均顯著升高(P<0.05)。在ED、MD和ND處理下,高寒草甸的N∶P顯著升高,而ED和MD處理下多裂委陵菜的N∶P顯著降低,ED和ND處理下二裂委陵菜N∶P顯著降低,而藏豆僅在MD處理下顯著升高(P<0.05)。

表2 不同生長時期干旱處理下植物功能性狀的多重比較結果
2.3 群落功能性狀對不同時期干旱的響應
各群落功能性狀多受干旱脅迫抑制,僅群落葉干物質含量和葉碳氮比表現出了正向響應(圖4)。在ED和MD干旱處理下LACWM、SLACWM、LNCCWM、LPCCWM和N∶PCWM均顯著低于CK(P<0.05),且ED處理對其的影響更強。在ND處理下僅LACWM和LPCCWM顯著低于對照(P<0.05),但仍高于ED和MD處理。同時干旱處理顯著增加了群落的LDMCCWM(P<0.05),且ED處理下LDMCCWM高于MD和ND處理(圖4)。各時期干旱處理下群落LCCCWM均與CK無顯著差異(圖4)。群落C∶NCWM僅在ED處理下顯著升高(圖4)。
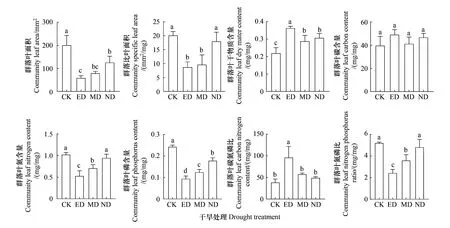
圖4 群落功能性狀對不同時期干旱的響應Fig.4 Responses of community functional traits to drought in different timing不同小寫字母代表組間差異顯著,顯著性水平P<0.05;圖中數據為平均值±標準差
2.4 群落功能多樣性對不同時期干旱的響應
不同時期干旱處理顯著改變了群落的功能多樣性。對照處理下各功能多樣性指數分別為,FEve指數為0.77±0.04,FDiv指數為0.88±0.01,FDis指數為2.44±0.10以及RaoQ指數為6.40±0.36(圖5)。FEve指數在MD處理下顯著低于CK(P<0.05),與CK的差值最低值達到-0.12(圖5)。ED處理下FDis、FDiv指數均有顯著低于CK(P<0.05),與CK的差值最低分別達到-0.33和-0.12(圖5)。ED處理下RaoQ指數顯著高于CK(P<0.05),與CK的差值最高達到1.64(圖5)。ND處理下各群落功能多樣性均與CK無顯著差異。
2.5 群落功能性狀與功能多樣性對干旱響應的相關性
各群落功能性狀的變化對FDiv、FDis以及RaoQ指數改變的相關性較為一致,而與ΔFEve的相關性存在差異(圖6)。ΔFEve與ΔLACWM(r=0.81,P<0.01)和ΔLPCCWM(r=0.75,P<0.05)呈顯著正相關關系。而ΔFDiv、ΔFDis以及ΔRaoQ與ΔLDMCCWM、ΔLCCCWM和ΔC∶NCWM的響應呈正相關關系,其中ΔLDMCCWM與ΔFDiv的影響呈顯著正相關(P<0.05),ΔLCCCWM和ΔC∶NCWM與ΔFDiv、ΔFDis以及ΔRaoQ之間均顯著正相關。而ΔFDiv、ΔFDis和ΔRaoQ與ΔLNCCWM、ΔLPCCWM、ΔN∶PCWM呈負相關關系,其中ΔFDiv與ΔLNCCWM(r=-0.83,P<0.01)、ΔLPCCWM(r=-0.73,P<0.05)以及ΔN∶PCWM(r=-0.69,P<0.05)呈顯著負相關,ΔFDis和ΔRaoQ與ΔN∶PCWM呈顯著負相關(r=-0.73和r=-0.71,P<0.05)。
3 討論
3.1 高寒草甸功能性狀對不同時期干旱的響應
藏北高寒草甸各植物功能性狀對干旱脅迫存在差異化響應。中高寒草甸優勢種的SLA、LA在生長季前期和中期干旱處理下顯著降低,其原因是干旱脅迫破壞了植物水分吸收和蒸騰作用間的動態平衡,導致植物葉片氣孔關閉、葉綠素含量降低,蒸騰速率和光合作用活力降低,CO2同化過程受到抑制,干物質積累受限[30],生產力下降的同時引起SLA、LA的降低。主要是土壤水分不足以維持植物蒸騰需要時,葉片萎蔫甚至凋落,以減少水分散失[36],更多的同化物分配到根系生長中。在個體表型上,植物葉片趨向于變小變厚以適應干旱脅迫[30]。而葉片薄而富含氮的植物具有高轉化率和高生物量同化率,即SLA、LA、LNC和LPC等功能性狀高時,植物具有高生產力和高競爭力[30]。干旱處理下,多裂委陵菜、二裂委陵菜和弱小火絨草的LDMC顯著增加,同時藏豆、多裂委陵菜和釘柱委陵菜的LCC顯著增加。LDMC和LCC表征植物葉片組織的密度,反映植物競爭資源的能力[37],植物葉片LDMC和LCC高時,葉片厚且壽命長,植物具有較低的生物量同化速率,即低生產力和低競爭力[30]。干旱脅迫使得高寒草甸優勢種葉片形態趨向小而厚的同時葉片干物質含量升高,表明高寒草甸植物通過降低生物量同化速率提升對干旱脅迫的抵御能力[37—38]。
葉片中N和P元素作為光合作用酶和生物能量分子的必需元素,其濃度與植物凈光合速率密切相關。高寒草甸群落功能性狀LNC和LPC在生長季前期和中期干旱處理下顯著降低,Wright提出的“葉片經濟波譜”中認為植物LNC、LPC和SLA與光合速率彼此之間呈顯著正相關,低SLA的植物具有低光捕獲率和凈光合速率[39]。干旱脅迫限制了高寒草甸植物的凈光合速率,高寒草甸群落植物通過調節葉片氮磷含量使其處于較低的相對生長速率以維持在干旱逆境中的生長。同時LNC和LPC共同反映植物體在生態系統中受N限制或P限制的養分供應情況,對照處理下群落N∶P為5.8±2.2,N∶P<10表征植物生長更大程度上受到N元素的限制[39],且在生長季前期和中期干旱處理下N∶P顯著降低,指示著高寒草甸群落在干旱脅迫下N限制加劇。植物碳氮比演化機制研究中提出的“適應性生長假說”指出在強烈N限制環境中,維持高C∶N有助于植物提高養分利用效率從而優先保證植物生存[40], 高寒草甸群落C∶N在生長季前期干旱處理下顯著升高,故高寒草甸植物在干旱脅迫下通過提高養分利用效率以維持生存。
3.2 高寒草甸功能多樣性對不同時期干旱的響應
高寒草甸植物群落功能多樣性對干旱事件的響應可能存在多種調控機制,即植物群落采取不同的干旱策略。以往研究認為,采取耐旱策略的優勢種在群落中占主導地位時,往往使功能多樣性在響應干旱時下降[18—19,34]。這與高寒草甸植物群落對生長季前期干旱的響應相一致,FDis、FDiv顯著降低,即群落中物種間功能離散程度和物種特征值差異性降低,群落物種生態位的分化和資源利用程度降低;而RaoQ指數呈顯著升高,群落物種間的差異度加大。表明高寒草甸植物群落在響應生長季前期干旱時可能更傾向于耐旱策略。此外,干旱脅迫下生態位的分化和物種共存機制卻能促使功能多樣性增加[41],群落應對干旱的策略傾向于避旱型[34]。生長季中期干旱處理下群落FEve多樣性顯著降低,即群落功能特征在空間中豐度分布的均勻性降低[34],高寒草甸植物群落在響應生長季中期干旱時表現出避旱型策略。
3.3 群落功能性狀與功能多樣性對干旱的響應
高寒草甸植物群落應對干旱的策略與不同功能性狀對干旱的響應密不可分。前期研究發現地中海生態系統中群落功能性狀通過影響功能多樣性進而作用于生態系統的多功能性[42],旱地群落內的功能多樣性改善了生態系統的多功能性。此外,植物功能多樣性驅動了黃土高原植物群落的碳存儲功能[43]。FDiv、FDis和RaoQ指數與群落植物的LDMC、LCC、LNC和LPC相關。可見群落功能性狀如群落植物葉片的干物質積累情況、葉碳氮磷元素計量,通過影響功能多樣性作用于生態過程。研究表明群落中具有低SLA和(或)LNC的植物多采取保守型生存策略即耐旱型,以增加在資源有限如干旱脅迫[11,44]的環境中持續存在的能力;而高SLA和LNC的植物則多采取避旱生存策略[30]。 高寒草甸群落應對生長季前期干旱的耐旱策略與植物群落的物質積累和養分循環有密切聯系,植物通過提高養分利用效率忍耐干旱[41]。而功能均勻度指數主要與群落植物的葉面積和葉氮含量相關,高寒草甸群落對生長季中期干旱的避旱策略與植物形態性狀和P元素利用有關。
4 結論
在干旱脅迫下高寒草甸群落植物葉片功能性狀表現為葉片小而厚且壽命長,同化速率降低,并且受氮元素限制加劇。結果中,ED干旱脅迫對群落功能性狀的抑制最強;ED和MD處理顯著改變了群落的功能多樣性,ED處理下群落物種間功能離散程度和物種特征值差異性降低,MD處理下群落功能特征在空間中豐度分布的均勻性降低。功能性狀與群落功能多樣性對ED和MD干旱存在響應差異,表明不同時期干旱可能采取不同的生存策略,即群落對生長季前期干旱采用耐旱策略、對生長季中期干旱采用避旱策略。探討了高寒草甸植物群落功能多樣性對不同生長時期干旱脅迫的響應情況,為預測未來季節性干旱事件對青藏高原高寒草甸植物群落功能性狀、群落特征和功能多樣性的影響提供科學依據。
猜你喜歡
英語世界(2023年10期)2023-11-17 09:18:18
小讀者(2021年2期)2021-03-29 05:03:48
華人時刊(2019年13期)2019-11-17 14:59:54
科學大眾(中學)(2019年3期)2019-05-17 10:04:30
文苑(2018年22期)2018-11-19 02:54:14
汽車觀察(2018年10期)2018-11-06 07:05:26
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
紅領巾·萌芽(2016年1期)2016-09-10 07:22:44
少兒科學周刊·兒童版(2015年10期)2015-11-07 03:42:03
